


Lactiplantibacillus plantarum and Saussurea costus as Therapeutic Agents against a Diabetic Rat Model—Approaches to Investigate Pharmacophore Modeling of Human IkB Kinase and Molecular Interaction with Dehydrocostus Lactone of Saussurea costus
 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. S. costus Collection and Extraction
2.3. LC–MASS Spectrometry Analysis of S. costus Extract
2.4. Sample Collecting, Bacterial Isolation, and Identification
2.5. Bacterial Genomic DNA Purification
2.6. PCR Recipe and Program
2.7. Molecular Identification
2.8. Cells Lysis
2.9. Animals
2.10. Experimental Design
2.11. Body Weight and Body Weight Gain
2.12. Sample Collection and Preparation
2.13. Biochemical Parameters
2.14. Histological Study
2.15. Total RNA Extraction
2.16. cDNA Synthesis
2.17. Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | References |
|---|---|---|
| 16S rRNA | F: 5′-TTTGAGTGAGTGGCGAACTG-3′ R: 5′-TTCATGTAGGCGAGTTGCAG-3′ | This study |
| GAPDH | F: 5′-AAGCTGGTCATCAATGGGAAAC-3′ R: 5′-ACCCCATTTGATGTTAGCGG-3′ | [36] |
| β-actin | F: 5′-CGGAACCGCTCATTGCC-3′ R: 5′-ACCCACACTGTGCCCATCTA-3′ | [36] |
| IKBKB | F: 5′-AGGGTGGTTTTTTATTTTTATTTT-3′ R: 5′-AACCCCCACTAAAACTAACTTAA-3′ | [37] |
| IKBKG | F: 5′-GGCCAAACAGGAGGTGAT-3´ R: 5′-TTCTTCTCGGCCAGCTTC-3´ | [38] |
| NfkB1 | F: 5´-CCTGTCTGAAGCCCTGCTACA-3´ R: 5´-CACACCCTGGTTCAGAAGCTG-3′ | [39] |
| IL-17A | F: 5′-CTTCACCCTGGACTCTGAGC-3′ R: 5′-TGGCGGACAATAGAGGAAAC-3′ | [40] |
| IL-6 | F: 5′-ACCACCCACAACAGACCAGT-3′ R: 5′-CAGAATTGCCATTGCACAAC-3′ | [41] |
| IL-17F | F: 5′-TCTTCAATCAAAACCAGGGCAT-3′ R: 5′-GGAGTTCATGGAGCCGTCTT-3′ | [42] |
| IL-1β | F: 5′-AAATGCCTCGTGCTGTCTGA-3′ R: 5′-AGGCCACAGGGATTTTGTCG-3′ | [43] |
| TNF-α | F: 5′-CCCAGACCCTCACACTCAGATCAT-3′ R: 5′-GCAGCCTTGTCCCTTGAAGAGAA-3′ | [44] |
| TRAF6 | F: 5′-CTCAGCGCTGTGCAAACTAC-3′ R: 5′-GATCAAGGATCGTGAGGCGT-3′ | [45] |
| MAPK | F: 5′-TGATATTTGGTCTGTGGGCTG-3′ R: 5′-TGTTCCACGGCACCTTATTTT-3′ | [42] |
2.18. Molecular Docking
2.18.1. Screening of Ligand Properties and ADMET Prediction
2.18.2. Preparation of the Receptor and Drug
2.18.3. Multiple Molecular Docking
2.18.4. Molecular Dynamic Simulation
2.19. Statistical Analysis
3. Results
3.1. Biochemical and Molecular Identification
3.2. LC Mass Analysis
3.3. Growth Performance Parameters
3.4. Biochemical Parameters
3.5. Histological Examinations
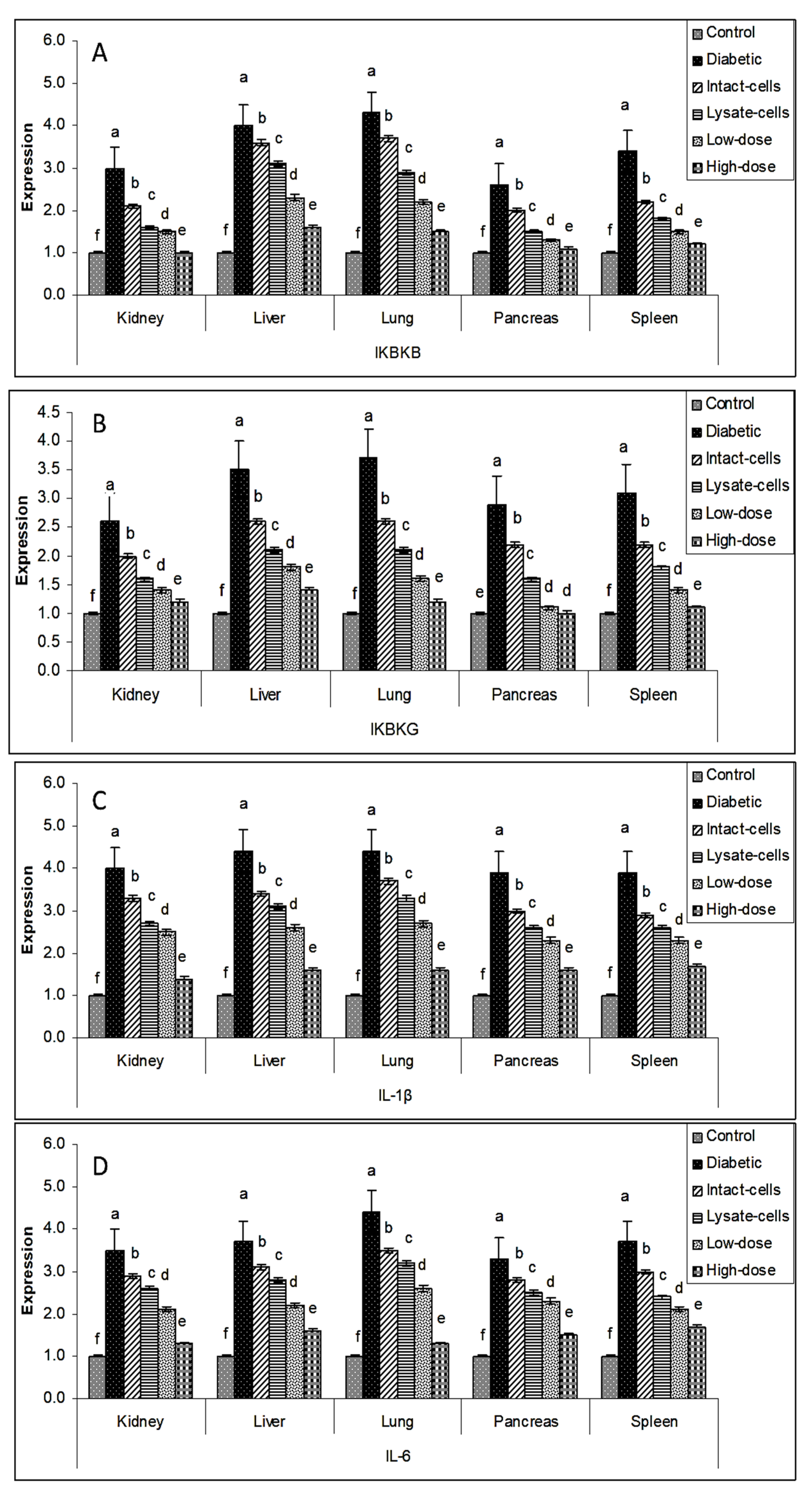
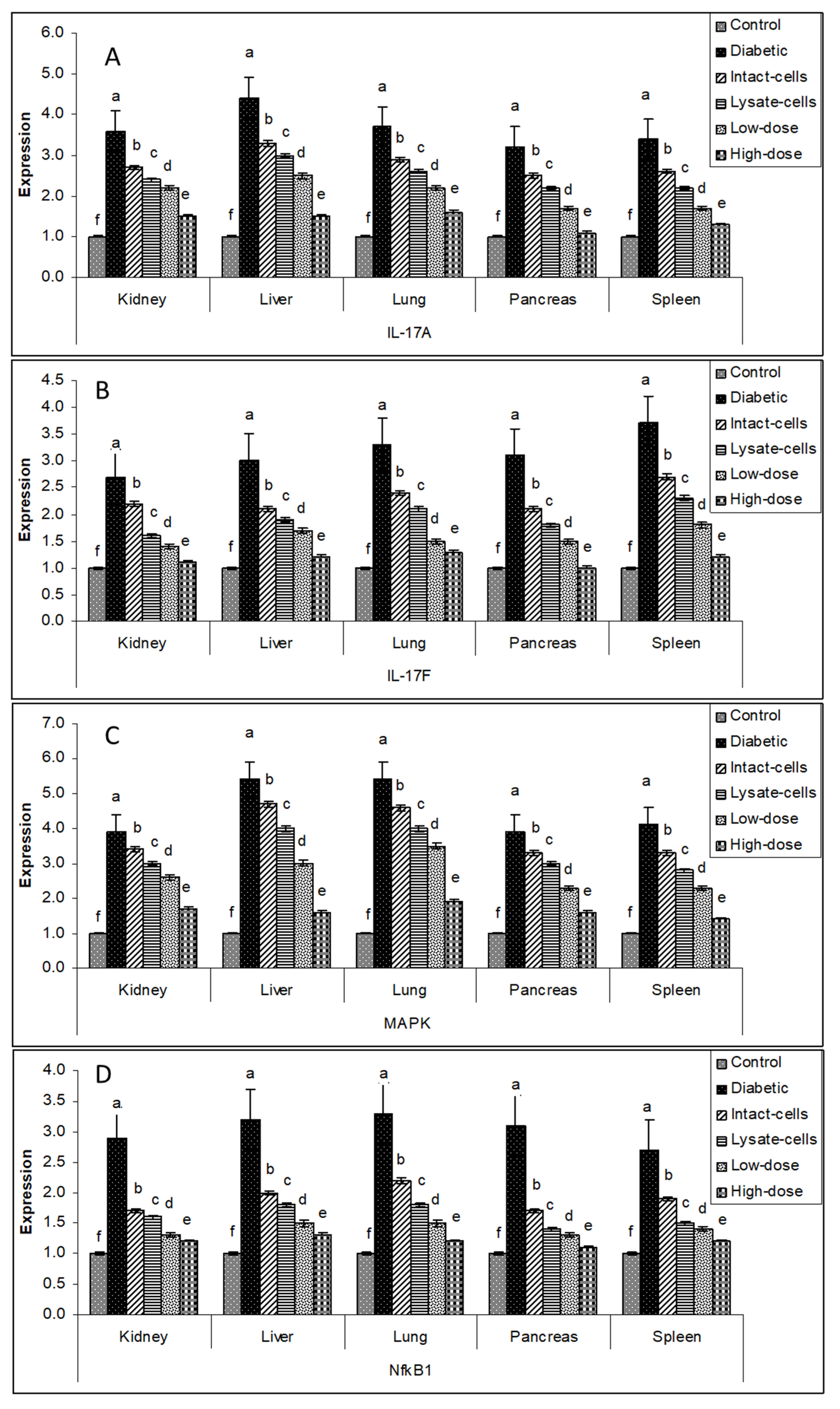
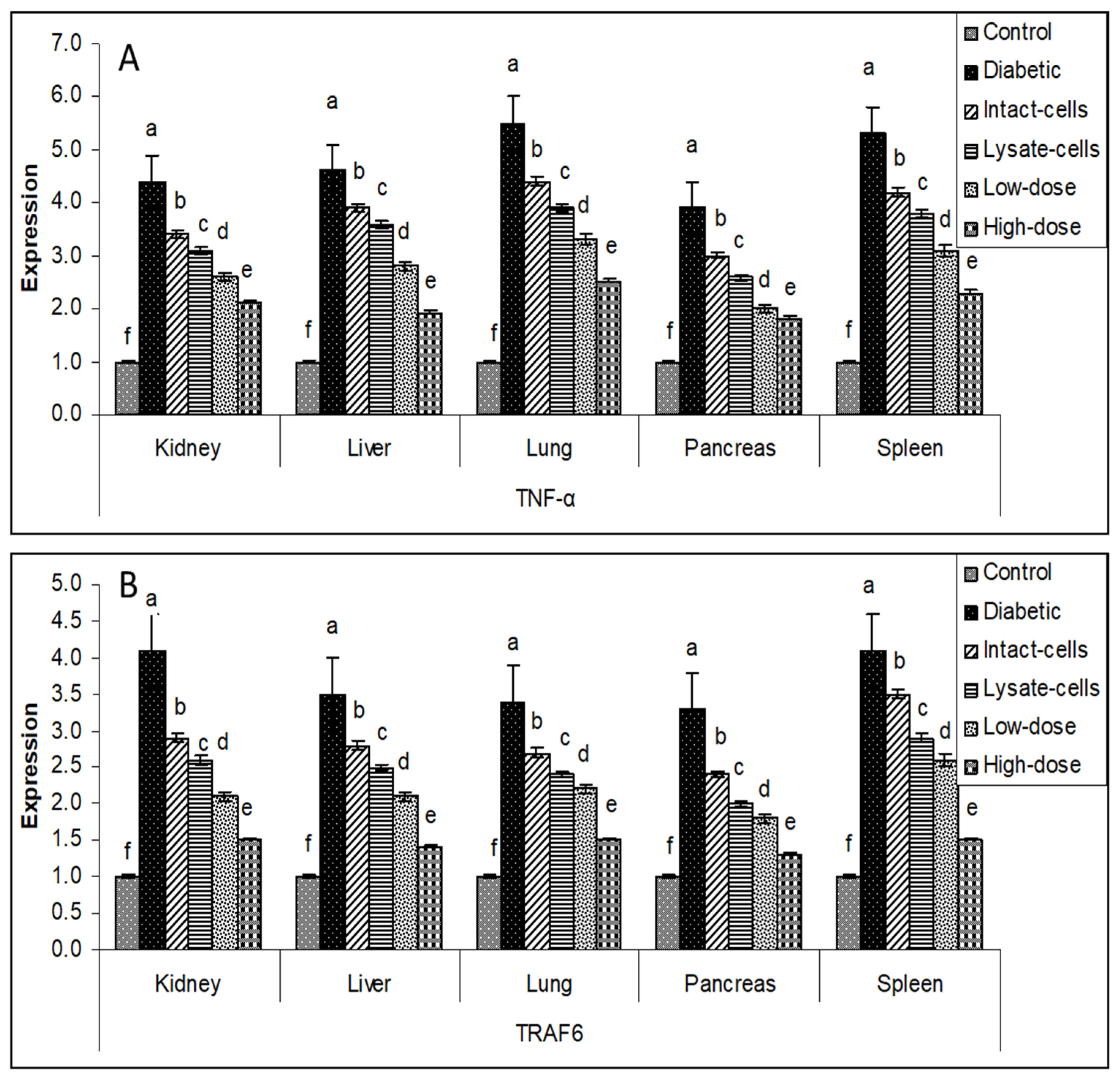
3.6. Gene Expression
3.7. Druglikeness and Toxicity Risk Assessment
3.8. Molecular Docking
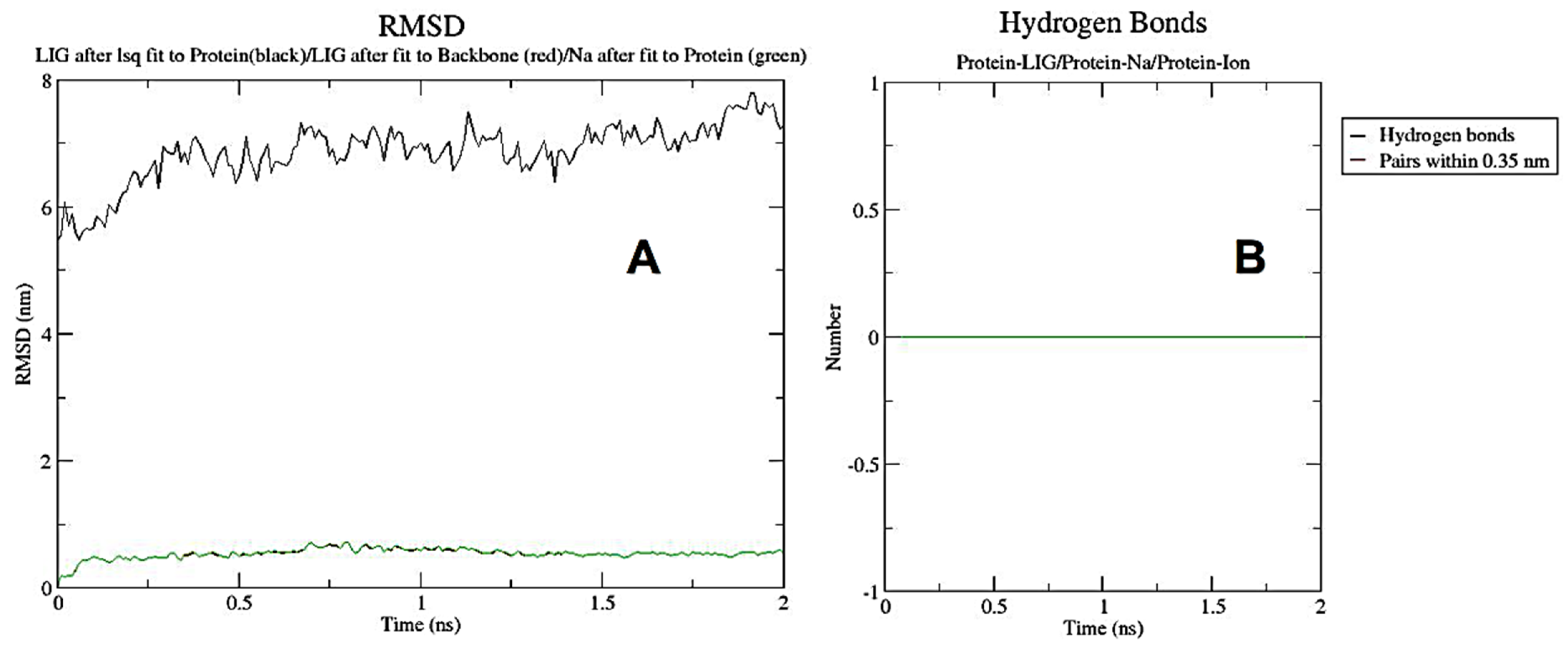
3.9. Molecular Dynamic Simulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lim, L.L.; Chow, E.; Chan, J.C.N. Cardiorenal Diseases in Type 2 Diabetes Mellitus: Clinical Trials and Real-World Practice. Nat. Rev. Endocrinol. 2023, 19, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; D’Alessio, D.A.; Fradkin, J.; Kernan, W.N.; Mathieu, C.; Mingrone, G.; Buse, J.B. Management of Hyperglycaemia in Type 2 Diabetes. A Consensus Report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2022, 65, 1925–1966. [Google Scholar] [CrossRef] [PubMed]
- Hegde, P.K.; Rao, H.A.; Rao, P.N. A Review on Insulin Plant (Costus igneus Nak). Pharmacogn. Rev. 2014, 8, 67–72. [Google Scholar]
- Mammate, N.; El Oumari, F.E.; Imtara, H.; Belchkar, S.; Lahrichi, A.; Alqahtani, A.S.; Houssaini, T.S. Antioxidant and Anti-Urolithiatic Activity of Aqueous and Ethanolic Extracts from Saussurea costus (Falc) Lispich Using Scanning Electron Microscopy. Life 2022, 12, 1026. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, B. Biomedical implications of plant-based principles as antidepressants: Prospects for novel drug development. Mini Rev. Med. Chem. 2022, 22, 904–926. [Google Scholar]
- Naik, A.; Adeyemi, S.B.; Vyas, B.; Krishnamurthy, R. Effect of co-administration of metformin and extracts of Costus pictus D. Don leaves on alloxan-induced diabetes in rats. J. Tradit. Complement. Med. 2022, 12, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.F.; Shetaia, Y.M.; Tayel, A.A.; Munshi, A.M.; Alatawi, F.A.; Alsieni, M.A.; Al-Saman, M.A. Exploring the Antifungal Activity and Action of Saussurea costus Root Extracts against Candida albicans and Non-albicans Species. Antibiotics 2022, 11, 327. [Google Scholar] [CrossRef]
- Reddy, P.J.; sri Motamarry, S.; Varma, K.S.; Anitha, P.; Potti, R.B. Chromatographic analysis of phytochemicals in Costus igneus and computational studies of flavonoids. Inform. Med. Unlocked. 2018, 13, 34–40. [Google Scholar]
- Selvakumarasamy, S. Costus pictus–transition from a medicinal plant to functional food: A review. Future Foods 2021, 4, 100068. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Zhang, C. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- Rani, P.; Santosh, K.T. Health benefits of bacteriocins produced by probiotic lactic acid bacteria. In Microbial Biomolecules; Academic Press: Cambridge, MA, USA, 2023; pp. 97–111. [Google Scholar]
- Tian, L.; Liu, R.; Zhou, Z.; Xu, X.; Feng, S.; Kushmaro, A.; Sun, Q. Probiotic Characteristics of Lactiplantibacillus plantarum N-1 and Its Cholesterol-Lowering Effect in Hypercholesterolemic Rats. Probiotics Antimicrob. Proteins 2022, 14, 337–348. [Google Scholar] [CrossRef]
- Hinz, M.; Scheidereit, C. The IκB kinase complex in NF-κB regulation and beyond. EMBO Rep. 2014, 15, 46–61. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef]
- Albensi, B.C. What Is Nuclear Factor Kappa B (NF-κB) Doing in and to the Mitochondrion? Front. Cell Dev. Biol. 2019, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Elshaer, S.E.; Hamad, G.M.; Hafez, E.E.; Baghdadi, H.H.; El-Demerdash, F.M.; Simal-Gandara, J. Root extracts of Saussurea costus as prospective detoxifying food additive against sodium nitrite toxicity in male rats. Food Chem. Toxicol. 2022, 166, 113225. [Google Scholar] [CrossRef]
- Gheraibia, S.; Belattar, N.; Abdel-Wahhab, M.A. HPLC analysis, antioxidant and cytotoxic activity of different extracts of Costus speciosus against HePG-2 cell lines. S. Afr. J. Bot. 2020, 131, 222–228. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, C.; Yang, L.; Jiang, L.; Zhang, J.; Yu, X.; Jia, H. Spatial and Temporal Persistence of Fluorescent Lactiplantibacillus plantarum RS-09 in Intestinal Tract. Front. Microbiol. 2022, 13, 843650. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.Y.; Atallah, A.A.; Badr, O.A.; Moustafa, M.M.; Esmael, A.; Ebrahim, N.; Mohammed, A.; Basim Al, S.; Ibrahim, A.A.; Mohamed, S.A. Bioprospecting for Novel Probiotic Strains from Human Milk and Infants: Molecular, Biochemical, and Ultrastructural Evidence. Biology 2022, 11, 1405. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef]
- San Millán, R.M.; Martínez-Ballesteros, I.; Rementeria, A.; Garaizar, J.; Bikandi, J. Online exercise for the design and simulation of PCR and PCR-RFLP experiments. BMC Res. Notes 2013, 6, 513. [Google Scholar] [CrossRef] [PubMed]
- Shehadul Islam, M.; Aryasomayajula, A.; Selvaganapathy, P. A Review on Macroscale and Microscale Cell Lysis Methods. Micromachines 2017, 8, 83. [Google Scholar] [CrossRef]
- Nakamura, H.; Ishigami, T.; Kawase, Y.; Yamada, A.; Minagawa, M.; Fukuta, H.; Kurono, Y.; Suzuki, H. Effects of acupuncture stimulation on blood glucose concentration in the Otsuka Long-Evans Tokushima Fatty (OLETF) rat, an animal model for type-2 diabetes mellitus. Med. Sci. Monit. Basic Res. 2014, 20, 70–75. [Google Scholar]
- Imtiaz, F.; Islam, M.; Saeed, H.; Ahmed, A.; Rathore, H.A. Assessment of the antidiabetic potential of extract and novel phytoniosomes formulation of Tradescantia pallida leaves in the alloxan-induced diabetic mouse model. FASEB J. 2023, 37, e22818. [Google Scholar] [CrossRef]
- Rolland, V.; Roseau, S.; Fromentin, G.; Nicolaidis, S.; Tomé, D.; Even, P.C. Body weight, body composition, and energy metabolism in lean and obese Zucker rats fed soybean oil or butter. Am. J. Clin. Nutr. 2002, 75, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Cubero, J.; Franco, L.; Mesa, M.; Galán, C.; Rodríguez, A.B.; Jarne, C.; Barriga, C. Body weight gain in rats by a high-fat diet produces chronodisruption in activity/inactivity circadian rhythm. Chronobiol. Int. 2014, 31, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Tang, J.; Chen, X. Sensitive determination of malondialdehyde in rat prostate by high performance liquid chromatography with fluorescence detection. Sci. Rep. 2020, 10, 3990. [Google Scholar] [CrossRef]
- Nuhu, F.; Gordon, A.; Sturmey, R.; Seymour, A.M.; Bhandari, S. Measurement of Glutathione as a Tool for Oxidative Stress Studies by High Performance Liquid Chromatography. Molecules 2020, 25, 4196. [Google Scholar] [CrossRef]
- Aberare, O.L.; Gordon, A.; Sturmey, R.; Seymour, A.M.; Bhandari, S. Triglycerides, total cholesterol, high density lipoprotein cholesterol and low density lipoprotein cholesterol in rats exposed to premium motor spirit fumes. N. Am. J. Med. Sci. 2011, 3, 277–280. [Google Scholar] [CrossRef]
- Ojiako, O.A.; Chikezie, P.C.; Ogbuji, A.C. Blood glucose level and lipid profile of alloxan-induced hyperglycemic rats treated with single and combinatorial herbal formulations. J. Tradit. Complement. Med. 2016, 6, 184–192. [Google Scholar] [CrossRef]
- Chao, P.C.; Li, Y.; Chang, C.H.; Shieh, J.P.; Cheng, J.T.; Cheng, K.C. Investigation of insulin resistance in the popularly used four rat models of type-2 diabetes. Biomed. Pharmacother. 2018, 101, 155–161. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Acknowledgments. In Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Suvarna, S.K., Layton, C., Bancroft, J.D., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. ix–x. [Google Scholar] [CrossRef]
- Brunt, E.M. Grading and staging the histopathological lesions of chronic hepatitis: The Knodell histology activity index and beyond. Hepatology 2000, 31, 241–246. [Google Scholar] [CrossRef]
- Badr, O.A.; EL-Shawaf, I.I.; El-Garhy, H.A.; Moustafa, M.M.; Ahmed-Farid, O.A. The potent therapeutic effect of novel cyanobacterial isolates against oxidative stress damage in redox rats. J. Appl. Microbiol. 2019, 126, 1278–1289. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods San Diego Calif. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Muller, D.N.; Mullally, A.; Dechend, R.; Park, J.K.; Fiebeler, A.; Pilz, B.; Löffler, B.M.; Blum-Kaelin, D.; Masur, S.; Dehmlow, H.; et al. Endothelin-converting enzyme inhibition ameliorates angiotensin II-induced cardiac damage. Hypertension 2002, 40, 840–846. [Google Scholar] [CrossRef]
- Bialek, K.; Czarny, P.; Wigner, P.; Synowiec, E.; Kolodziej, L.; Bijak, M.; Szemraj, J.; Papp, M.; Sliwinski, T. Agomelatine Changed the Expression and Methylation Status of Inflammatory Genes in Blood and Brain Structures of Male Wistar Rats after Chronic Mild Stress Procedure. Int. J. Mol. Sci. 2022, 23, 8983. [Google Scholar] [CrossRef]
- Wu, C.X.; Mao, C.Y.; Deng, J.; Zhang, T.; Zhao, Y.; Guan, Z.Z.; Hu, X.X.; Qi, X.L. Fluoride induced down-regulation of IKBKG Gene expression inhibits hepatocytes senescence. J. Trace Elem. Med. Biol. 2022, 69, 126896. [Google Scholar] [CrossRef]
- Ruslanova, B.; Abishev, Z.; Chaizhunussova, N.; Shabdarbayeva, D.; Tokesheva, S.; Amantayeva, G.; Kairkhanova, Y.; Stepanenko, V.; Hoshi, M.; Fujimoto, N. Hepatic gene expression changes in rats internally exposed to radioactive 56MnO2 particles at low doses. Curr. Issues Mol. Biol. 2021, 43, 758–766. [Google Scholar] [CrossRef]
- Ávalos, A.M.; Apablaza, F.A.; Quiroz, M.; Toledo, V.; Peña, J.P.; Michea, L.; Peña, D.; Michea, L.; Figueroa, F.E. IL-17A levels increase in the infarcted region of the left ventricle in a rat model of myocardial infarction. Biol. Res. 2012, 45, 193–200. [Google Scholar] [CrossRef]
- Loof, T.; Krämer, S.; Gaedeke, J.; Neumayer, H.H.; Peters, H. IL-17 Expression in the Time Course of Acute Anti-Thy1 Glomerulonephritis. PLoS ONE 2016, 11, e0156480. [Google Scholar] [CrossRef]
- Aslani, M.R.; Sharghi, A.; Boskabady, M.H.; Ghobadi, H.; Keyhanmanesh, R.; Alipour, M.R.; Ahmadi, M.; Saadat, S.; Naghizadeh, P. Altered gene expression levels of IL-17/TRAF6/MAPK/USP25 axis and pro-inflammatory cytokine levels in lung tissue of obese ovalbumin-sensitized rats. Life Sci. 2022, 296, 120425. [Google Scholar] [CrossRef] [PubMed]
- Pandur, E.; Szabó, I.; Hormay, E.; Pap, R.; Almási, A.; Sipos, K.; Farkas, V.; Karádi, Z. Alterations of the expression levels of glucose, inflammation, and iron metabolism related miRNAs and their target genes in the hypothalamus of STZ-induced rat diabetes model. Diabetol. Metab. Syndr. 2022, 14, 147. [Google Scholar] [CrossRef]
- Ghatpande, N.S.; Misar, A.V.; Waghole, R.J.; Jadhav, S.H.; Kulkarni, P.P. Tinospora cordifolia protects against inflammation associated anemia by modulating inflammatory cytokines and hepcidin expression in male Wistar rats. Sci. Rep. 2019, 9, 10969. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, H.; Shi, J.; Huang, T.; Wang, Y. miRNA-146a and miRNA-202-3p Attenuate Inflammatory Response by Inhibiting TLR4, IRAK1, and TRAF6 Expressions in Rats following Spinal Cord Injury. J. Nanomater. 2021, 2021, e5452239. [Google Scholar] [CrossRef]
- Lemkul, J.A. From Proteins to Perturbed Hamiltonians: A Suite of Tutorials for the GROMACS-2018 Molecular Simulation Package [Article v1.0]. Living J. Comput. Mol. Sci. 2019, 1, 5068. [Google Scholar] [CrossRef]
- Li, J.; Liu, G.; Zhen, Z.; Shen, Z.; Li, S.; Li, H. Molecular Docking for Ligand-Receptor Binding Process Based on Heterogeneous Computing. Sci. Program. 2022, 2022, e9197606. [Google Scholar] [CrossRef]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP: Expanding the scope of the protein–ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef]
- SantaCruz-Calvo, S.; Bharath, L.; Pugh, G.; SantaCruz-Calvo, L.; Lenin, R.R.; Lutshumba, J.; Liu, R.; Bachstetter, A.D.; Beibei ZhuNikolajczyk, B.S. Adaptive immune cells shape obesity-associated type 2 diabetes mellitus and less prominent comorbidities. Nat. Rev. Endocrinol. 2022, 18, 23–42. [Google Scholar] [CrossRef]
- Vanweert, F.; Schrauwen, P.; Phielix, E. Role of branched-chain amino acid metabolism in the pathogenesis of obesity and type 2 diabetes-related metabolic disturbances BCAA metabolism in type 2 diabetes. Nutr. Diabetes 2022, 12, 1–13. [Google Scholar]
- Reid, R.L.; Hinks, N.T.; Mills, S.C. Alloxan Diabetes In Pregnant Ewes. J. Endocrinol. 1963, 27, 1–19. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef]
- Yang, S.E.; Lin, Y.F.; Liao, J.W.; Chen, J.T.; Chen, C.L.; Chen, C.I.; Hsu, S.L.; Song, T.Y. Insulin sensitizer and antihyperlipidemic effects of Cajanus cajan (L.) millsp. root in methylglyoxal-induced diabetic rats. Chin. J. Physiol. 2022, 65, 125. [Google Scholar] [PubMed]
- Kim, E.J.; Kim, B.H.; Seo, H.S.; Lee, Y.J.; Kim, H.H.; Son, H.H.; Choi, M.H. Cholesterol-induced non-alcoholic fatty liver disease and atherosclerosis aggravated by systemic inflammation. PLoS ONE 2014, 9, e97841. [Google Scholar] [CrossRef] [PubMed]
- Marchesini, G.; Brizi, M.; Morselli-Labate, A.M.; Bianchi, G.; Bugianesi, E.; McCullough, A.J.; Forlani, G.; Melchionda, N. Association of nonalcoholic fatty liver disease with insulin resistance. Am. J. Med. 1999, 107, 450–455. [Google Scholar] [CrossRef]
- Eliza, J.; Daisy, P.; Ignacimuthu, S.; Duraipandiyan, V. Antidiabetic and antilipidemic effect of eremanthin from Costus speciosus (Koen.)Sm., in STZ-induced diabetic rats. Chem. Biol. Interact. 2009, 182, 67–72. [Google Scholar] [CrossRef]
- Reis, S.A.; Conceição, L.L.; Rosa, D.D.; Siqueira, N.P.; Peluzio, M.C.G. Mechanisms responsible for the hypocholesterolaemic effect of regular consumption of probiotics. Nutr. Res. Rev. 2017, 30, 36–49. [Google Scholar] [CrossRef]
- Tian, Y.; Wang, Y.; Zhang, N.; Xiao, M.; Zhang, J.; Xing, X.; Zhang, Y.; Fan, Y.; Li, X.; Nan, B.; et al. Antioxidant Mechanism of Lactiplantibacillus plantarum KM1 Under H2O2 Stress by Proteomics Analysis. Front. Microbiol. 2022, 13, 2237. [Google Scholar] [CrossRef]
- Wang, L.-X.; Liu, K.; Gao, D.-W.; Hao, J.-K. Protective effects of two Lactobacillus plantarum strains in hyperlipidemic mice. World J. Gastroenterol. 2013, 19, 3150–3156. [Google Scholar] [CrossRef] [PubMed]
- Werning, M.L.; Hernández-Alcántara, A.M.; Ruiz, M.J.; Soto, L.P.; Dueñas, M.T.; López, P.; Frizzo, L.S. Biological Functions of Exopolysaccharides from Lactic Acid Bacteria and Their Potential Benefits for Humans and Farmed Animals. Foods 2022, 11, 1284. [Google Scholar] [CrossRef]
- Pisosch, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Geicu, O.I.; Bilteanu, L.; Serban, A.I. Antioxidant, anti-inflammatory and immunomodulatory roles of vitamins in COVID-19 therapy. Eur. J. Med. Chem. 2022, 232, 114175. [Google Scholar] [CrossRef] [PubMed]
- El-Kholie, E.; El Sheikh, N.; Hassan, M. Potential Anti-obesity Effects of Costus (Saussurea costus) and Bitter Melon (Citrullus colocynthis (in Rats Fed on High Fat Diet. J. Home Econ.-Menofia Univ. 1999, 32, 127–140. [Google Scholar]
- Shediwah, F.M.H.; Naji, K.M.; Gumaih, H.S.; Alhadi, F.A.; Al-Hammami, A.L.; D’Souza, M.R. Antioxidant and antihyperlipidemic activity of Costus speciosus against atherogenic diet-induced hyperlipidemia in rabbits. J. Integr. Med. 2019, 17, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, M.; Hassan, T.M.; Baba, S.S.; Radda, M.I.; Mutawakkil, M.M.; Musa, M.A.; AbuBakar, S.; Loong, S.K.; Loong Yusuf, I. Exploring NFkB pathway as a potent strategy to mitigate COVID-19 severe morbidity and mortality. J. Public Health Afr. 2022, 13, 1679. [Google Scholar] [CrossRef]
- Shin, C.; Choi, D.S. Essential Roles for the Non-Canonical IκB Kinases in Linking Inflammation to Cancer, Obesity, and Diabetes. Cells 2019, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sahu, K.; Singh, C.; Singh, A. Lipopolysaccharide induced altered signaling pathways in various neurological disorders. Naunyn. Schmiedebergs Arch. Pharmacol. 2022, 395, 285–294. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF Family Cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Püschel, G.P.; Klauder, J.; Henkel, J. Macrophages, Low-Grade Inflammation, Insulin Resistance and Hyperinsulinemia: A Mutual Ambiguous Relationship in the Development of Metabolic Diseases. J. Clin. Med. 2022, 11, 4358. [Google Scholar] [CrossRef]
- Kwiat, V.R.; Reis, G.; Valera, I.C.; Parvatiyar, K.; Parvatiyar, M.S. Autoimmunity as a sequela to obesity and systemic inflammation. Front. Physiol. 2022, 13, 2393. [Google Scholar] [CrossRef] [PubMed]
- Yip, L.; Fuhlbrigge, R.; Alkhataybeh, R.; Fathman, C.G. Gene Expression Analysis of the Pre-Diabetic Pancreas to Identify Pathogenic Mechanisms and Biomarkers of Type 1 Diabetes. Front. Endocrinol. 2020, 11, 609271. [Google Scholar] [CrossRef]
- Revathy, J.; Abdullah, S.S.; Kumar, P.S. Antidiabetic effect of Costus speciosus rhizome extract in alloxan induced albino rats. J. Chem. Biochem. 2014, 2, 13–22. [Google Scholar]















| Groups | Parameters | ||
|---|---|---|---|
| Initial Body Weight (g) | Final Body Weight (g) | Body Weight Gain (g) | |
| Control | 154 ± 5.85 a | 291 ± 2.36 a | 137 ± 5.74 a |
| Diabetic | 159 ± 11.23 a | 153 ± 2.12 e | −6 ± 12.15 e |
| Diabetic + intact cells | 161 ± 5.29 a | 203 ± 1.51 d | 42 ± 4.25 d |
| Diabetic + lysate cells | 166 ± 9.81 a | 235 ± 3.11 c | 69 ± 7.88 c |
| Diabetic + low dose | 168 ± 7.78 a | 237 ± 2.16 c | 69 ± 8.91 c |
| Diabetic + high dose | 165 ± 2.88 a | 255 ± 3.00 b | 91 ± 2.87 b |
| Groups | Parameters | ||
|---|---|---|---|
| MDA nmol/mL | GSH µmol/mL | GSSG µmol/mL | |
| Control | 83.9 ± 3.49 e | 3.6 ± 0.14 a | 0.74 ± 0.03 d |
| Diabetic | 195.8 ± 8.42 a | 2.0 ± 0.08 b | 1.82 ± 0.06 a |
| Diabetic + intact cells | 157.1 ± 6.65 b | 2.5 ± 0.11 b | 1.74 ± 0.08 a |
| Diabetic + lysate cells | 149.4 ± 6.05 b | 2.5 ± 0.11 b | 1.47 ± 0.06 b |
| Diabetic + low dose | 130.3 ± 5.55 c | 3.0 ± 0.12 a | 1.41 ± 0.05 b |
| Diabetic + high dose | 106.9 ± 4.26 d | 3.6 ± 0.15 a | 1.11 ± 0.04 c |
| Groups | Parameters | |||
|---|---|---|---|---|
| TG mg dL−1 | TC mg dL−1 | HDL mg dL−1 | LDL mg dL−1 | |
| Control | 96.6 ± 4.03 d | 83.8 ± 3.033 e | 34.5 ± 1.29 a | 45.7 ± 21 f |
| Diabetic | 231.1 ± 10.02 a | 215.9 ± 7.88 a | 19.5 ± 0.79 d | 162.2 ± 6.98 a |
| Diabetic + intact cells | 230.6 ± 8.99 a | 209.4 ± 8.54 a | 24.3 ± 0.94 c | 150.7 ± 6.02 b |
| Diabetic + lysate cells | 184.7 ± 8.17 b | 186.4 ± 7.724 b | 23.2 ± 0.88 c | 134.2 ± 4.87 c |
| Diabetic + low dose | 165.2 ± 6.54 c | 165.1 ± 6.07 c | 31.4 ± 1.28 b | 120 ± 4.93 d |
| Diabetic + high dose | 158.4 ± 5.67 c | 132.4 ± 5.59 d | 33.3 ± 1.32 a | 83.2 ± 3.51 e |
| Groups | Parameters | ||
|---|---|---|---|
| Glucose mg dL−1 | Insulin pmol L−1 | HOMA-IR | |
| Control | 92.4 ± 3.44 f | 57.5 ± 2.14 a | 13.1 ± 0.1 c |
| Diabetic | 233.5 ± 8.81 a | 28.8 ± 1.12 d | 16.6 ± 0.12 b |
| Diabetic + intact cells | 221.4 ± 8.81 b | 31.9 ± 1.28 cd | 17.4 ± 0.29 b |
| Diabetic + lysate cells | 193.3 ± 7.31 c | 35.8 ± 1.37 c | 17.1 ± 0.35 b |
| Diabetic + low dose | 57.5 ± 6.83 d | 44.9 ± 1.96 b | 18.4 ± 0.39 a |
| Diabetic + high dose | 152.9 ± 6.54 e | 47.2 ± 1.84 b | 17.8 ± 0.36 a |
| Groups | Histological Score | Degenerative Changes | Type of Pathological Changes |
|---|---|---|---|
| Control | 1 | No abnormalities | None |
| Diabetic | 5 | Extensive and marked | Fatty degeneration (microvesicular and macrovesicular steatosis), fibrosis, inflammation, and blood vessel congestion. |
| Diabetic + intact cells | 4 | Severe | Moderate fatty degeneration (microvesicular and macrovesicular steatosis), inflammation, and blood vessel congestion. |
| Diabetic + lysate cells | 3 | Moderate | Minimal fatty degeneration (microvesicular and macrovesicular steatosis), inflammation, and blood vessel congestion. |
| Diabetic + low dose | 3 | Moderate | Minimal fatty degeneration (microvesicular and macrovesicular steatosis), inflammation, and congested hepatic sinusoids. |
| Diabetic + high dose | 2 | Mild | Minimal microvesicular steatosis, few congested hepatic sinusoids. |
| Compound | Formula | Similarity Score | MW (<500 Da) | H-Bond Acceptors (<10) | H-Bond Donors (<5) | Consensus Log P (<5) | Lipinski Violations | Ali Log S |
|---|---|---|---|---|---|---|---|---|
| Dehydrocostus lactone | C15H18O2 | 1 | 230.3 | 2 | 0 | 2.98 | 0 | −2.82 |
| Bigelovin | C17H20O5 | 0.943 | 304.34 | 5 | 0 | 1.89 | 0 | −2.25 |
| Caprolactone | C6H10O2 | 0.90 | 114.14 | 2 | 0 | 1.08 | 0 | −0.11 |
| Methyl nonanoate ester | C10H20O2 | 0.816 | 172.26 | 2 | 0 | 3.13 | 0 | −4.59 |
| (3E)-3-[(phenylamino)methylidene]oxan-2-one | C12H13NO2 | 0.748 | 203.24 | 2 | 1 | 2.14 | 0 | −2.74 |
| Gamma-valerolactone | C5H8O2 | 0.739 | 100.12 | 2 | 0 | 0.91 | 0 | −0.74 |
| Ethyl levulinate | C7H12O3 | 0.735 | 144.17 | 3 | 0 | 0.81 | 0 | −0.53 |
| Butyl acetate | C6H12O2 | 0.698 | 116.16 | 2 | 0 | 1.53 | 0 | −1.95 |
| (3E)-3-[(phenylamino)methylidene]dihydrofuran-2(3H)-one | C11H11NO2 | 0.646 | 189.21 | 2 | 1 | 1.84 | 0 | −2.37 |
| R-carvone | C10H14O | 0.583 | 150.22 | 1 | 0 | 2.44 | 0 | −2.72 |
| Compound | Hepatotoxicity (P) | Carcinogenicity (P) | Immunotoxicity (P) | Mutagenicity (P) | Cytotoxicity (P) | LD50 mg kg−1 |
|---|---|---|---|---|---|---|
| Dehydrocostus lactone | 0.59 i | 0.61 i | 0.94 a | 0.72 i | 0.76 i | 1330 |
| Bigelovin | 0.61 i | 0.50 i | 0.99 i | 0.78 i | 0.81 i | 125 |
| Caprolactone | 0.72 i | 0.61 i | 0.98 i | 0.96 i | 0.66 i | 4290 |
| Methyl nonanoate ester | 0.58 i | 0.55 i | 0.99 i | 0.98 i | 0.73 i | 5000 |
| (3E)-3-[(phenylamino)methylidene]oxan-2-one | 0.62 i | 0.50 i | 0.99 i | 0.72 i | 0.64 i | 1800 |
| Gamma-valerolactone | 0.66 i | 0.54 i | 0.98 i | 0.91i | 0.66 i | 8800 |
| Ethyl levulinate | 0.78 i | 0.59 a | 0.99 i | 0.96 i | 0.82 i | 5000 |
| Butyl acetate | 0.77 i | 0.57 a | 0.98 i | 0.97 i | 0.81 i | 6000 |
| (3E)-3-[(phenylamino)methylidene]dihydrofuran-2(3H)-one | 0.58 i | 0.53 i | 0.99 i | 0.69 i | 0.67 i | 2761 |
| R-carvone | 0.65 i | 0.83 i | 0.99 i | 0.97 i | 0.80 i | 1640 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AlGeffari, M.A.; Mansour, D.; Ahmed-Farid, O.; Mohamed Yousef, E.; Mohamed, S.A.; Moustafa, M.M.A.; Barakat, H.; Abd El Ghany, K. Lactiplantibacillus plantarum and Saussurea costus as Therapeutic Agents against a Diabetic Rat Model—Approaches to Investigate Pharmacophore Modeling of Human IkB Kinase and Molecular Interaction with Dehydrocostus Lactone of Saussurea costus. Metabolites 2023, 13, 764. https://doi.org/10.3390/metabo13060764
AlGeffari MA, Mansour D, Ahmed-Farid O, Mohamed Yousef E, Mohamed SA, Moustafa MMA, Barakat H, Abd El Ghany K. Lactiplantibacillus plantarum and Saussurea costus as Therapeutic Agents against a Diabetic Rat Model—Approaches to Investigate Pharmacophore Modeling of Human IkB Kinase and Molecular Interaction with Dehydrocostus Lactone of Saussurea costus. Metabolites. 2023; 13(6):764. https://doi.org/10.3390/metabo13060764
Chicago/Turabian StyleAlGeffari, Metab A., Dina Mansour, Omar Ahmed-Farid, Einas Mohamed Yousef, Shereen A. Mohamed, Mahmoud M. A. Moustafa, Hassan Barakat, and Khalid Abd El Ghany. 2023. "Lactiplantibacillus plantarum and Saussurea costus as Therapeutic Agents against a Diabetic Rat Model—Approaches to Investigate Pharmacophore Modeling of Human IkB Kinase and Molecular Interaction with Dehydrocostus Lactone of Saussurea costus" Metabolites 13, no. 6: 764. https://doi.org/10.3390/metabo13060764
APA StyleAlGeffari, M. A., Mansour, D., Ahmed-Farid, O., Mohamed Yousef, E., Mohamed, S. A., Moustafa, M. M. A., Barakat, H., & Abd El Ghany, K. (2023). Lactiplantibacillus plantarum and Saussurea costus as Therapeutic Agents against a Diabetic Rat Model—Approaches to Investigate Pharmacophore Modeling of Human IkB Kinase and Molecular Interaction with Dehydrocostus Lactone of Saussurea costus. Metabolites, 13(6), 764. https://doi.org/10.3390/metabo13060764