
Antioxidant Capacity, Antitumor Activity and Metabolomic Profile of a Beetroot Peel Flour

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methodology
2.1. Flour Development
2.2. Antioxidant Activity
2.2.1. Total Phenolic Compounds (TPCs)
2.2.2. ABTS Radical Scavenging
2.2.3. DPPH Radical Scavenging
2.2.4. Ferric Iron Reducing Antioxidant Parameter (FRAP)
2.2.5. Oxygen Radical Absorbance Capacity (ORAC)
2.3. Influence on Cancer Cells
2.3.1. BPF Extract Preparation
2.3.2. Cell Culture
2.3.3. Cell Viability
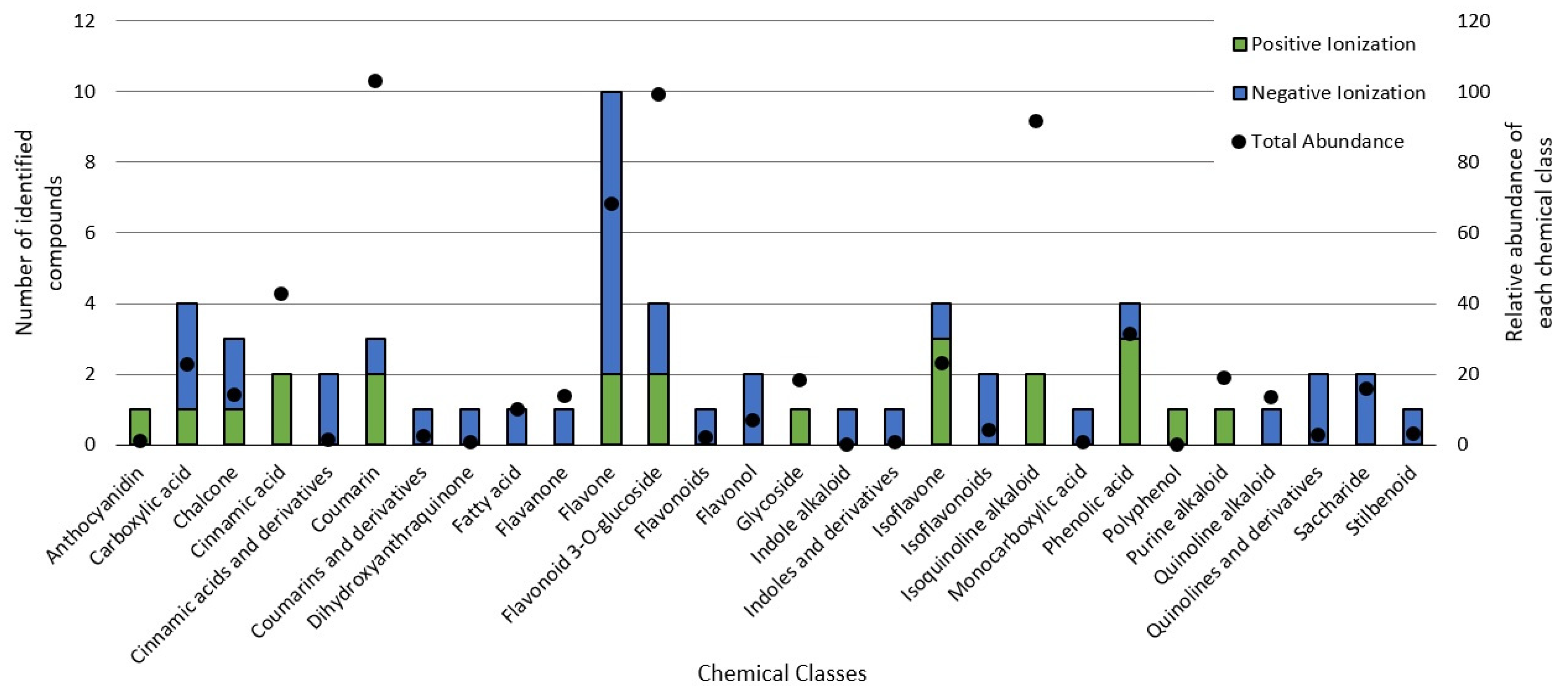
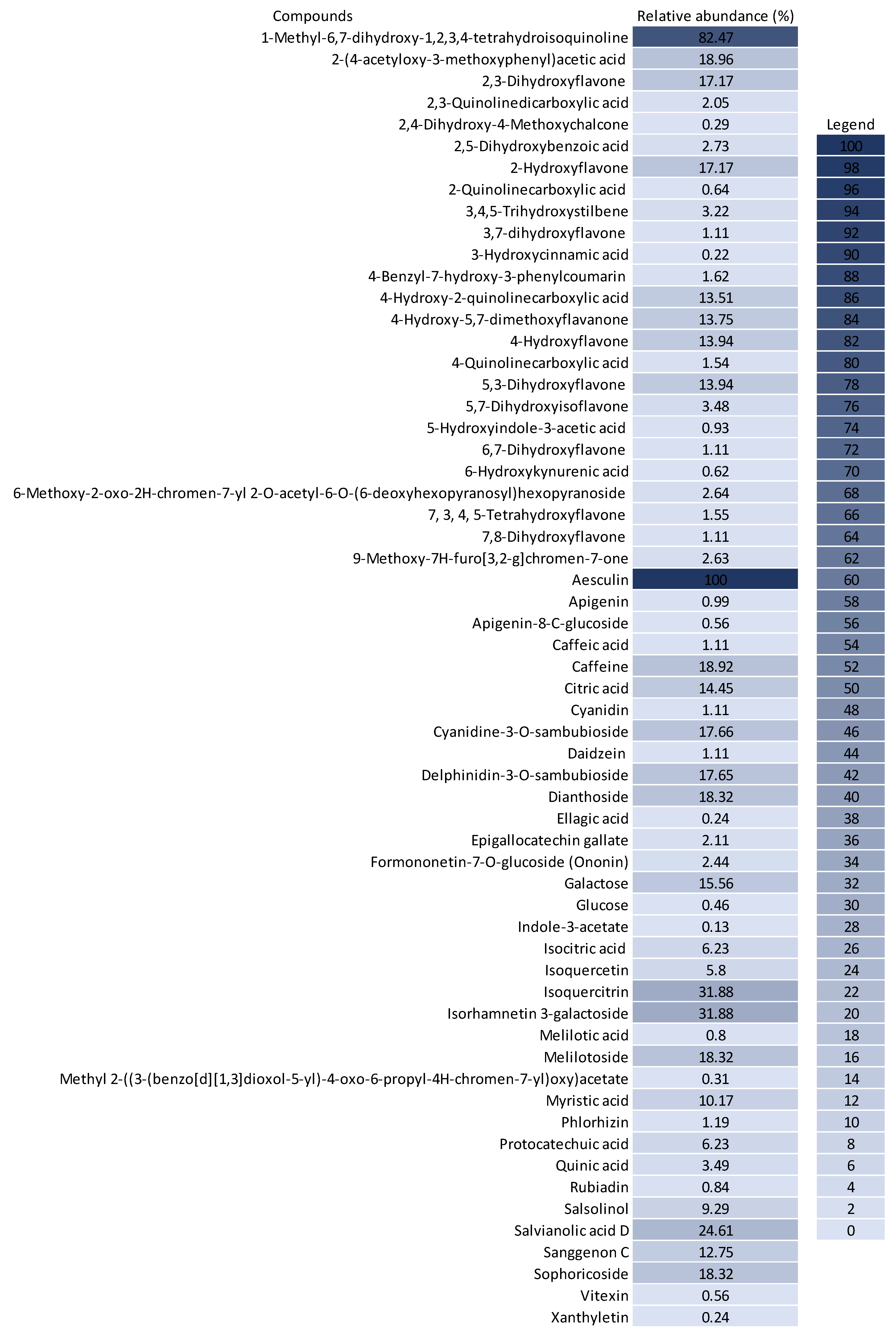
2.4. Qualitative Chemical Profile
2.5. Statistical Analysis
3. Results and Discussion
3.1. Antioxidant Activity of BPF Extracts
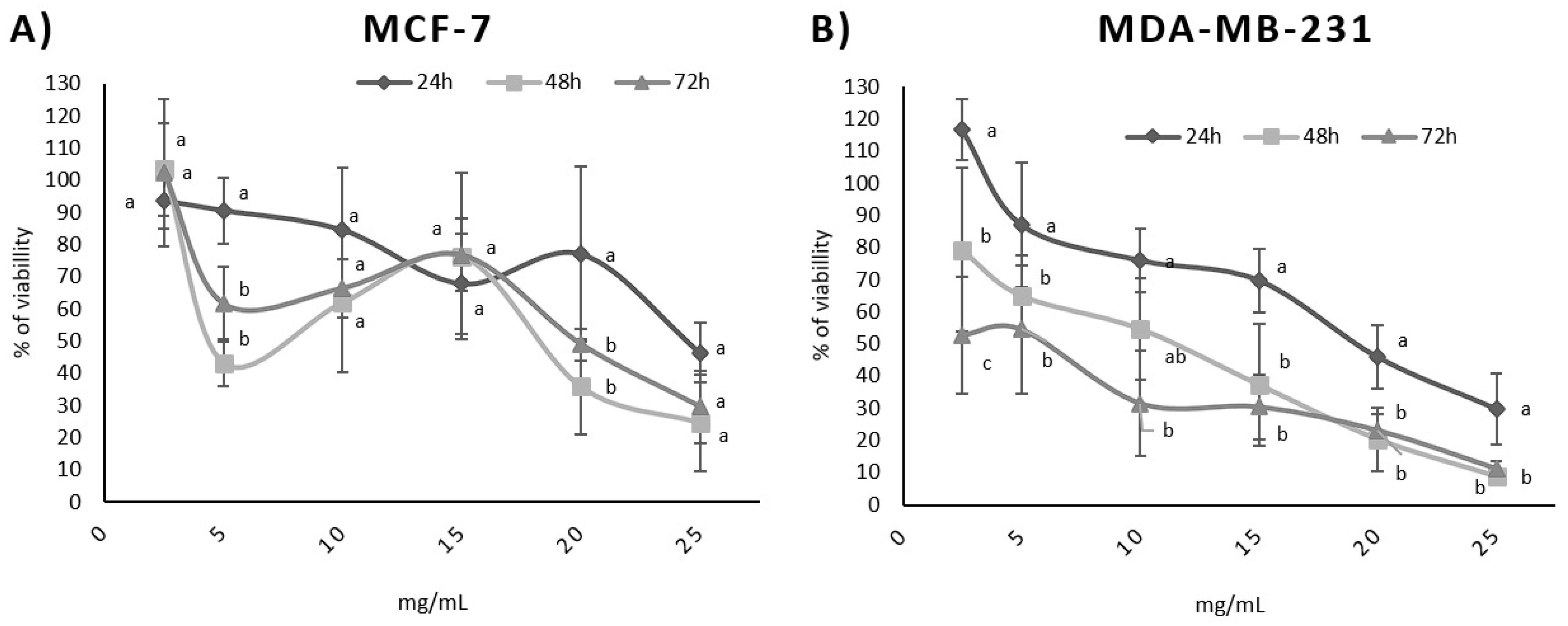
3.2. Influence of BPFw on Breast Cancer Cell Survival
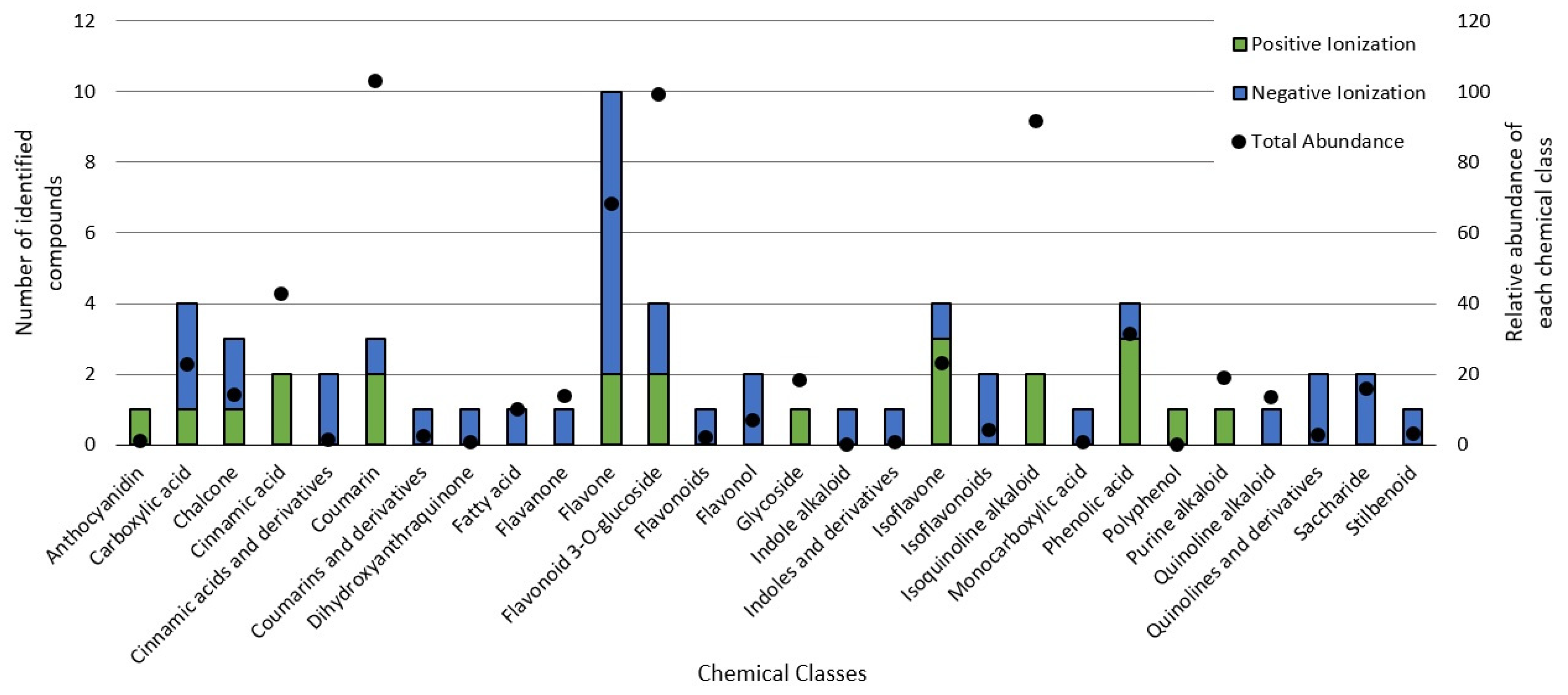
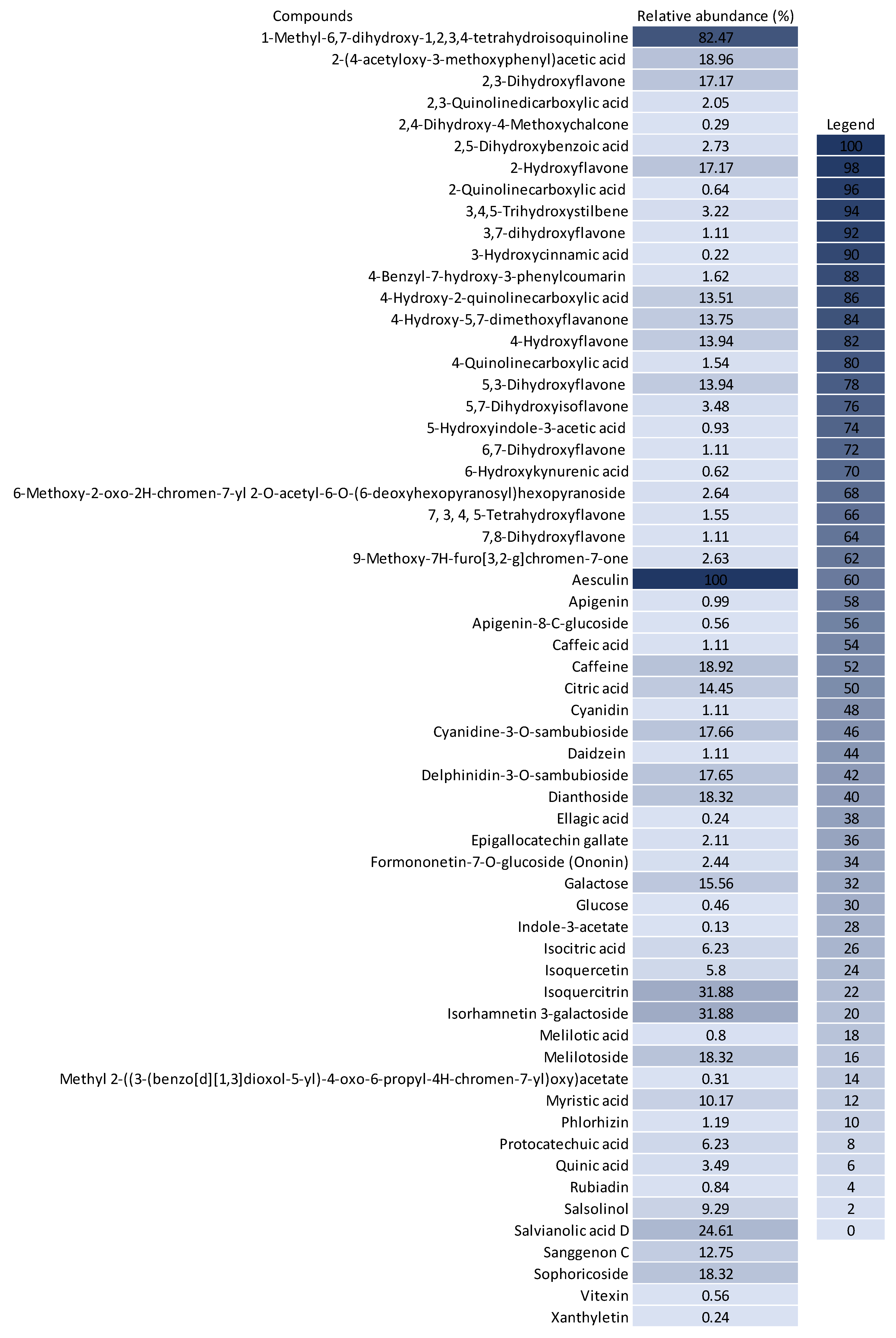
3.3. Chemical Profile of BPF Extracts and Their Influence on Cancer Cell Metabolism

{kind=link}
{kind=link}
{kind=link}
| Compound | Formula | [M + H]+ Theoretical | [M + H]+ Observed | Mass Accuracy (ppm) | Molecular Structure | Reference |
|---|---|---|---|---|---|---|
| Betacyanins | ||||||
| Betanin | C24H26N2O13 | 551.1508 | 551.1500 | −1.38 | 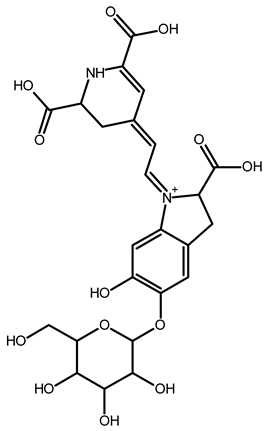 | [49] |
| 15,17-bidecarboxy-betanin | C16H17N2O4 | 301.1194 | 301.1185 | 0.72 | 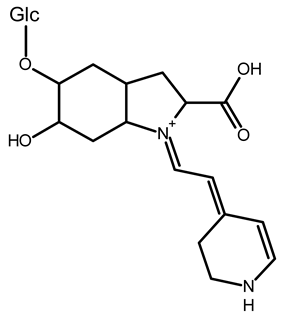 | [11] |
| 2,15-bidecarboxy-xanbetanin | C16H15N2O4 | 461.1547 | 461.1544 | −2.99 |  | [11] |
| 2,15,17-tridecarboxybidehydro-amaranthin | C27H30N2O13 | 591.1821 | 591.1832 | 1.86 | 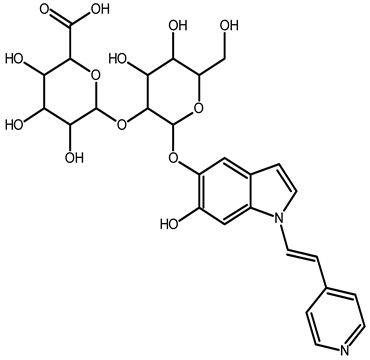 | [49] |
| 6′-feruloyl-betanin | C34H35N2O16+ | 727.1981 | 727.1962 | −2.61 | 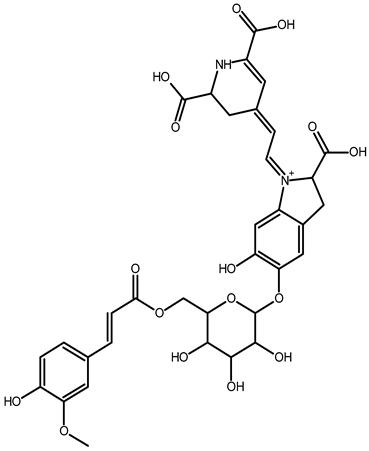 | [52] |
| Betaxanthins | ||||||
| Ethanolamine-betaxanthin | C12H14N2O5 | 255.0970 | 255.0988 | 7.05 | 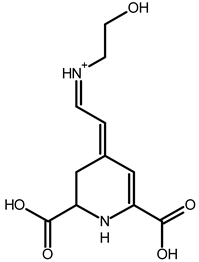 | [49,53] |
| Threonine-betaxanthin | C13H16N2O7 | 313.1022 | 313.1027 | 1.59 | 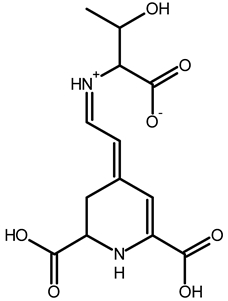 | [49,53] |
| Vulgaxanthin I (glutamine-betaxanthin) | C14H17N387 | 340.1134 | 340.1135 | 0.29 | 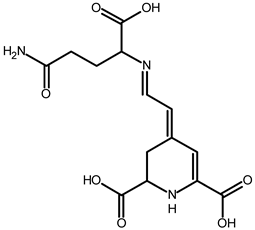 | [49] |
| Vulgaxanthin IV (leucine-betaxanthin) | C15H20N2O6 | 325.1387 | 325.1389 | 0.61 | 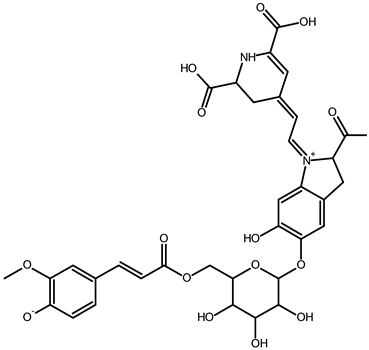 | [49] |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Babarykin, D.; Smirnova, G.; Pundinsh, I.; Vasiljeva, S.; Krumina, G.; Agejchenko, V. Red Beet (Beta vulgaris) Impact on Human Health. J. Biosci. Med. 2019, 07, 61–79. [Google Scholar] [CrossRef]
- Mancini, M.C.S.; Ponte, L.G.S.; Silva, C.H.R.; Fagundes, I.; Pavan, I.C.B.; Romeiro, S.A.; da Silva, L.G.S.; Morelli, A.P.; Rostagno, M.A.; Simabuco, F.M.; et al. Beetroot and Leaf Extracts Present Protective Effects against Prostate Cancer Cells, Inhibiting Cell Proliferation, Migration, and Growth Signaling Pathways. Phyther. Res. 2021, 35, 5241–5258. [Google Scholar] [CrossRef]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative Stress and Cancer: An Overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, 28. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Yin, Z.; Yang, Y.; Guo, T.; Veeraraghavan, V.P.; Wang, X. Potential Chemotherapeutic Effect of Betalain against Human Non-Small Cell Lung Cancer through PI3K/Akt/MTOR Signaling Pathway. Environ. Toxicol. 2021, 36, 1011–1020. [Google Scholar] [CrossRef]
- Šeremet, D.; Durgo, K.; Jokić, S.; Hudek, A.; Cebin, A.V.; Mandura, A.; Jurasović, J.; Komes, D. Valorization of Banana and Red Beetroot Peels: Determination of Basic Macrocomponent Composition, Application of Novel Extraction Methodology and Assessment of Biological Activity in Vitro. Sustainability 2020, 12, 4539. [Google Scholar] [CrossRef]
- Slatnar, A.; Stampar, F.; Veberic, R.; Jakopic, J. HPLC-MSn Identification of Betalain Profile of Different Beetroot (Beta vulgaris L. Ssp. Vulgaris) Parts and Cultivars. J. Food Sci. 2015, 80, 1952–1958. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Megahed, B.M.H.; Gamal, M.; Safwat, G. Evaluation of Some Chemical Constituents, Antioxidant, Antibacterial and Anticancer Activities of Beta vulgaris L. Root. Fresenius Environ. Bull. 2018, 27, 6369–6378. [Google Scholar]
- Lazăr, S.; Constantin, O.E.; Stănciuc, N.; Aprodu, I.; Croitoru, C.; Râpeanu, G. Optimization of Betalain Pigments Extraction Using Beetroot By-Products as a Valuable Source. Inventions 2021, 6, 50. [Google Scholar] [CrossRef]
- Sutor-świeży, K.; Antonik, M.; Proszek, J.; Nemzer, B.; Pietrzkowski, Z.; Popenda, Ł.; Świergosz, T.; Wybraniec, S. Dehydrogenation of Betacyanins in Heated Betalain-Rich Extracts of Red Beet (Beta vulgaris L.). Int. J. Mol. Sci. 2022, 23, 1245. [Google Scholar] [CrossRef]
- Sevastre, A.S.; Manea, E.V.; Popescu, O.S.; Tache, D.E.; Danoiu, S.; Sfredel, V.; Tataranu, L.G.; Dricu, A. Intracellular Pathways and Mechanisms of Colored Secondary Metabolites in Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 9943. [Google Scholar] [CrossRef] [PubMed]
- Parveen, A.; Akash, M.S.H.; Rehman, K.; Kyunn, W.W. Anticancer Activities of Medicinal Plants: Modulation of P53 Expression and Induction of Apoptosis. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 257–271. [Google Scholar] [CrossRef]
- United Nations. Sustainable Development Goals. Available online: https://sdgs.un.org/ (accessed on 22 November 2022).
- Akan, S.; Tuna Gunes, N.; Erkan, M. Red Beetroot: Health Benefits, Production Techniques, and Quality Maintaining for Food Industry. J. Food Process. Preserv. 2021, 45, e15781. [Google Scholar] [CrossRef]
- Larrosa, A.P.Q.; Otero, D.M. Flour Made from Fruit By-Products: Characteristics, Processing Conditions, and Applications. J. Food Process. Preserv. 2021, 45, e15398. [Google Scholar] [CrossRef]
- Salamatullah, A.M.; Hayat, K.; Alkaltham, M.S.; Ahmed, M.A.; Arzoo, S.; Husain, F.M.; Al-Dossari, A.M.; Shamlan, G.; Al-Harbi, L.N. Bioactive and Antimicrobial Properties of Oven-Dried Beetroot (Pulp and Peel) Using Different Solvents. Processes 2021, 9, 588. [Google Scholar] [CrossRef]
- Abdo, E.; El-Sohaimy, S.; Shaltout, O.; Abdalla, A.; Zeitoun, A. Nutritional Evaluation of Beetroots (Beta vulgaris L.) and Its Potential Application in a Functional Beverage. Plants 2020, 9, 1752. [Google Scholar] [CrossRef]
- Parveen, A.; Akash, M.S.H.; Rehman, K.; Kyunn, W.W. Recent Investigations for Discovery of Natural Antioxidants: A Comprehensive Review. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Abreu, J.; Quintino, I.; Pascoal, G.; Postingher, B.; Cadena, R.; Teodoro, A. Antioxidant Capacity, Phenolic Compound Content and Sensory Properties of Cookies Produced from Organic Grape Peel (Vitis labrusca) Flour. Int. J. Food Sci. Technol. 2019, 54, 1215–1224. [Google Scholar] [CrossRef]
- Rufino, M.d.S.; Alves, R.E.; De Brito, E.S.; De Morais, S.M.; Sampaio, C.D.G.; Saura-Calixto, F.D. Comunicado Técnico 128—Metodologia Científica: Determinação Da Atividade Antioxidante Total Em Frutas Pela Captura Do Radical Livre ABTS +. Fortaleza, Ceará, 2007. ISSN 1679-6535. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/CNPAT/10225/1/Cot_128.pdf (accessed on 22 November 2022).
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC Assays for Estimating Antioxidant Activity from Guava Fruit Extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Prior, R.L.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for Hydrophilic and Lipophilic Antioxidant Capacity (Oxygen Radical Absorbance Capacity (ORACFL)) of Plasma and Other Biological and Food Samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Antonio, A.D.S.; dos Santos, G.R.C.; Pereira, H.M.G.; da Veiga-Junior, V.F.; Wiedemann, L.S.M. Chemophenetic Study of Ocotea Canaliculata (Lauraceae) by UHPLC–HRMS and GNPS. Nat. Prod. Res. 2020, 36, 984–988. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tong, G.H.; Wei, X.X.; Chen, H.Y.; Liang, T.; Tang, H.P.; Wu, C.A.; Wen, G.M.; Yang, W.K.; Liang, L.; et al. Identification of Five Cytotoxicity-Related Genes Involved in the Progression of Triple-Negative Breast Cancer. Front. Genet. 2022, 12, 2312. [Google Scholar] [CrossRef] [PubMed]
- Maatouk, M.; Ibrahim, A. Small and Equipped: The Rich Repertoire of Antibiotic. mSystems 2021, 6, e00898-21. [Google Scholar] [CrossRef] [PubMed]
- Zin, M.M.; Bánvölgyi, S. Emerging Technology Approach for Extractability and Stability of Betalains from the Peel of Beetroot (Beta vulgaris L.). Biomass Convers. Biorefinery 2021. [Google Scholar] [CrossRef]
- Ceclu, L.; Nistor, O.-V. Red Beetroot: Composition and Health Effects—A Review. J. Nutr. Med. Diet Care 2020, 6, 043. [Google Scholar] [CrossRef]
- Chhikara, N.; Kushwaha, K.; Sharma, P.; Gat, Y.; Panghal, A. Bioactive Compounds of Beetroot and Utilization in Food Processing Industry: A Critical Review. Food Chem. 2019, 272, 192–200. [Google Scholar] [CrossRef]
- Kumorkiewicz, A.; Sutor, K.; Nemzer, B.; Pietrzkowski, Z.; Wybraniec, S. Thermal Decarboxylation of Betacyanins in Red Beet Betalain-Rich Extract. Polish J. Food Nutr. Sci. 2020, 70, 7–14. [Google Scholar] [CrossRef]
- Lefebvre, T.; Destandau, E.; Lesellier, E. Selective Extraction of Bioactive Compounds from Plants Using Recent Extraction Techniques: A Review. J. Chromatogr. A 2021, 1635, 461770. [Google Scholar] [CrossRef]
- Lima, L.G.B.; Montenegro, J.; de Abreu, J.P.; Santos, M.C.B.; do Nascimento, T.P.; Santos, M.d.S.; Ferreira, A.G.; Cameron, L.C.; Ferreira, M.S.L.; Teodoro, A.J. Metabolite Profiling by UPLC-MSE, NMR, and Antioxidant Properties of Amazonian Fruits: Mamey Apple (Mammea Americana), Camapu (Physalis Angulata), and Uxi (Endopleura Uchi). Molecules 2020, 25, 342. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Comşa, Ş.; Cîmpean, A.M.; Raica, M. The Story of MCF-7 Breast Cancer Cell Line: 40 Years of Experience in Research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar]
- Wright, L.; Frye, J.; Gorti, B.; Timmermann, B.; Funk, J. Bioactivity of Turmeric-Derived Curcuminoids and Related Metabolites in Breast Cancer. Curr. Pharm. Des. 2013, 19, 6218–6225. [Google Scholar] [CrossRef]
- Figueroa-Magalhães, M.C.; Jelovac, D.; Connolly, R.M.; Wolff, A.C. Treatment of HER2-Positive Breast Cancer. Breast 2014, 23, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Tsatsakis, A.M.; Vassilopoulou, L.; Kovatsi, L.; Tsitsimpikou, C.; Karamanou, M.; Leon, G.; Liesivuori, J.; Hayes, A.W.; Spandidos, D.A. The Dose Response Principle from Philosophy to Modern Toxicology: The Impact of Ancient Philosophy and Medicine in Modern Toxicology Science. Toxicol. Rep. 2018, 5, 1107–1113. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Kim, M.Y.; Bo, H.H.; Choi, E.O.; Kwon, D.H.; Kim, H.J.; Ahn, K.I.; Ji, S.Y.; Jeong, J.W.; Park, S.H.; Hong, S.H.; et al. Induction of Apoptosis by Citrus Unshiu Peel in Human Breast Cancer MCF-7 Cells: Involvement of ROS-Dependent Activation of AMPK. Biol. Pharm. Bull. 2018, 41, 713–721. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-MTOR Pathway and Prostate Cancer: At the Crossroads of AR, MAPK, and WNT Signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Dong, C.; Wu, J.; Chen, Y.; Nie, J.; Chen, C. Activation of PI3K/AKT/MTOR Pathway Causes Drug Resistance in Breast Cancer. Front. Pharmacol. 2021, 12, 628690. [Google Scholar] [CrossRef]
- Sanchez, C.G.; Ma, C.X.; Crowder, R.J.; Guintoli, T.; Phommaly, C.; Gao, F.; Lin, L.; Ellis, M.J. Preclinical Modeling of Combined Phosphatidylinositol-3-Kinase Inhibition with Endocrine Therapy for Estrogen Receptor-Positive Breast Cancer. Breast Cancer Res. 2011, 13, R21. [Google Scholar] [CrossRef]
- Carneiro, B.A.; El-Deiry, W.S. Targeting Apoptosis in Cancer Therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Sawicki, T.; Bączek, N.; Wiczkowski, W. Betalain Profile, Content and Antioxidant Capacity of Red Beetroot Dependent on the Genotype and Root Part. J. Funct. Foods 2016, 27, 249–261. [Google Scholar] [CrossRef]
- Rahimi, P.; Abedimanesh, S.; Mesbah-Namin, S.A.; Ostadrahimi, A. Betalains, the Nature-Inspired Pigments, in Health and Diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 2949–2978. [Google Scholar] [CrossRef]
- Xie, G.-R.; Chen, H.-J. Comprehensive Betalain Profiling of Djulis (Chenopodium formosanum) Cultivars Using HPLC-Q-Orbitrap High-Resolution Mass Spectrometry. J. Agric. Food Chem. 2021, 69, 15699–15715. [Google Scholar] [CrossRef] [PubMed]
- Otálora, C.M.; Bonifazi, E.L.; Fissore, E.N.; Basanta, M.F.; Gerschenson, L.N. Thermal Stability of Betalains in By-Products of the Blanching and Cutting of Beta vulgaris L. Var Conditiva. Pol. J. Food Nutr. Sci. 2020, 70, 15–24. [Google Scholar] [CrossRef]
- Kazimierczak, R.; Hallmann, E.; Lipowski, J.; Drela, N.; Kowalik, A.; Püssa, T.; Matt, D.; Luik, A.; Gozdowskif, D.; Rembiałkowska, E. Beetroot (Beta vulgaris L.) and Naturally Fermented Beetroot Juices from Organic and Conventional Production: Metabolomics, Antioxidant Levels and Anticancer Activity. J. Sci. Food Agric. 2014, 94, 2618–2629. [Google Scholar] [CrossRef]
- Wang, J.; Jayaprakasha, G.K.; Patil, B.S. UPLC-QTOF-MS Fingerprinting Combined with Chemometrics to Assess the Solvent Extraction Efficiency, Phytochemical Variation, and Antioxidant Activities of Beta vulgaris L. J. Food Drug Anal. 2020, 28, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Kugler, F.; Graneis, S.; Stintzing, F.C.; Carle, R. Studies on Betaxanthin Profiles of Vegetables and Fruits from the Chenopodiaceae and Cactaceae. Z. Naturforsch.—Sect. C J. Biosci. 2007, 62, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Pan, J.; Wang, Y.; Lubet, R.; You, M. Beetroot Red (Betanin) Inhibits Vinyl Carbamate- and Benzo(a)Pyrene-Induced Lung Tumorigenesis through Apoptosis. Mol. Carcinog. 2013, 52, 686–691. [Google Scholar] [CrossRef]
- Lechner, J.F.; Stoner, G.D. Red Beetroot and Betalains as Cancer Chemopreventative Agents. Molecules 2019, 24, 1602. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.L.; Hamid, S.B.S. Beetroot as a Potential Functional Food for Cancer Chemoprevention, a Narrative Review. J. Cancer Prev. 2021, 26, 1–17. [Google Scholar] [CrossRef]
- Naselli, F.; Belshaw, N.J.; Gentile, C.; Tutone, M.; Tesoriere, L.; Livrea, M.A.; Caradonna, F. Phytochemical Indicaxanthin Inhibits Colon Cancer Cell Growth and Affects the DNA Methylation Status by Influencing Epigenetically Modifying Enzyme Expression and Activity. J. Nutrigenet. Nutrigenom. 2015, 8, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Kim, B. Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review. Nutrients 2018, 10, 1021. [Google Scholar] [CrossRef]
- Valentová, K.; Vrba, J.; Bancířová, M.; Ulrichová, J.; Křen, V. Isoquercitrin: Pharmacology, Toxicology, and Metabolism. Food Chem. Toxicol. 2014, 68, 267–282. [Google Scholar] [CrossRef]
- Liskova, A.; Koklesova, L.; Samec, M.; Smejkal, K.; Samuel, S.M.; Varghese, E.; Abotaleb, M.; Biringer, K.; Kudela, E.; Danko, J.; et al. Flavonoids in Cancer Metastasis. Cancers 2020, 12, 1498. [Google Scholar] [CrossRef]
- Kaneko, T.; Tahara, S.; Takabayashi, F. Inhibitory Effect of Natural Coumarin Compounds, Esculetin and Esculin, on Oxidative DNA Damage and Formation of Aberrant Crypt Foci and Tumors Induced by 1,2-Dimethylhydrazine in Rat Colons. Biol. Pharm. Bull. 2007, 30, 2052–2057. [Google Scholar] [CrossRef]
- Rashmi, R.; Prakash, N.; Swamy, H.D.N.; Swamy, M.N. Cytotoxicity Screening of Esculin with or without Piperine. J. Entomol. Zool. Stud. 2020, 8, 230–233. [Google Scholar]
- Storch, A.; Kaftan, A.; Burkhardt, K.; Schwarz, J. 1-Methyl-6,7-Dihydroxy-1,2,3,4-Tetrahydroisoquinoline (Salsolinol) Is Toxic to Dopaminergic Neuroblastoma SH-SY5Y Cells via Impairment of Cellular Energy Metabolism. Brain Res. 2000, 855, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ł, M.K.-; Latacz, G.; Martyniak, A.; Bugajski, A.; Kie, K.; Gil, K. Salsolinol—Neurotoxic or Neuroprotective? Neurotox. Res. 2020, 37, 286–297. [Google Scholar]
- Maiti, M.; Kumar, G.S. Polymorphic Nucleic Acid Binding of Bioactive Isoquinoline Alkaloids and Their Role in Cancer. J. Nucleic Acids 2010, 2010, 593408. [Google Scholar] [CrossRef] [PubMed]
- Yun, D.; Yoon, S.Y.; Park, S.J.; Park, Y.J. The Anticancer Effect of Natural Plant Alkaloid Isoquinolines. Int. J. Mol. Sci. 2021, 22, 1653. [Google Scholar] [CrossRef]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic Acids: Potential Source of Natural Drugs for the Treatment of Fibrosis Disease and Cancer. Front. Pharmacol. 2019, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Gholami, M.H.; Zabolian, A.; Saleki, H.; Farahani, M.V.; Hamzehlou, S.; Far, F.B.; Sharifzadeh, S.O.; Samarghandian, S.; Khan, H.; et al. Caffeic Acid and Its Derivatives as Potential Modulators of Oncogenic Molecular Pathways: New Hope in the Fight against Cancer. Pharmacol. Res. 2021, 171, 105759. [Google Scholar] [CrossRef]
- Pelinson, L.P.; Assmann, C.E.; Palma, T.V.; da Cruz, I.B.M.; Pillat, M.M.; Mânica, A.; Stefanello, N.; Weis, G.C.C.; de Oliveira Alves, A.; de Andrade, C.M.; et al. Antiproliferative and Apoptotic Effects of Caffeic Acid on SK-Mel-28 Human Melanoma Cancer Cells. Mol. Biol. Rep. 2019, 46, 2085–2092. [Google Scholar] [CrossRef]
- Rosendahl, A.H.; Perks, C.M.; Zeng, L.; Markkula, A.; Simonsson, M.; Rose, C.; Ingvar, C.; Holly, J.M.P.; Jernström, H. Caffeine and Caffeic Acid Inhibit Growth and Modify Estrogen Receptor and Insulin-like Growth Factor i Receptor Levels in Human Breast Cancer. Clin. Cancer Res. 2015, 21, 1877–1887. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, Y.; Hu, Z.; Wu, S.; Jin, C. Beetroot as a Functional Food with Huge Health Benefits: Antioxidant, Antitumor, Physical Function, and Chronic Metabolomics Activity. Food Sci. Nutr. 2021, 9, 6406–6420. [Google Scholar] [CrossRef]
- Singh, V.; Kumar, K.; Purohit, D.; Verma, R.; Pandey, P.; Bhatia, S.; Malik, V.; Mittal, V.; Rahman, M.H.; Albadrani, G.M.; et al. Exploration of Therapeutic Applicability and Different Signaling Mechanism of Various Phytopharmacological Agents for Treatment of Breast Cancer. Biomed. Pharmacother. 2021, 139, 111584. [Google Scholar] [CrossRef]

| Ionization Mode | Positive | Negative |
|---|---|---|
| Step 1. Mass detection | ||
| Mass detector | Exact mass | |
| MS level | 1 | |
| Noise level | 4.00 × 106 | 5.00 × 105 |
| MS level | 2 | |
| Noise level | 1.00 × 104 | 1.00 × 104 |
| Step 2. ADAP chromatogram builder | ||
| Min group size of scan | 5 | |
| Group intensity threshold | 1.20 × 107 | 1.00 × 104 |
| Min highest intensity | 6.00 × 104 | 1.00 × 104 |
| m/z tolerance | 0.0 Da or 10 ppm | |
| Step 3. Chromatogram deconvolution | ||
| Algorithm | baseline cut-off | |
| Min. peak height | 1.00 × 106 | 1.00 × 105 |
| Peak duration (min) | 0 to 4 | |
| Baseline level | 1.20 × 105 | 4.00 × 105 |
| m/z range for MS2 scan pairing (Da) | 0.01 | |
| m/z range for MS2 scan pairing (min) | 0.2 | |
| m/z center calculation | Average | |
| Step 4. Deisotope | ||
| m/z tolerance | 0.0 Da or 10 ppm | |
| Retention time tolerance (%) | 30 | |
| Maximum charge | 3 | |
| Representative isotope | most intense | |
| Extract | In Vitro Analysis | ||||
|---|---|---|---|---|---|
| TPC (mg GAE/100 g) | DPPH (µmol T/100 g) | ABTS (µmol T/100 g) | FRAP (µM FS/g) | ORAC (µmol T/100 g) | |
| BPFw | 301.64 ± 0.20 a | 259.72 ± 17.26 a | 7692.31 ± 0.01 a | 29.72 ± 0.04 a | 3032.78 ± 55.00 a |
| BPFe | 246.99 ± 0.03 b | 299.37 ± 54.71 b | 9081.50 ± 0.03 b | 96.98 ± 0.03 b | 462.50 ± 89.00 b |
| Treatment Time | LC50 (mg/mL) | |
|---|---|---|
| MCF-7 | MDA-MB-231 | |
| 24 h | 22.57 ± 3.12 a* | 20.12 ± 3.42 a |
| 48 h | 16.64 ± 6.72 b* | 13.21 ± 1.90 b |
| 72 h | 20.81 ± 3.53 a* | 7.87 ± 7.13 c |
| Cell Line | Dose-Dependent Correlation for Each Exposure Time (p Values) | Time-Dependent Correlation (p Values) | ||
|---|---|---|---|---|
| 24 h | 48 h | 72 h | ||
| MCF-7 | −0.904 | −0.700 | −0.819 | −0.292 |
| MDA-MB-231 | −0.969 | −0.996 | −0.968 | −0.997 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coimbra, P.P.S.; Silva-e-Silva, A.C.A.G.d.; Antonio, A.d.S.; Pereira, H.M.G.; Veiga-Junior, V.F.d.; Felzenszwalb, I.; Araujo-Lima, C.F.; Teodoro, A.J. Antioxidant Capacity, Antitumor Activity and Metabolomic Profile of a Beetroot Peel Flour. Metabolites 2023, 13, 277. https://doi.org/10.3390/metabo13020277
Coimbra PPS, Silva-e-Silva ACAGd, Antonio AdS, Pereira HMG, Veiga-Junior VFd, Felzenszwalb I, Araujo-Lima CF, Teodoro AJ. Antioxidant Capacity, Antitumor Activity and Metabolomic Profile of a Beetroot Peel Flour. Metabolites. 2023; 13(2):277. https://doi.org/10.3390/metabo13020277
Chicago/Turabian StyleCoimbra, Pedro Paulo Saldanha, Anna Carolina Alves Gomes da Silva-e-Silva, Ananda da Silva Antonio, Henrique Marcelo Gualberto Pereira, Valdir Florêncio da Veiga-Junior, Israel Felzenszwalb, Carlos Fernando Araujo-Lima, and Anderson Junger Teodoro. 2023. "Antioxidant Capacity, Antitumor Activity and Metabolomic Profile of a Beetroot Peel Flour" Metabolites 13, no. 2: 277. https://doi.org/10.3390/metabo13020277
APA StyleCoimbra, P. P. S., Silva-e-Silva, A. C. A. G. d., Antonio, A. d. S., Pereira, H. M. G., Veiga-Junior, V. F. d., Felzenszwalb, I., Araujo-Lima, C. F., & Teodoro, A. J. (2023). Antioxidant Capacity, Antitumor Activity and Metabolomic Profile of a Beetroot Peel Flour. Metabolites, 13(2), 277. https://doi.org/10.3390/metabo13020277