
Mass-Spectrometry-Based Lipidomics Discriminates Specific Changes in Lipid Classes in Healthy and Dyslipidemic Adults

 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Experimental Design
2.1. Study Design, Participants and Sampling
2.2. Mass Spectrometry Analysis
2.3. Data Analysis and Feature Selection
3. Results
3.1. MS-Annotated Long-Chain TAG and Cer, Are Positively Associated with TAG Serum Levels
3.2. MS-Annotated PC Associated with Serum HDL-C Levels
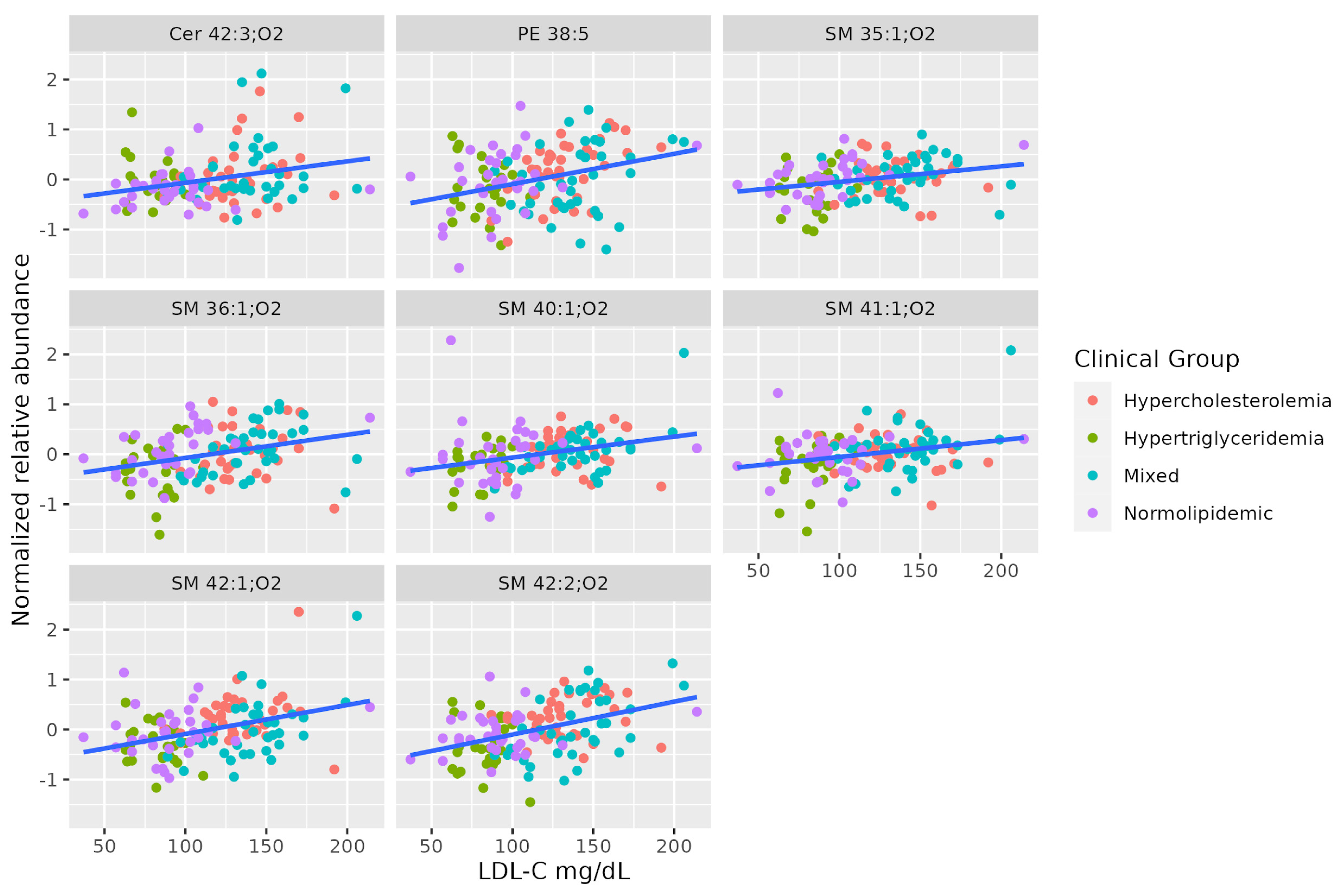
3.3. MS-Annotated SM Associated with Serum LDL-C Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Noncommunicable Diseases: Risk Factors. 2021. Available online: https://www.who.int/data/gho/data/themes/topics/noncommunicable-diseases-risk-factors. (accessed on 1 December 2022).
- Berberich, A.J.; Hegele, R.A. A Modern Approach to Dyslipidemia. Endocr. Rev. 2022, 43, 4611–4653. [Google Scholar] [CrossRef] [PubMed]
- Musunuru, K. Atherogenic Dyslipidemia: Cardiovascular Risk and Dietary Intervention. Lipids 2010, 45, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Sokooti, S.; Flores-Guerrero, J.L.; Heerspink, H.J.L.; Connelly, M.A.; Bakker, S.J.L.; Dullaart, R.P.F. Triglyceride-rich lipoprotein and LDL particle subfractions and their association with incident type 2 diabetes: The PREVEND study. Cardiovasc. Diabetol. 2021, 20, 156. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Ye, J.; Shao, C.; Li, F.; Lin, Y.; Ma, Q.; Wang, W.; Feng, S.; Zhong, B. Varied Relationship of Lipid and Lipoprotein Profiles to Liver Fat Content in Phenotypes of Metabolic Associated Fatty Liver Disease. Front. Endocrinol. 2021, 12, 691556. [Google Scholar] [CrossRef] [PubMed]
- Berglund, L.; Brunzell, J.D.; Goldberg, A.C.; Goldberg, I.J.; Sacks, F.; Murad, M.H.; Stalenhoef, A.F. Evaluation and Treatment of Hypertriglyceridemia: An Endocrine Society Clinical Practice Guideline. J. Clin. Endocrinol. Metab. 2012, 97, 2969–2989. [Google Scholar] [CrossRef]
- Hegele, R.A. Plasma lipoproteins: Genetic influences and clinical implications. Nat. Rev. Genet. 2009, 10, 109–121. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Ginsberg, H.N.; Packard, C.J.; Chapman, M.J.; Borén, J.; Aguilar-Salinas, C.A.; Averna, M.; Ference, B.A.; Gaudet, D.; Hegele, R.A.; Kersten, S.; et al. Triglyceride-rich lipoproteins and their remnants: Metabolic insights, role in atherosclerotic cardiovascular disease, and emerging therapeutic strategies—A consensus statement from the European Atherosclerosis Society. Eur. Heart J. 2021, 42, 4791–4806. [Google Scholar] [CrossRef]
- Aciksari, G.; Cetinkal, G.; Kocak, M.; Atici, A.; Celik, F.B.; Caliskan, M. The relationship between triglyceride/high-density lipoprotein cholesterol ratio and coronary slow-flow phenomenon. Int. J. Cardiovasc. Imaging 2022, 38, 5–13. [Google Scholar] [CrossRef]
- Podrez, E.A.; Febbraio, M.; Sheibani, N.; Schmitt, D.; Silverstein, R.L.; Hajjar, D.P.; Cohen, P.A.; Frazier, W.A.; Hoff, H.F.; Hazen, S.L. Macrophage scavenger receptor CD36 is the major receptor for LDL modified by monocyte-generated reactive nitrogen species. J. Clin. Investig 2000, 105, 1095–1108. [Google Scholar] [CrossRef]
- Wolska, A.; Yang, Z.-H.; Remaley, A.T. Hypertriglyceridemia: New approaches in management and treatment. Curr. Opin. Lipidol. 2020, 31, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.K. Emerging Risk Biomarkers in Cardiovascular Diseases and Disorders. J. Lipids 2015, 2015, 971453. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cong, H.; Zhang, J.; Hu, Y.; Wei, A.; Zhang, Y.; Yang, H.; Ren, L.; Qi, W.; Li, W. Predictive Value of the Triglyceride to High-Density Lipoprotein Cholesterol Ratio for All-Cause Mortality and Cardiovascular Death in Diabetic Patients with Coronary Artery Disease Treated With Statins. Front. Cardiovasc. Med. 2021, 8, 718604. [Google Scholar] [CrossRef] [PubMed]
- Voight, B.F.; Peloso, G.M.; Orho-Melander, M.; Frikke-Schmidt, R.; Barbalic, M.; Jensen, M.K.; Hindy, G.; Hólm, H.; Ding, E.L.; Johnson, T.; et al. Plasma HDL cholesterol and risk of myocardial infarction: A mendelian randomisation study. Lancet 2012, 380, 572–580. [Google Scholar] [CrossRef]
- Ding, M.; Rexrode, K.M. A Review of Lipidomics of Cardiovascular Disease Highlights the Importance of Isolating Lipoproteins. Metabolites 2020, 10, 163. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Faludi, A.; Izar, M.; Saraiva, J.; Chacra, A.; Bianco, H.; Afiune, N.; Bertolami, A.; Pereira, A.C.; Lottenberg, A.M.; Sposito, A.C.; et al. Atualização da Diretriz Brasileira de Dislipidemias e Prevenção da Aterosclerose. Arq. Bras. Cardiol. 2017, 109, 1–76. [Google Scholar] [CrossRef]
- Mecatti, G.C.; Sánchez-Vinces, S.; Fernandes, A.M.A.P.; Messias, M.C.F.; de Santis, G.K.D.; Porcari, A.M.; Marson, F.A.; Carvalho, P.D.O. Potential Lipid Signatures for Diagnosis and Prognosis of Sepsis and Systemic Inflammatory Response Syndrome. Metabolites 2020, 10, 359. [Google Scholar] [CrossRef]
- Fernandes, A.M.A.P.; Messias, M.C.F.; Duarte, G.H.B.; de Santis, G.K.D.; Mecatti, G.C.; Porcari, A.M.; Murgu, M.; Simionato, A.V.C.; Rocha, T.; Martinez, C.A.; et al. Plasma Lipid Profile Reveals Plasmalogens as Potential Biomarkers for Colon Cancer Screening. Metabolites 2020, 10, 262. [Google Scholar] [CrossRef]
- Fan, S.; Kind, T.; Cajka, T.; Hazen, S.L.; Tang, W.H.W.; Kaddurah-Daouk, R.; Irvin, M.R.; Arnett, D.K.; Barupal, D.K.; Fiehn, O. Systematic Error Removal Using Random Forest for Normalizing Large-Scale Untargeted Lipidomics Data. Anal. Chem. 2019, 91, 3590–3596. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Li, S.; Xia, J. MetaboAnalystR 3.0: Toward an Optimized Workflow for Global Metabolomics. Metabolites 2020, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Sud, M.; Fahy, E.; Cotter, D.; Brown, A.; Dennis, E.A.; Glass, C.K.; Merrill, A.H.J.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; et al. LMSD: LIPID MAPS structure database. Nucleic Acids Res. 2007, 35, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Sah, S.; Ma, X.; Botros, A.; Gaul, D.A.; Yun, S.R.; Park, E.Y.; Kim, O.; Moore, S.G.; Kim, J.; Fernández, F.M. Space- and Time-Resolved Metabolomics of a High-Grade Serous Ovarian Cancer Mouse Model. Cancers 2022, 14, 2262. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Köfeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Invest. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Sanders, F.; McNally, B.; Griffin, J.L. Blood triacylglycerols: A lipidomic window on diet and disease. Biochem. Soc. Trans. 2016, 44, 638–644. [Google Scholar] [CrossRef]
- Meikle, P.J.; Summers, S.A. Sphingolipids and phospholipids in insulin resistance and related metabolic disorders. Nat. Rev. Endocrinol. 2017, 13, 79–91. [Google Scholar] [CrossRef]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Hussain, M.M. Sphingolipids and Lipoproteins in Health and Metabolic Disorders. Trends Endocrinol. Metab. 2017, 28, 506–518. [Google Scholar] [CrossRef]
- Yun, H.; Sun, L.; Wu, Q.; Zong, G.; Qi, Q.; Li, H.; Zheng, H.; Zeng, R.; Liang, L.; Lin, X. Associations among circulating sphingolipids, β-cell function, and risk of developing type 2 diabetes: A population-based cohort study in China. PLoS Med. 2020, 17, e1003451. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Salonurmi, T.; Havulinna, A.S.; Kauhanen, D.; Pedersen, E.R.; Tell, G.S.; Meyer, K.; Teeriniemi, A.M.; Laatikainen, T.; Jousilahti, P.; et al. Ceramide stearic to palmitic acid ratio predicts incident diabetes. Diabetologia 2018, 61, 1424–1434. [Google Scholar] [CrossRef] [PubMed]
- Lachkar, F.; Ferré, P.; Foufelle, F.; Papaioannou, A. Dihydroceramides: Their emerging physiological roles and functions in cancer and metabolic diseases. Am. J. Physiol. Endocrinol. Metab. 2020, 320, 122–130. [Google Scholar] [CrossRef]
- Hammad, S.M.; Harden, O.C.; Wilson, D.A.; Twal, W.O.; Nietert, P.J.; Oates, J.C. Plasma Sphingolipid Profile Associated with Subclinical Atherosclerosis and Clinical Disease Markers of Systemic Lupus Erythematosus: Potential Predictive Value. Front. Immunol. 2021, 12, 694318. [Google Scholar] [CrossRef]
- Ishigami, M.; Ogasawara, F.; Nagao, K.; Hashimoto, H.; Kimura, Y.; Kioka, N.; Ueda, K. Temporary sequestration of cholesterol and phosphatidylcholine within extracellular domains of ABCA1 during nascent HDL generation. Sci. Rep. 2018, 8, 6170. [Google Scholar] [CrossRef] [PubMed]
- Storlien, L.H.; Jenkins, A.B.; Chisholm, D.J.; Pascoe, W.S.; Khouri, S.; Kraegen, E.W. Influence of Dietary Fat Composition on Development of Insulin Resistance in Rats: Relationship to Muscle Triglyceride and ω-3 Fatty Acids in Muscle Phospholipid. Diabetes 1991, 40, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Funai, K.; Lodhi, I.J.; Spears, L.D.; Yin, L.; Song, H.; Klein, S.; Semenkovich, C.F. Skeletal Muscle Phospholipid Metabolism Regulates Insulin Sensitivity and Contractile Function. Diabetes 2016, 65, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 2007, 46, 1081–1090. [Google Scholar] [CrossRef]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab. 2006, 3, 321–331. [Google Scholar] [CrossRef]
- Wang, X.; Lam, S.M.; Cao, M.; Wang, T.; Wang, Z.; Yu, M.; Li, B.; Zhang, H.; Ping, F.; Song, G.; et al. Localized increases in CEPT1 and ATGL elevate plasmalogen phosphatidylcholines in HDLs contributing to atheroprotective lipid profiles in hyperglycemic GCK-MODY. Redox. Biol. 2021, 40, 101855. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | TAG (mg dL−1) | TC (mg dL−1) | HDL-C (mg dL−1) | LDL-C (mg dL−1) | |
|---|---|---|---|---|---|
| Normolipidemic | 32 | 96.68 (28.76) | 190.73 (28.6) | 63.92 (13.67) | 107.32 (26.56) |
| Isolated hypertriglyceridemia | 23 | 211.2 (58.58) | 205.15 (30.37) | 52.87 (10.83) | 111.24 (30.26) |
| Isolated hypercholesterolemia | 36 | 112.33 (25.67) | 223.5 (59.74) | 57.33 (17.18) | 178.33 (20.75) |
| Mixed dyslipidemia | 37 | 213.4 (60.05) | 274.8 (25.45) | 51.6 (10.83) | 183.4 (17.84) |
| rt_m/z | Coefficient TAG | Adj.P. Value | Annotation | Mass Error (ppm) | Matched Fragments | Annotation Confidence Level * |
|---|---|---|---|---|---|---|
| 2.82_720.5891 m/z | −0.01 | 0.00 | PC 32:0 | −1.43 | 537.5244 a | 2 |
| 0.70_495.3337 n | 0.01 | 0.02 | PC 16:0 | 2.49 | 184.0744 a | 2 |
| 2.33_747.5641 m/z | −0.02 | 0.00 | SM 34:1;O2 | −2.35 | 687.5428 a | 2 |
| 2.33_766.5372 m/z | 0.01 | 0.02 | PE 38:4 | −2.59 | 303.2329 b | 2 |
| 2.33_826.5580 m/z | 0.01 | 0.01 | PC 36:4 | −3.02 | 766.5370 a | 2 |
| 2.55_835.5318 m/z | 0.01 | 0.03 | PI 34:1 | −2.93 | 581.3099/241.0119 a | 2 |
| 2.56_885.5479 m/z | 0.02 | 0.00 | PI 38:4 | −2.19 | 581.3092/241.0119 a | 2 |
| 2.57_734.5702 m/z | −0.01 | 0.01 | PEth 35:1 | 1.03 | 383.3521 a | 2 |
| 2.67_801.6098 m/z | −0.01 | 0.01 | SM 38:2;O2 | −3.93 | 741.5892 a | 2 |
| 2.74_766.5376 m/z | 0.02 | 0.00 | PE 38:4 | −2.13 | 303.2329 b | 2 |
| 2.78_774.5420 m/z | −0.01 | 0.02 | PE 40:7 | −2.99 | 305.2482 b | 2 |
| 3.47_638.5728 m/z | 0.01 | 0.01 | Cer 38:1;O2 | −0.14 | 592.5703 a | 2 |
| 3.50_664.5877 m/z | 0.01 | 0.03 | Cer 40:2;O2 | −1.43 | 618.5831 a | 2 |
| 3.76_678.6035 m/z | 0.01 | 0.02 | Cer 41:2;O2 | −1.16 | 632.6000 a | 2 |
| 3.95_692.6179 m/z | 0.01 | 0.02 | Cer 42:2;O2 | −2.99 | 646.6137 a | 2 |
| 4.01_666.6029 m/z | 0.01 | 0.01 | Cer 40:1;O2 | −2.03 | 620.5986 a | 2 |
| 5.37_774.6727 n | 0.01 | 0.00 | TAG 46:2 | −1.33 | 521.4569/519.4422 a | 2 |
| 5.58_776.6897 n | 0.01 | 0.00 | TAG 46:1 | 0.34 | 523.4730/521.4577 a | 2 |
| 5.59_802.7060 n | 0.01 | 0.00 | TAG 48:2 | 1.15 | 549.4890/547.4738 a | 2 |
| 5.79_830.7374 n | 0.01 | 0.00 | TAG 50:2 | 1.23 | 577.5199/549.4895 a | 2 |
| 6.02_858.7692 n | 0.01 | 0.00 | TAG 52:2 | 1.77 | 603.5365/577.5206 a | 2 |
| 6.25_886.7993 n | 0.01 | 0.00 | TAG 54:2 | 0.44 | 631.5711/605.5506 a | 2 |
| 6.26_860.7828 n | 0.01 | 0.04 | TAG 52:1 | −0.59 | 605.5506/579.5338 a | 2 |
| 6.27_834.7655 n | 0.01 | 0.01 | TAG 50:0 | −2.59 | 579.5338 a | 2 |
| Rt_m/z | Coefficient HDL-C | Adj.P.Value | Annotation | Mass Error (ppm) | Match Fragments | Annotation Confidence Level * |
|---|---|---|---|---|---|---|
| 2.04_848.5460 m/z | 0.06 | 0.00 | PC 38:7 | 1.55 | 788.5238/327.2329 b | 2 |
| 2.52_786.5633 m/z | 0.09 | 0.00 | PC 34:3 | −2.89 | 726.5423 a | 2 |
| 2.84_866.6248 m/z | 0.05 | 0.02 | PC 40:5 | −3.99 | 806.6031 a | 2 |
| 2.86_790.5978 m/z | 0.05 | 0.02 | PC 34:1 | 1.37 | 730.5785 a | 2 |
| rt_m/z | Coefficient LDL-C | Adj.P.Value | Annotation | Mass Error (ppm) | Match Fragments | Annotation Confidence Level * |
|---|---|---|---|---|---|---|
| 2.48_761.5795 m/z | 0.02 | 0.04 | SM 35:1;O2 | −2.65 | 701.5586 a | 2 |
| 2.65_775.5947 m/z | 0.03 | 0.01 | SM 36:1;O2 | −3.23 | 715.5747 a | 2 |
| 2.91_750.5440 m/z | 0.03 | 0.03 | PE 38:5 | −0.38 | 464.3146/303.2329 b | 2 |
| 3.43_857.6733 m/z | 0.04 | 0.00 | SM 42:2;O2 | −2.47 | 797.6521 a | 2 |
| 3.47_690.6029 m/z | 0.02 | 0.04 | Cer 42:3;O2 | −2.08 | 644.5987 a | 2 |
| 3.47_831.6576 m/z | 0.02 | 0.03 | SM 40:1;O2 | −2.62 | 771.6366 a | 2 |
| 3.75_845.6731 m/z | 0.02 | 0.03 | SM 41:1;O2 | −2.82 | 785.6521 a | 2 |
| 4.03_859.6888 m/z | 0.03 | 0.02 | SM 42:1;O2 | −2.72 | 799.6677 a | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Vinces, S.; Garcia, P.H.D.; Silva, A.A.R.; Fernandes, A.M.A.d.P.; Barreto, J.A.; Duarte, G.H.B.; Antonio, M.A.; Birbrair, A.; Porcari, A.M.; Carvalho, P.d.O. Mass-Spectrometry-Based Lipidomics Discriminates Specific Changes in Lipid Classes in Healthy and Dyslipidemic Adults. Metabolites 2023, 13, 222. https://doi.org/10.3390/metabo13020222
Sánchez-Vinces S, Garcia PHD, Silva AAR, Fernandes AMAdP, Barreto JA, Duarte GHB, Antonio MA, Birbrair A, Porcari AM, Carvalho PdO. Mass-Spectrometry-Based Lipidomics Discriminates Specific Changes in Lipid Classes in Healthy and Dyslipidemic Adults. Metabolites. 2023; 13(2):222. https://doi.org/10.3390/metabo13020222
Chicago/Turabian StyleSánchez-Vinces, Salvador, Pedro Henrique Dias Garcia, Alex Ap. Rosini Silva, Anna Maria Alves de Piloto Fernandes, Joyce Aparecida Barreto, Gustavo Henrique Bueno Duarte, Marcia Aparecida Antonio, Alexander Birbrair, Andreia M. Porcari, and Patricia de Oliveira Carvalho. 2023. "Mass-Spectrometry-Based Lipidomics Discriminates Specific Changes in Lipid Classes in Healthy and Dyslipidemic Adults" Metabolites 13, no. 2: 222. https://doi.org/10.3390/metabo13020222
APA StyleSánchez-Vinces, S., Garcia, P. H. D., Silva, A. A. R., Fernandes, A. M. A. d. P., Barreto, J. A., Duarte, G. H. B., Antonio, M. A., Birbrair, A., Porcari, A. M., & Carvalho, P. d. O. (2023). Mass-Spectrometry-Based Lipidomics Discriminates Specific Changes in Lipid Classes in Healthy and Dyslipidemic Adults. Metabolites, 13(2), 222. https://doi.org/10.3390/metabo13020222