

Oral Administration of Chaetoceros gracilis—A Marine Microalga—Alleviates Hepatic Lipid Accumulation in Rats Fed a High-Sucrose and Cholesterol-Containing Diet

 ,
, 
Abstract
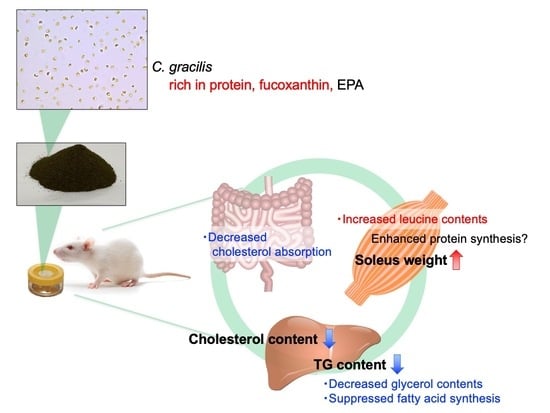
1. Introduction
2. Experimental Design
2.1. Materials
2.2. Animals and Diets
2.3. Measurement of Serum Biochemical Parameters
2.4. Measurement of Triglycerides, Cholesterol, Phospholipids, and Glycogen Contents in the Liver
2.5. Assays for Hepatic Enzyme Activities
2.6. Measurement of Serum Steroid Levels
2.7. Analysis of Hepatic mRNA Levels
2.8. Analysis of Water-Soluble Metabolites in the Liver and Soleus Muscle (Non-Target Metabolome Analysis)
2.9. Statistical Analysis
3. Results
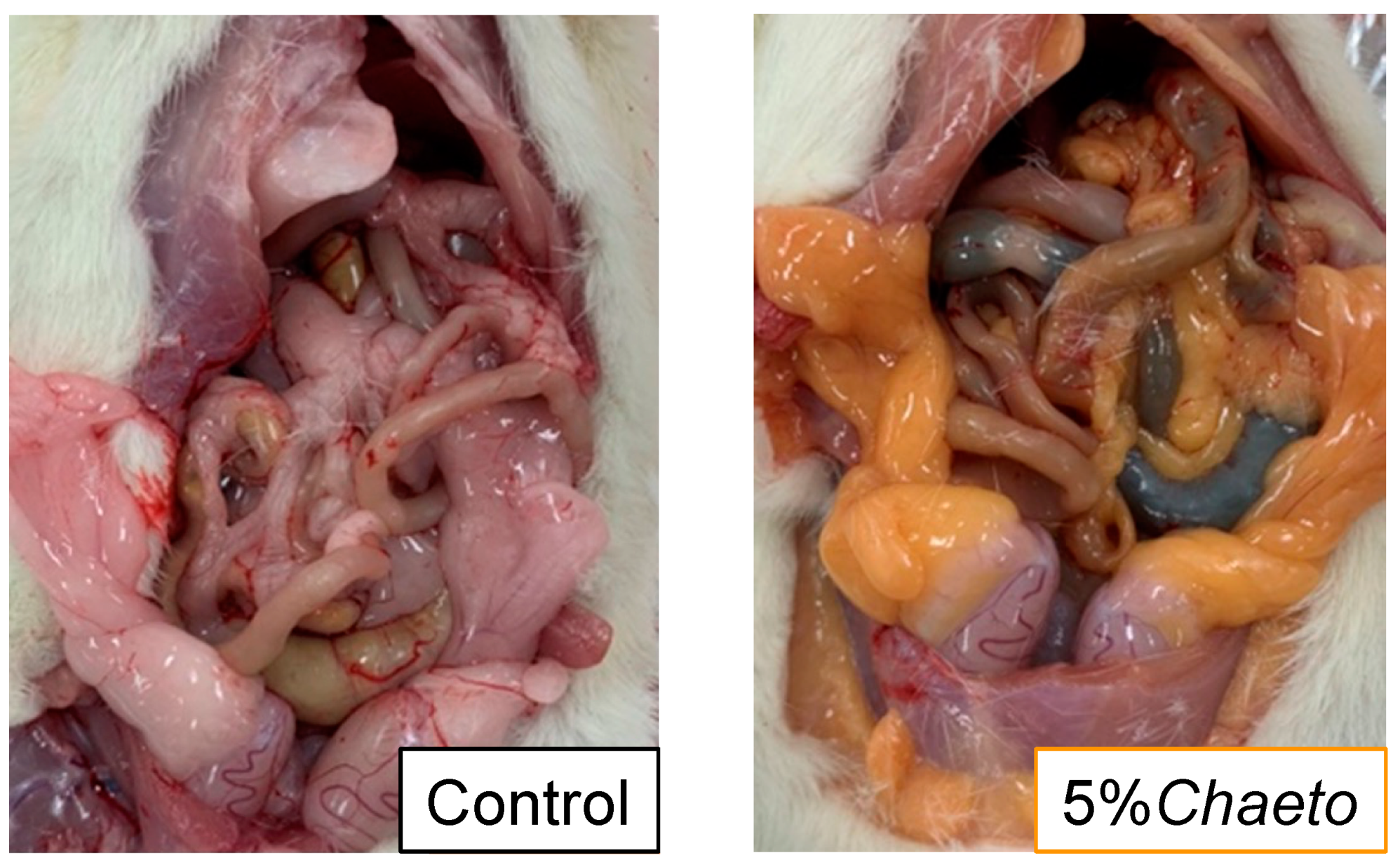
3.1. Effects of C. gracilis Feeding on Morphometric Variables in SD Rats
3.2. Effects of C. gracilis Feeding on Hepatic Lipid and Glycogen Contents and Serum Biochemical Parameters in SD Rats
3.3. Effects of C. gracilis Feeding on Activities of Hepatic Enzymes and mRNA Levels Related to Fatty Acid Metabolism in SD Rats
3.4. Effects of C. gracilis Feeding on Serum Levels of Steroids and Relative Levels of Hepatic mRNA Related to Cholesterol Metabolism in SD Rats
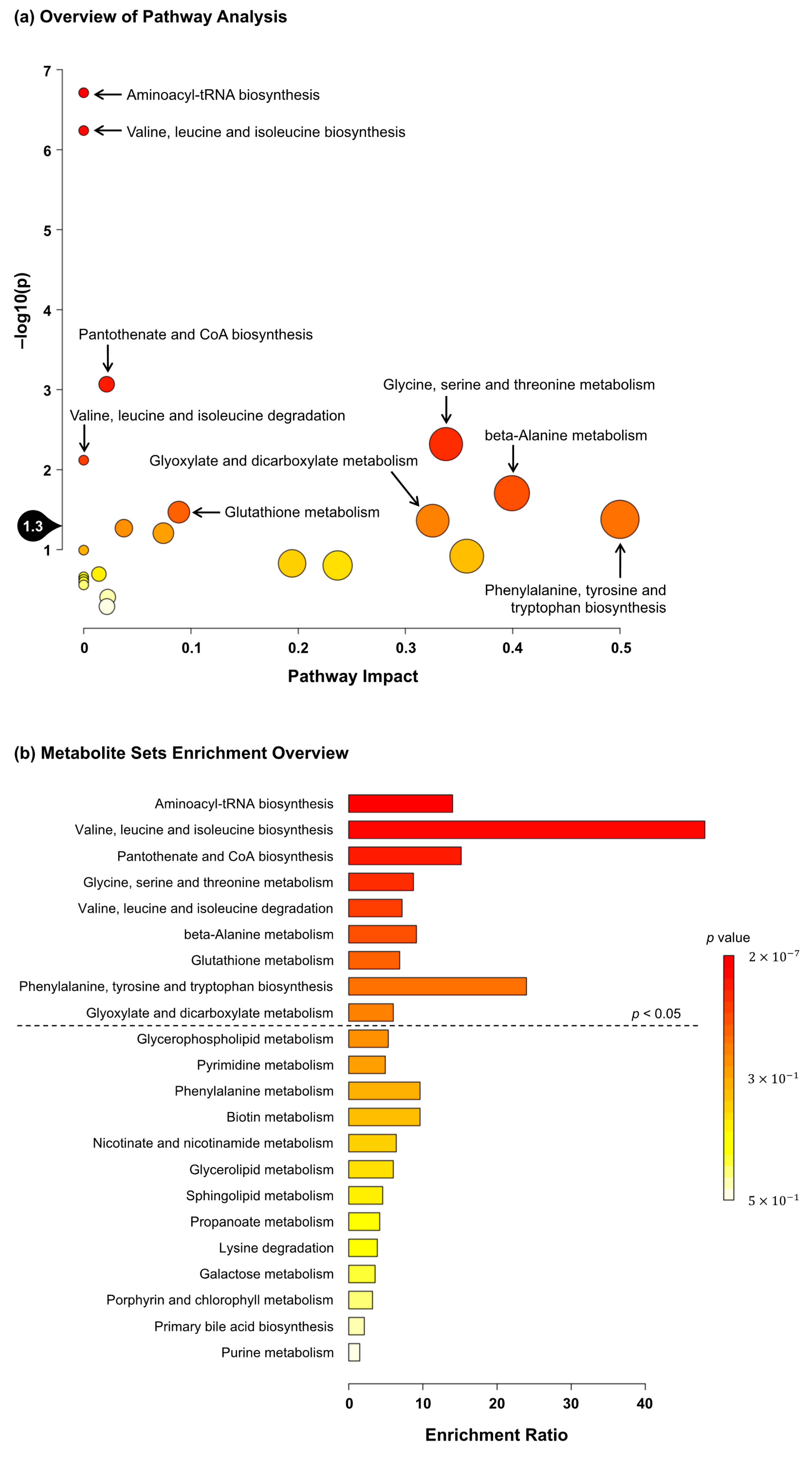
3.5. Effects of C. gracilis Feeding on Water-Soluble Metabolites in the Liver of SD Rats
3.6. Effects of C. gracilis Feeding on Water-Soluble Metabolites in the Soleus Muscle of SD Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garcia-Perez, P.; Cassani, L.; Garcia-Oliveira, P.; Xiao, J.; Simal-Gandara, J.; Prieto, M.A.; Lucini, L. Algal nutraceuticals: A perspective on metabolic diversity, current food applications, and prospects in the field of metabolomics. Food Chem. 2023, 409, 135295. [Google Scholar] [CrossRef]
- Barbosa, M.J.; Janssen, M.; Südfeld, C.; D’Adamo, S.; Wijffels, R.H. Hypes, hopes, and the way forward for microalgal biotechnology. Trends Biotechnol. 2023, 41, 452–471. [Google Scholar] [CrossRef]
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M.; et al. Current status of the algae production industry in Europe: An emerging sector of the blue bioeconomy. Front. Mar. Sci. 2020, 7, 626389. [Google Scholar] [CrossRef]
- Katayama, T.; Rahman, N.A.; Khatoon, H.; Kasan, N.A.; Nagao, N.; Yamada, Y.; Takahashi, K.; Furuya, K.; Wahid, M.E.A.; Yusoff, F.M.; et al. Bioprospecting of tropical microalgae for high-value products: n-3 polyunsaturated fatty acids and carotenoids. Aquacult. Rep. 2022, 27, 101406. [Google Scholar] [CrossRef]
- Mayer, C.; Côme, M.; Ulmann, L.; Martin, I.; Zittelli, G.C.; Faraloni, C.; Ouguerram, K.; Chénais, B.; Mimouni, V. The potential of the marine microalga Diacronema lutheri in the prevention of obesity and metabolic syndrome in high-fat-fed Wistar rats. Molecules 2022, 27, 4246. [Google Scholar] [CrossRef]
- Commission Implementing Regulation (EU) 2018/1023 of 23 July 2018 Correcting Implementing Regulation (EU) 2017/2470 Establishing the Union List of Novel Foods. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=OJ:L:2018:187:FULL&from=IT (accessed on 26 February 2023).
- Tachihana, S.; Nagao, N.; Katayama, T.; Hirahara, M.; Yusoff, F.M.; Banerjee, S.; Shariff, M.; Kurosawa, N.; Toda, T.; Furuya, K. High productivity of eicosapentaenoic acid and fucoxanthin by a marine diatom Chaetoceros gracilis in a semi-continuous culture. Front. Bioeng. Biotechnol. 2020, 8, 602721. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, L.; Hosokawa, M.; Miyashita, K. Total lipids content, lipid class and fatty acid composition of ten species of microalgae. J. Oleo Sci. 2020, 69, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J. Fucoxanthin and its metabolite fucoxanthinol in cancer prevention and treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [PubMed]
- Lau, T.Y.; Kwan, H.Y. Fucoxanthin is a potential therapeutic agent for the treatment of breast cancer. Mar. Drugs 2022, 20, 370. [Google Scholar] [CrossRef]
- Gammone, M.A.; D’Orazio, N. Anti-obesity activity of the marine carotenoid fucoxanthin. Mar. Drugs 2015, 13, 2196–2214. [Google Scholar] [CrossRef]
- Hosokawa, M.; Miyashita, T.; Nishikawa, S.; Emi, S.; Tsukui, T.; Beppu, F.; Okada, T.; Miyashita, K. Fucoxanthin regulates adipocytokine mRNA expression in white adipose tissue of diabetic/obese KK-Ay mice. Arch. Biochem. Biophys. 2010, 504, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin promotes translocation and induction of glucose transporter 4 in skeletal muscles of diabetic/obese KK-A(y) mice. Phytomedicine 2012, 19, 389–394. [Google Scholar] [CrossRef]
- Skulas-Ray, A.C.; Wilson, P.W.F.; Harris, W.S.; Brinton, E.A.; Kris-Etherton, P.M.; Richter, C.K.; Jacobson, T.A.; Engler, M.B.; Miller, M.; Robinson, J.G.; et al. Omega-3 fatty acids for the management of hypertriglyceridemia: A science advisory from the American Heart Association. Circulation 2019, 140, e673–e691. [Google Scholar] [CrossRef] [PubMed]
- Djuricic, I.; Calder, P.C. Beneficial outcomes of omega-6 and omega-3 polyunsaturated fatty acids on human health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef] [PubMed]
- The Pharmaceutical Society of Japan. 2.1.2.2 Amino Acids. In Methods of Analysis in Human Science; Kanehara & Co., Ltd.: Tokyo, Japan, 2020; pp. 211–214. [Google Scholar]
- Analytical Methods for Nutrients (March 30, 2015 Tsuuchi No.139 Attachment Consumer Affairs Agency Food Labelling Division). Available online: https://www.caa.go.jp/policies/policy/food_labeling/food_labeling_act/assets/food_labeling_cms201_220830_03.pdf (accessed on 11 March 2023).
- American Institute of Nutrition. Report of the American Institute of Nutrition ad hoc Committee on Standards for Nutritional Studies. J. Nutr. 1977, 107, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Niibo, M.; Shirouchi, B.; Umegatani, M.; Morita, Y.; Ogawa, A.; Sakai, F.; Kadooka, Y.; Sato, M. Probiotic Lactobacillus gasseri SBT2055 improves insulin secretion in a diabetic rat model. J. Dairy Sci. 2019, 102, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Shirouchi, B.; Fukuda, A.; Akasaka, T. Unlike glycerophosphocholine or choline chloride, dietary phosphatidylcholine does not increase plasma trimethylamine-N-oxide levels in Sprague-Dawley rats. Metabolites 2022, 12, 64. [Google Scholar] [CrossRef]
- Carr, T.P.; Andresen, C.J.; Rudel, L.L. Enzymatic determination of triglyceride, free cholesterol, and total cholesterol in tissue lipid extracts. Clin. Biochem. 1993, 26, 39–42. [Google Scholar] [CrossRef]
- Rouser, G.; Siakotos, A.N.; Fleischer, S. Quantitative analysis of phospholipids by thin-layer chromatography and phosphorus analysis of spots. Lipids 1966, 1, 85–86. [Google Scholar] [CrossRef]
- Lo, S.; Russell, J.C.; Taylor, A.W. Determination of glycogen in small tissue samples. J. Appl. Physiol. 1970, 28, 234–236. [Google Scholar] [CrossRef]
- Matsuoka, R.; Shirouchi, B.; Umegatani, M.; Fukuda, M.; Muto, A.; Masuda, Y.; Kunou, M.; Sato, M. Dietary egg-white protein increases body protein mass and reduces body fat mass through an acceleration of hepatic β-oxidation in rats. Br. J. Nutr. 2017, 118, 423–430. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Gylling, H.; Nissinen, M.J. The role of serum non-cholesterol sterols as surrogate markers of absolute cholesterol synthesis and absorption. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Gylling, H.; Hallikainen, M.; Simonen, P.; Miettinen, H.E.; Nissinen, M.J.; Miettinen, T.A. Serum and lipoprotein sitostanol and non-cholesterol sterols after an acute dose of plant stanol ester on its long-term consumption. Eur. J. Nutr. 2012, 51, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Bamba, T.; Shinohara, M.; Nishiumi, S.; Yoshida, M.; Fukusaki, E. Practical non-targeted gas chromatography/mass spectrometry-based metabolomics platform for metabolic phenotype analysis. J. Biosci. Bioeng. 2011, 112, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Tsugawa, H.; Ikeda, K.; Takahashi, M.; Satoh, A.; Mori, Y.; Uchino, H.; Okahashi, N.; Yamada, Y.; Tada, I.; Bonini, P.; et al. A lipidome atlas in MS-DIAL 4. Nat. Biotechnol. 2020, 38, 1159–1163. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software “EZR” for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- MetaboAnalyst. MetaboAnalyst 5.0—User-Friendly, Streamlined Metabolomics Data Analysis. Available online: https://www.metaboanalyst.ca/home.xhtml (accessed on 11 March 2023).
- Beppu, F.; Niwano, Y.; Tsukui, T.; Hosokawa, M.; Miyashita, K. Single and repeated oral dose toxicity study of fucoxanthin (FX), a marine carotenoid, in mice. J. Toxicol. Sci. 2009, 34, 501–510. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ozaki, Y.; Taminato, M.; Das, S.K.; Mizuno, M.; Yoshimura, K.; Maoka, T.; Kanazawa, K. The distribution and accumulation of fucoxanthin and its metabolites after oral administration in mice. Br. J. Nutr. 2009, 102, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Deane, C.S.; Ely, I.A.; Wilkinson, D.J.; Smith, K.; Phillips, B.E.; Atherton, P.J. Dietary protein, exercise, ageing and physical inactivity: Interactive influences on skeletal muscle proteostasis. Proc. Nutr. Soc. 2021, 80, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Hosokawa, M.; Miyashita, K.; Nishino, H.; Hashimoto, T. Effects of fucoxanthin on the inhibition of dexamethasone-induced skeletal muscle loss in mice. Nutrients 2021, 13, 1079. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.N.; Jeon, S.M.; Kim, H.J.; Lee, M.K.; Shin, S.K.; Shin, Y.C.; Park, Y.B.; Choi, M.S. Fucoxanthin supplementation improves plasma and hepatic lipid metabolism and blood glucose concentration in high-fat fed C57BL/6N mice. Chem. Biol. Interact. 2010, 186, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Beppu, F.; Hosokawa, M.; Niwano, Y.; Miyashita, K. Effects of dietary fucoxanthin on cholesterol metabolism in diabetic/obese KK-A(y) mice. Lipids Health Dis. 2012, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Innes, J.K.; Calder, P.C. The differential effects of eicosapentaenoic acid and docosahexaenoic acid on cardiometabolic risk factors: A systematic review. Int. J. Mol. Sci. 2018, 19, 532. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, S.H.; Rémond, D.; van Loon, L.J. The muscle protein synthetic response to food ingestion. Meat Sci. 2015, 109, 96–100. [Google Scholar] [CrossRef]
- Zaromskyte, G.; Prokopidis, K.; Ioannidis, T.; Tipton, K.D.; Witard, O.C. Evaluating the leucine trigger hypothesis to explain the post-prandial regulation of muscle protein synthesis in young and older adults: A systematic review. Front. Nutr. 2021, 8, 685165. [Google Scholar] [CrossRef]
- Yu, Y.C.; Han, J.M.; Kim, S. Aminoacyl-tRNA synthetases and amino acid signaling. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118889. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Calories (kcal/100 g d.w.) | 411 |
| Moisture (g/100 g d.w.) | 4.3 |
| Protein (g/100 g d.w.) | 48.4 |
| Fat (g/100 g d.w.) | 17.1 |
| Carbohydrate * (g/100 g d.w.) | 15.9 |
| Ash (g/100 g d.w.) | 14.3 |
| Sodium (mg/100 g d.w.) | 1199 |
| Salt equivalent (g/100 g d.w.) | 3.0 |
| Control | 2%Chaeto | 5%Chaeto | |
|---|---|---|---|
| Ingredients | (g/kg diet) | ||
| Sucrose | 449 | 442.10 | 431.75 |
| Casein | 200 | 190.32 | 175.80 |
| β-Cornstarch | 150 | 150 | 150 |
| Cellulose | 50 | 50 | 50 |
| Corn oil | 100 | 96.58 | 91.45 |
| C. gracilis | --- | 20 | 50 |
| Mineral mixture (AIN-76) | 35 | 35 | 35 |
| Vitamin mixture (AIN-76) | 10 | 10 | 10 |
| DL-Methionine | 3 | 3 | 3 |
| Choline bitartrate | 2 | 2 | 2 |
| Cholesterol | 1 | 1 | 1 |
| Energy (kcal/kg diet) | 3977 | 3967 | 3952 |
| Control | 2%Chaeto | 5%Chaeto | p for Trends # | |
|---|---|---|---|---|
| Initial body weight (g) | 185 ± 3 | 186 ± 3 | 186 ± 3 | N.S. |
| Final body weight (g) | 428 ± 6 | 424 ± 12 | 419 ± 4 | N.S. |
| Food intake (g/day) | 24.8 ± 0.4 | 24.5 ± 0.8 | 24.1 ± 0.6 | N.S. |
| Food efficiency (%) ‡ | 35.0 ± 0.6 | 34.5 ± 0.7 | 34.5 ± 0.3 | N.S. |
| Organ weights (g/100 g of body weight) | ||||
| Liver | 4.43 ± 0.13 | 4.44 ± 0.17 | 4.25 ± 0.11 | N.S. |
| Soleus muscle | 1.02 ± 0.03 | 1.09 ± 0.02 | 1.09 ± 0.03 | p < 0.05 |
| White adipose tissue (WAT) weights (g/100 g of body weight) | ||||
| Epididymal | 1.33 ± 0.08 | 1.22 ± 0.03 | 1.30 ± 0.16 | N.S. |
| Perirenal | 1.59 ± 0.08 | 1.56 ± 0.14 | 1.54 ± 0.30 | N.S. |
| Mesenteric | 1.57 ± 0.09 | 1.39 ± 0.16 | 1.19 ± 0.18 | N.S. |
| Abdominal † | 4.48 ± 0.18 | 4.17 ± 0.30 | 4.03 ± 0.63 | N.S. |
| Brown adipose tissue (BAT) weight (g/100 g of body weight) | ||||
| 0.147 ± 0.007 | 0.182 ± 0.017 | 0.163 ± 0.014 | N.S. | |
| Feces weights (mg feces/g food intake) | ||||
| 81.1 ± 3.1 | 91.0 ± 4.9 | 104 ± 5 * | p < 0.05 | |
| Control | 2%Chaeto | 5%Chaeto | p for Trends # | |
|---|---|---|---|---|
| Hepatic contents of lipids and glycogen (mg/g liver) | ||||
| TG | 49.8 ± 5.2 | 33.4 ± 5.6 * | 18.8 ± 2.4 * | p < 0.05 |
| T-Chol | 8.29 ± 0.61 | 6.84 ± 0.72 | 4.26 ± 0.27 * | p < 0.05 |
| PL | 27.1 ± 0.7 | 27.2 ± 2.0 | 29.3 ± 1.3 | N.S. |
| Glycogen | 33.2 ± 6.1 | 27.3 ± 4.2 | 25.6 ± 4.4 | N.S. |
| Serum biochemical parameters | ||||
| TG (mg/dL) | 181 ± 21 | 156 ± 33 | 134 ± 24 | N.S. |
| T-Chol (mg/dL) | 95.0 ± 2.6 | 124 ± 4 * | 138 ± 4 * | p < 0.05 |
| HDL Chol (mg/dL) | 43.9 ± 2.4 | 62.7 ± 5.0 * | 80.8 ± 5.1 * | p < 0.05 |
| Non-HDL Chol (mg/dL) | 51.1 ± 2.6 | 61.5 ± 5.4 | 57.7 ± 4.9 | N.S. |
| PL (mg/dL) | 193 ± 10 | 213 ± 16 | 217 ± 7 | N.S. |
| NEFAs (mmol/L) | 0.609 ± 0.055 | 0.630 ± 0.060 | 0.654 ± 0.054 | N.S. |
| Glucose (mg/dL) | 131 ± 6 | 128 ± 6 | 121 ± 4 | N.S. |
| C-peptide (pg/mL) | 229 ± 68 | 222 ± 22 | 164 ± 23 | N.S. |
| Insulin (pg/mL) | 273 ± 134 | 141 ± 29 | 148 ± 26 | N.S. |
| Adiponectin (µg/mL) | 1.69 ± 0.15 | 1.54 ± 0.16 | 1.55 ± 0.15 | N.S. |
| ALT (IU/L) | 8.42 ± 0.69 | 7.64 ± 0.68 | 7.24 ± 0.45 | N.S. |
| Control | 2%Chaeto | 5%Chaeto | p for Trends # | |
|---|---|---|---|---|
| Hepatic activities of enzymes related to fatty acid synthesis (mmol/min/mg protein) | ||||
| FAS | 10.9 ± 0.6 | 9.22 ± 0.90 | 8.49 ± 0.96 (p = 0.0865) | p < 0.05 |
| G6PDH | 35.0 ± 3.9 | 27.7 ± 8.2 | 18.9 ± 2.8 * | p < 0.05 |
| ME | 41.3 ± 4.8 | 38.3 ± 7.8 | 30.5 ± 4.3 | N.S. |
| Hepatic activities of enzyme related to fatty acid β-oxidation (mmol/min/mg protein) | ||||
| CPT | 5.34 ± 0.13 | 5.32 ± 0.23 | 5.05 ± 0.27 | N.S. |
| Cholesterol synthesis marker in serum (mg/dL) | ||||
| Lathosterol | 0.136 ± 0.028 | 0.163 ± 0.023 | 0.170 ± 0.024 | N.S. |
| Cholesterol absorption markers in serum (mg/dL) | ||||
| Campesterol | 5.41 ± 0.21 | 4.82 ± 0.14 * | 4.34 ± 0.07 * | p < 0.05 |
| β-Sitosterol | 9.83 ± 0.45 | 7.04 ± 0.33 * | 6.86 ± 0.15 * | p < 0.05 |
| Relative levels of hepatic mRNA related to fatty acid synthesis and cholesterol metabolism (arbitrary unit) | ||||
| Fasn | 1.00 ± 0.40 | 0.792 ± 0.350 | 0.452 ± 0.128 | N.S. |
| G6pd | 1.00 ± 0.08 | 1.21 ± 0.24 | 1.07 ± 0.21 | N.S. |
| Hmgcr | 1.00 ± 0.13 | 1.07 ± 0.15 | 0.971 ± 0.177 | N.S. |
| Soat1 | 1.00 ± 0.07 | 1.14 ± 0.21 | 0.786 ± 0.199 | N.S. |
| Scarb1 | 1.00 ± 0.06 | 0.886 ± 0.049 | 0.683 ± 0.105 * | p < 0.05 |
| Abca1 | 1.00 ± 0.15 | 1.20 ± 0.13 | 1.06 ± 0.20 | N.S. |
| Control | 5%Chaeto | |
|---|---|---|
| Levels of water-soluble metabolites in the liver (arbitrary unit #) | ||
| Glycerol | 402 ± 22 | 239 ± 40 * |
| Hypotaurine | 32.5 ± 5.3 | 10.6 ± 1.6 † |
| Inositol | 24.7 ± 1.3 | 17.6 ± 1.3 * |
| Levels of water-soluble metabolites in the soleus muscle (arbitrary unit #) | ||
| 2-Aminoethnol | 10.3 ± 0.7 | 17.8 ± 0.8 * |
| 3-Hydroxypyruvate | 194 ± 17 | 283 ± 18 * |
| β-Alanine | 23.0 ± 3.0 | 54.8 ± 7.2 * |
| Cadaverine | 17.6 ± 1.8 | 26.2 ± 3.5 * |
| Creatinine | 52.7 ± 3.6 | 76.2 ± 3.7 * |
| Glycerol | 189 ± 16 | 275 ± 14 * |
| Glycine | 500 ± 69 | 727 ± 49 * |
| Hypoxanthine | 39.5 ± 6.5 | 63.8 ± 8.6 * |
| Iminodiacetate | 14.0 ± 2.2 | 28.5 ± 4.3 * |
| Isoleucine | 72.6 ± 3.1 | 129 ± 14 † |
| Leucine | 142 ± 9 | 237 ± 27 † |
| Lysine | 154 ± 16 | 282 ± 45 * |
| Nicotinamide | 31.0 ± 1.4 | 35.9 ± 1.6 * |
| O-Phosphoethanolamine | 5.34 ± 0.41 | 7.59 ± 0.55 * |
| Oxalate | 17.6 ± 1.4 | 13.1 ± 1.5 * |
| Phenylalanine | 11.2 ± 1.2 | 23.4 ± 3.5 * |
| Serine | 24.6 ± 2.5 | 51.0 ± 5.7 * |
| Threonine | 23.4 ± 2.5 | 39.0 ± 3.7 * |
| Uracil | 4.42 ± 0.61 | 7.11 ± 0.98 * |
| Valine | 81.3 ± 3.3 | 142 ± 19 † |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shirouchi, B.; Kawahara, Y.; Kutsuna, Y.; Higuchi, M.; Okumura, M.; Mitsuta, S.; Nagao, N.; Tanaka, K. Oral Administration of Chaetoceros gracilis—A Marine Microalga—Alleviates Hepatic Lipid Accumulation in Rats Fed a High-Sucrose and Cholesterol-Containing Diet. Metabolites 2023, 13, 436. https://doi.org/10.3390/metabo13030436
Shirouchi B, Kawahara Y, Kutsuna Y, Higuchi M, Okumura M, Mitsuta S, Nagao N, Tanaka K. Oral Administration of Chaetoceros gracilis—A Marine Microalga—Alleviates Hepatic Lipid Accumulation in Rats Fed a High-Sucrose and Cholesterol-Containing Diet. Metabolites. 2023; 13(3):436. https://doi.org/10.3390/metabo13030436
Chicago/Turabian StyleShirouchi, Bungo, Yuri Kawahara, Yuka Kutsuna, Mina Higuchi, Mai Okumura, Sarasa Mitsuta, Norio Nagao, and Kazunari Tanaka. 2023. "Oral Administration of Chaetoceros gracilis—A Marine Microalga—Alleviates Hepatic Lipid Accumulation in Rats Fed a High-Sucrose and Cholesterol-Containing Diet" Metabolites 13, no. 3: 436. https://doi.org/10.3390/metabo13030436
APA StyleShirouchi, B., Kawahara, Y., Kutsuna, Y., Higuchi, M., Okumura, M., Mitsuta, S., Nagao, N., & Tanaka, K. (2023). Oral Administration of Chaetoceros gracilis—A Marine Microalga—Alleviates Hepatic Lipid Accumulation in Rats Fed a High-Sucrose and Cholesterol-Containing Diet. Metabolites, 13(3), 436. https://doi.org/10.3390/metabo13030436