
Impact of Cesarean Delivery and Breastfeeding on Secretory Immunoglobulin A in the Infant Gut Is Mediated by Gut Microbiota and Metabolites

 , ,
, ,  , and
, and
Abstract
1. Introduction
2. Methods
2.1. Study Design
Infant Fecal Microbiota, Metabolites and SIgA
2.2. Statistical Analysis
2.3. Covariates
3. Results
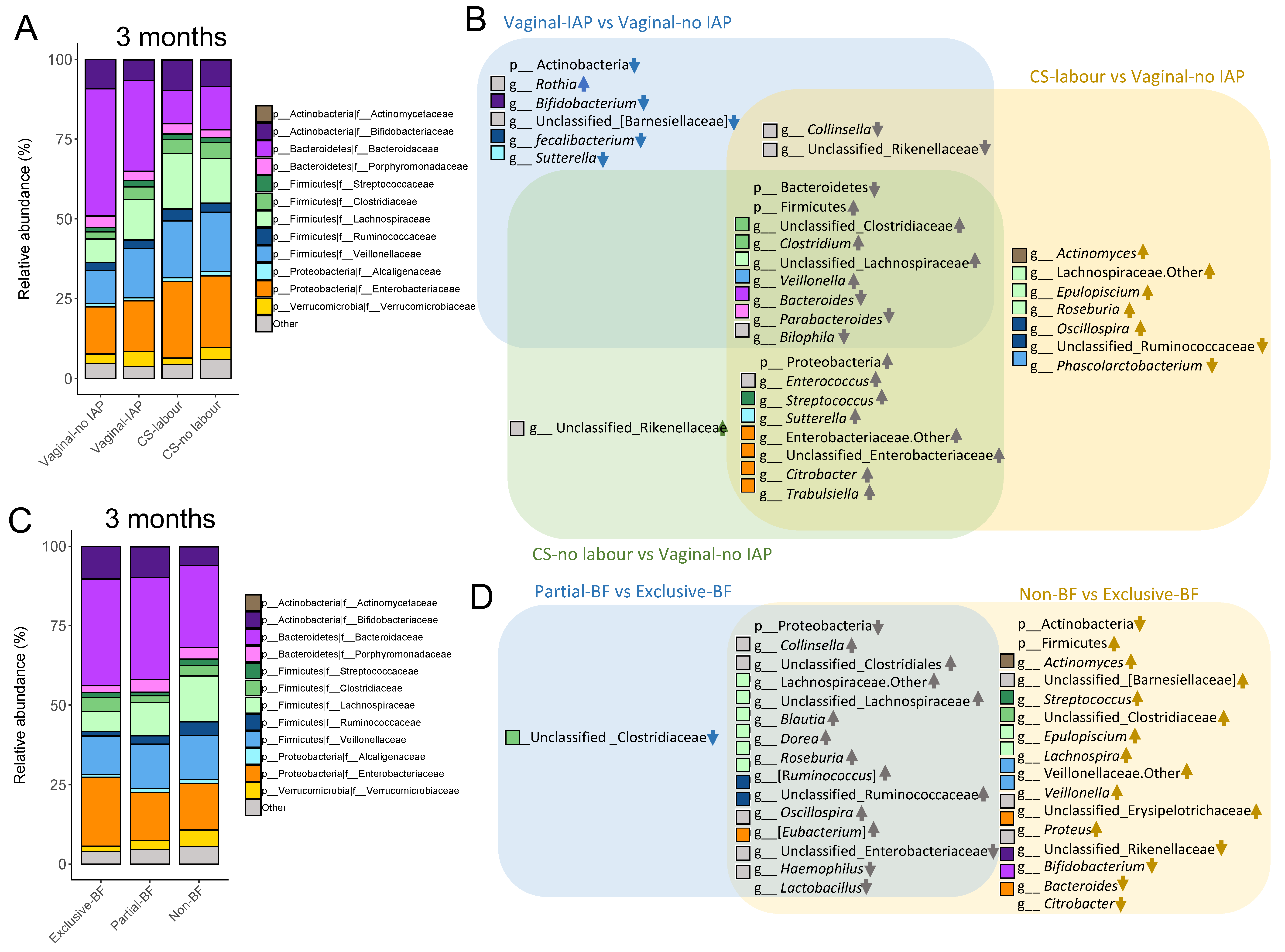
3.1. Birth Mode, SIgA Levels and Gut Microbiota of Infants at 3 and 12 Months
3.2. SIgA Correlations with Gut Microbial Taxa and Metabolites
3.3. Mediating Microbe—Metabolite Pathways from Birth Mode to SIgA Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peterson, D.A.; McNulty, N.P.; Guruge, J.L.; Gordon, J.I. IgA Response to Symbiotic Bacteria as a Mediator of Gut Homeostasis. Cell Host Microbe 2007, 2, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Rol, N.; Corthésy, B. Secretory IgA’s complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Battersby, A.J.; Gibbons, D.L. The gut mucosal immune system in the neonatal period. Pediatr. Allergy Immunol. 2013, 24, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Kawano, A.; Emori, Y. Changes in maternal secretory immunoglobulin a levels in human milk during 12 weeks after parturition. Am. J. Hum. Biol. 2013, 25, 399–403. [Google Scholar] [CrossRef]
- Rogier, E.W.; Frantz, A.L.; Bruno, M.E.C.; Wedlund, L.; Cohen, D.A.; Stromberg, A.J.; Kaetzel, C.S. Secretory antibodies in breast milk promote long-term intestinal homeostasis by regulating the gut microbiota and host gene expression. Proc. Natl. Acad. Sci. USA 2014, 111, 3074–3079. [Google Scholar] [CrossRef]
- Lamberti, L.M.; Walker, C.L.F.; Noiman, A.; Victora, C.; Black, R.E. Breastfeeding and the risk for diarrhea morbidity and mortality. BMC Public Health 2011, 11 (Suppl. 3), S15. [Google Scholar] [CrossRef] [PubMed]
- Cervia, C.; Nilsson, J.; Zurbuchen, Y.; Valaperti, A.; Schreiner, J.; Wolfensberger, A.; Raeber, M.E.; Adamo, S.; Weigang, S.; Emmenegger, M.; et al. Systemic and mucosal antibody responses specific to SARS-CoV-2 during mild versus severe COVID-19. J. Allergy Clin. Immunol. 2021, 147, 545–557. [Google Scholar] [CrossRef]
- Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Viant, C.; Gaebler, C.; Cipolla, M.; Hoffmann, H.-H.; Oliveira, T.Y.; Oren, D.A.; et al. Enhanced SARS-CoV-2 neutralization by dimeric IgA. Sci. Transl. Med. 2021, 13, eabf1555. [Google Scholar] [CrossRef]
- Goenka, A.; Halliday, A.; Gregorova, M.; Milodowski, E.; Thomas, A.; Williamson, M.K.; Baum, H.; Oliver, E.; Long, A.E.; Knezevic, L.; et al. Young infants exhibit robust functional antibody responses and restrained IFN-γ production to SARS-CoV-2. Cell Rep. Med. 2021, 2, 100327. [Google Scholar] [CrossRef]
- Conti, M.G.; Terreri, S.; Mortari, E.P.; Albano, C.; Natale, F.; Boscarino, G.; Zacco, G.; Palomba, P.; Cascioli, S.; Corrente, F.; et al. Immune Response of Neonates Born to Mothers Infected With SARS-CoV-2. JAMA Netw. Open 2021, 4, e2132563. [Google Scholar] [CrossRef]
- Boix-Amoros, A.; Collado, M.C.; Van’t Land, B.; Calvert, A.; Le Doare, K.; Garssen, J.; Hanna, H.; Khaleva, E.; Peroni, D.G.; Geddes, D.T.; et al. Reviewing the evidence on breast milk composition and immunological outcomes. Nutr. Rev. 2019, 77, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef] [PubMed]
- Sanidad, K.Z.; Zeng, M.Y. Neonatal gut microbiome and immunity. Curr. Opin. Microbiol. 2020, 56, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.-H.; Kim, M.; Yun, C.-H. Regulation of Gastrointestinal Immunity by Metabolites. Nutrients 2021, 13, 167. [Google Scholar] [CrossRef]
- Brandtzaeg, P. The Mucosal Immune System and Its Integration with the Mammary Glands. J. Pediatr. 2010, 156, S8–S15. [Google Scholar] [CrossRef]
- Bunker, J.J.; Flynn, T.M.; Koval, J.C.; Shaw, D.G.; Meisel, M.; McDonald, B.D.; Ishizuka, I.E.; Dent, A.L.; Wilson, P.C.; Jabri, B.; et al. Innate and Adaptive Humoral Responses Coat Distinct Commensal Bacteria with Immunoglobulin A. Immunity 2015, 43, 541–553. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Gatto, D.; Sainsbury, E.; Harriman, G.R.; Hengartner, H.; Zinkernagel, R.M. A Primitive T Cell-Independent Mechanism of Intestinal Mucosal IgA Responses to Commensal Bacteria. Science 2000, 288, 2222–2226. [Google Scholar] [CrossRef]
- Lin, L.; Zhang, J. Role of intestinal microbiota and metabolites on gut homeostasis and human diseases. BMC Immunol. 2017, 18, 2. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Rutayisire, E.; Huang, K.; Liu, Y.; Tao, F. The mode of delivery affects the diversity and colonization pattern of the gut microbiota during the first year of infants’ life: A systematic review. BMC Gastroenterol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Azad, M.; Konya, T.; Persaud, R.; Guttman, D.; Chari, R.; Field, C.; Sears, M.R.; Mandhane, P.; Turvey, S.; Subbarao, P.; et al. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: A prospective cohort study. BJOG Int. J. Obstet. Gynaecol. 2015, 123, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Reyman, M.; Van Houten, M.A.; Van Baarle, D.; Bosch, A.A.T.M.; Man, W.H.; Chu, M.L.J.N.; Arp, K.; Watson, R.L.; Sanders, E.A.M.; Fuentes, S.; et al. Impact of delivery mode-associated gut microbiota dynamics on health in the first year of life. Nat. Commun. 2019, 10, 4997. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Zhao, X.; Moeder, W.; Tun, H.M.; Simons, E.; Mandhane, P.J.; Moraes, T.J.; Turvey, S.E.; Subbarao, P.; Scott, J.A.; et al. Impact of Maternal Intrapartum Antibiotics, and Caesarean Section with and without Labour on Bifidobacterium and Other Infant Gut Microbiota. Microorganisms 2021, 9, 1847. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, H.; Kuguoglu, S. Comparison of the breastfeeding patterns of mothers who delivered their babies per vagina and via cesarean section: An observational study using the LATCH breastfeeding charting system. Int. J. Nurs. Stud. 2007, 44, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Chari, R.S.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L. Gut microbiota of healthy Canadian infants: Profiles by mode of delivery and infant diet at 4 months. Can. Med. Assoc. J. 2013, 185, 385–394. [Google Scholar] [CrossRef]
- Drall, K.M.; Tun, H.M.; Morales-Lizcano, N.P.; Konya, T.B.; Guttman, D.S.; Field, C.; Mandal, R.; Wishart, D.S.; Becker, A.B.; Azad, M.B.; et al. Clostridioides difficile Colonization Is Differentially Associated With Gut Microbiome Profiles by Infant Feeding Modality at 3–4 Months of Age. Front. Immunol. 2019, 10, 2866. [Google Scholar] [CrossRef] [PubMed]
- Vu, K.; Lou, W.; Tun, H.M.; Konya, T.B.; Morales-Lizcano, N.; Chari, R.S.; Field, C.J.; Guttman, D.S.; Mandal, R.; Wishart, D.S.; et al. From Birth to Overweight and Atopic Disease: Multiple and Common Pathways of the Infant Gut Microbiome. Gastroenterology 2021, 160, 128–144. [Google Scholar] [CrossRef] [PubMed]
- Mirpuri, J.; Raetz, M.; Sturge, C.R.; Wilhelm, C.L.; Benson, A.; Savani, R.C.; Hooper, L.V.; Yarovinsky, F. Proteobacteria-specific IgA regulates maturation of the intestinal microbiota. Gut Microbes 2013, 5, 28–39. [Google Scholar] [CrossRef]
- Morita, N.; Umemoto, E.; Fujita, S.; Hayashi, A.; Kikuta, J.; Kimura, I.; Haneda, T.; Imai, T.; Inoue, A.; Mimuro, H.; et al. GPR31-dependent dendrite protrusion of intestinal CX3CR1+ cells by bacterial metabolites. Nature 2019, 566, 110–114. [Google Scholar] [CrossRef]
- Azad, M.B.; Konya, T.; Guttman, D.S.; Field, C.J.; Sears, M.R.; HayGlass, K.T.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Becker, A.B.; et al. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin. Exp. Allergy 2015, 45, 632–643. [Google Scholar] [CrossRef]
- Bridgman, S.L.; Azad, M.B.; Field, C.J.; Haqq, A.M.; Becker, A.B.; Mandhane, P.J.; Subbarao, P.; Turvey, S.E.; Sears, M.R.; Scott, J.A.; et al. Fecal Short-Chain Fatty Acid Variations by Breastfeeding Status in Infants at 4 Months: Differences in Relative versus Absolute Concentrations. Front. Nutr. 2017, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.J.; Koleva, P.T.; Field, C.J.; Giesbrecht, G.F.; Wine, E.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Sears, M.R.; et al. Maternal depressive symptoms linked to reduced fecal Immunoglobulin A concentrations in infants. Brain Behav. Immun. 2018, 68, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Osborne, J. Improving your data transformations: Applying the Box-Cox transformation. Pract. Assess. Res. Eval. 2010, 15, 12. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Ogle, D.H. Introductory Fisheries Analyses with R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2018. [Google Scholar]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach; Guilford Publications: New York, NY, USA, 2017. [Google Scholar]
- Bridgman, S.L.; Konya, T.; Azad, M.B.; Sears, M.R.; Becker, A.B.; Turvey, S.E.; Mandhane, P.J.; Subbarao, P.; Scott, J.A.; Field, C.J.; et al. Infant gut immunity: A preliminary study of IgA associations with breastfeeding. J. Dev. Orig. Health Dis. 2015, 7, 68–72. [Google Scholar] [CrossRef]
- Czosnykowska-Łukacka, M.; Lis-Kuberka, J.; Królak-Olejnik, B.; Orczyk-Pawiłowicz, M. Changes in Human Milk Immunoglobulin Profile During Prolonged Lactation. Front. Pediatr. 2020, 8, 428. [Google Scholar] [CrossRef]
- Shrier, I.; Platt, R.W. Reducing bias through directed acyclic graphs. BMC Med. Res. Methodol. 2008, 8, 70. [Google Scholar] [CrossRef]
- Evans, D.; Chaix, B.; Lobbedez, T.; Verger, C.; Flahault, A. Combining directed acyclic graphs and the change-in-estimate procedure as a novel approach to adjustment-variable selection in epidemiology. BMC Med. Res. Methodol. 2012, 12, 156. [Google Scholar] [CrossRef]
- Huus, K.E.; Petersen, C.; Finlay, B.B. Diversity and dynamism of IgA−microbiota interactions. Nat. Rev. Immunol. 2021, 21, 514–525. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Mazzoni, C.; Hogstrom, L.; Bryant, A.; Bergerat, A.; Cher, A.; Pochan, S.; Herman, P.; Carrigan, M.; Sharp, K.; et al. Delivery Mode Affects Stability of Early Infant Gut Microbiota. Cell Rep. Med. 2020, 1, 100156. [Google Scholar] [CrossRef]
- Lawson, M.A.E.; O’Neill, I.J.; Kujawska, M.; Javvadi, S.G.; Wijeyesekera, A.; Flegg, Z.; Chalklen, L.; Hall, L.J. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2019, 14, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.K.; Wall, R. Selective removal of alpha heavy-chain glycosylation sites causes immunoglobulin A degradation and reduced secretion. Mol. Cell. Biol. 1988, 8, 4197–4203. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Gay, M.C.L.; Koleva, P.T.; Slupsky, C.M.; Du Toit, E.; Eggesbo, M.; Johnson, C.C.; Wegienka, G.; Shimojo, N.; Campbell, D.E.; Prescott, S.L.; et al. Worldwide Variation in Human Milk Metabolome: Indicators of Breast Physiology and Maternal Lifestyle? Nutrients 2018, 10, 1151. [Google Scholar] [CrossRef] [PubMed]
- Limon, A.; Gallegos-Perez, J.-L.; Reyes-Ruiz, J.M.; Aljohi, M.A.; Alshanqeeti, A.S.; Miledi, R. The endogenous GABA bioactivity of camel, bovine, goat and human milks. Food Chem. 2014, 145, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and immunity enhancement effects of γ-Aminobutyric acid (GABA) administration in humans. Biofactors 2006, 26, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Kubota, A.; Kobayashi, M.; Sarashina, S.; Takeno, R.; Yasuda, G.; Narumi, K.; Furugen, A.; Takahashi-Suzuki, N.; Iseki, K. Gamma-Aminobutyric Acid (GABA) Attenuates Ischemia Reperfusion-Induced Alterations in Intestinal Immunity. Biol. Pharm. Bull. 2018, 41, 1874–1878. [Google Scholar] [CrossRef]
- Goonatilleke, E.; Smilowitz, J.T.; Mariño, K.V.; German, B.J.; Lebrilla, C.B.; Barboza, M. Immunoglobulin A N-glycosylation Presents Important Body Fluid-specific Variations in Lactating Mothers. Mol. Cell. Proteom. 2019, 18, 2165–2177. [Google Scholar] [CrossRef]
- Pham, V.T.; Chassard, C.; Rifa, E.; Braegger, C.; Geirnaert, A.; Martin, V.N.R.; Lacroix, C. Lactate Metabolism Is Strongly Modulated by Fecal Inoculum, pH, and Retention Time in PolyFermS Continuous Colonic Fermentation Models Mimicking Young Infant Proximal Colon. Msystems 2019, 4, e00264-18. [Google Scholar] [CrossRef]
- Holscher, H.D.; Czerkies, L.A.; Cekola, P.; Litov, R.; Benbow, M.; Santema, S.; Alexander, D.D.; Perez, V.; Sun, S.; Saavedra, J.M.; et al. Bifidobacterium lactis Bb12 Enhances Intestinal Antibody Response in Formula-Fed Infants. J. Parenter. Enter. Nutr. 2012, 36, 106S–117S. [Google Scholar] [CrossRef]
- Lam, W.Y.; Becker, A.M.; Kennerly, K.M.; Wong, R.; Curtis, J.D.; Llufrio, E.M.; McCommis, K.S.; Fahrmann, J.; Pizzato, H.A.; Nunley, R.M.; et al. Mitochondrial Pyruvate Import Promotes Long-Term Survival of Antibody-Secreting Plasma Cells. Immunity 2016, 45, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Wilmore, J.R.; Gaudette, B.T.; Atria, D.G.; Hashemi, T.; Jones, D.D.; Gardner, C.A.; Cole, S.D.; Misic, A.M.; Beiting, D.P.; Allman, D. Commensal Microbes Induce Serum IgA Responses that Protect against Polymicrobial Sepsis. Cell Host Microbe 2018, 23, 302–311. [Google Scholar] [CrossRef]
- Neumann-Schaal, M.; Jahn, D.; Schmidt-Hohagen, K. Metabolism the Difficile Way: The Key to the Success of the Pathogen Clostridioides difficile. Front. Microbiol. 2019, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Ichimura, M.; Tsuneyama, K.; Moritoki, Y.; Tsunashima, H.; Omagari, K.; Hara, M.; Yasuda, I.; Miyakawa, H.; Kikuchi, K. Fructo-oligosaccharides and intestinal barrier function in a methionine–choline-deficient mouse model of nonalcoholic steatohepatitis. PLoS ONE 2017, 12, e0175406. [Google Scholar] [CrossRef] [PubMed]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary Choline Intake: Current State of Knowledge Across the Life Cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, Å.; Duan, R.-D. Pancreatic and mucosal enzymes in choline phospholipid digestion. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G425–G445. [Google Scholar] [CrossRef]
- Olson, A.; Diebel, L.N.; Liberati, D.M. Effect of host defenses on Clostridium difficile toxin–induced intestinal barrier injury. J. Trauma Inj. Infect. Crit. Care 2013, 74, 983–990. [Google Scholar] [CrossRef]
- Fagone, P.; Gunter, C.; Sage, C.R.; Gunn, K.E.; Brewer, J.W.; Jackowski, S. CTP:Phosphocholine Cytidylyltransferase α Is Required for B-cell Proliferation and Class Switch Recombination. J. Biol. Chem. 2009, 284, 6847–6854. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Q.; Luo, K.; He, P.; Hou, R.; Zhao, X.; Wang, Q.; Yi, H.; Liang, H.; Deng, Y.; et al. Analysis of B Cell Receptor Repertoires Reveals Key Signatures of the Systemic B Cell Response after SARS-CoV-2 Infection. J. Virol. 2022, 96, Jvi0160021. [Google Scholar] [CrossRef]
- Ji, S.-Q.; Zhang, M.; Zhang, Y.; Xia, K.; Chen, Y.; Chu, Q.; Wei, Y.-C.; Zhou, F.-L.; Bu, B.-T.; Tu, H.-L.; et al. Characteristics of immune and inflammatory responses among different age groups of pediatric patients with COVID-19 in China. World J. Pediatr. 2021, 17, 375–384. [Google Scholar] [CrossRef]
- Britton, G.J.; Chen-Liaw, A.; Cossarini, F.; Livanos, A.E.; Spindler, M.P.; Plitt, T.; Eggers, J.; Mogno, I.; Gonzalez-Reiche, A.S.; Siu, S.; et al. Limited intestinal inflammation despite diarrhea, fecal viral RNA and SARS-CoV-2-specific IgA in patients with acute COVID-19. Sci. Rep. 2021, 11, 13308. [Google Scholar] [CrossRef] [PubMed]
- Arias, N.; Arboleya, S.; Allison, J.; Kaliszewska, A.; Higarza, S.; Gueimonde, M.; Arias, J. The Relationship between Choline Bioavailability from Diet, Intestinal Microbiota Composition, and Its Modulation of Human Diseases. Nutrients 2020, 12, 2340. [Google Scholar] [CrossRef] [PubMed]
- Rojo, D.; Gosalbes, M.J.; Ferrari, R.; Pérez-Cobas, A.E.; Hernández, E.; Oltra, R.; Buesa, J.; Latorre, A.; Barbas, C.; Ferrer, M.; et al. Clostridium difficile heterogeneously impacts intestinal community architecture but drives stable metabolome responses. ISME J. 2015, 9, 2206–2220. [Google Scholar] [CrossRef]
- Prentice, M.B. Bacterial microcompartments and their role in pathogenicity. Curr. Opin. Microbiol. 2021, 63, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Bai, Y.; Zhou, J.; Huang, W.; Yan, J.; Tao, J.; Fan, Q.; Liu, Y.; Mei, D.; Yan, Q.; et al. Core Fucosylation of Maternal Milk N-Glycan Evokes B Cell Activation by Selectively Promoting the l -Fucose Metabolism of Gut Bifidobacterium spp. and Lactobacillus spp. Mbio 2019, 10, e00128-19. [Google Scholar] [CrossRef]
- Man, W.H.; Clerc, M.; Piters, W.D.S.; Van Houten, M.A.; Chu, M.L.J.N.; Kool, J.; Keijser, B.J.F.; Sanders, E.A.M.; Bogaert, D. Loss of Microbial Topography between Oral and Nasopharyngeal Microbiota and Development of Respiratory Infections Early in Life. Am. J. Respir. Crit. Care Med. 2019, 200, 760–770. [Google Scholar] [CrossRef]
- Aparicio, M.; Alba, C.; Proctocolitis Study Group of CAM Public Health Area 6 Proctocolitis Study Group of CAM Public Health Area 6; Rodríguez, J.M.; Fernández, L. Microbiological and Immunological Markers in Milk and Infant Feces for Common Gastrointestinal Disorders: A Pilot Study. Nutrients 2020, 12, 634. [Google Scholar] [CrossRef]
- Niu, H.; Zhou, X.; Zhang, X.; Liu, T.; Wu, Y.; Lyu, L.; Liang, C.; Chen, S.; Gong, P.; Zhang, J.; et al. Breast milk contains probiotics with anti-infantile diarrhoea effects that may protect infants as they change to solid foods. Environ. Microbiol. 2021, 23, 1750–1764. [Google Scholar] [CrossRef]
- Zamakhchari, M.; Wei, G.; Dewhirst, F.; Lee, J.; Schuppan, D.; Oppenheim, F.G.; Helmerhorst, E.J. Identification of Rothia Bacteria as Gluten-Degrading Natural Colonizers of the Upper Gastro-Intestinal Tract. PLoS ONE 2011, 6, e24455. [Google Scholar] [CrossRef]
- Wu, Y.; Cheng, X.; Jiang, G.; Tang, H.; Ming, S.; Tang, L.; Lu, J.; Guo, C.; Shan, H.; Huang, X. Altered oral and gut microbiota and its association with SARS-CoV-2 viral load in COVID-19 patients during hospitalization. npj Biofilms Microbiomes 2021, 7, 61. [Google Scholar] [CrossRef]
- Gao, B.; Gallagher, T.; Zhang, Y.; Elbadawi-Sidhu, M.; Lai, Z.; Fiehn, O.; Whiteson, K.L. Tracking Polymicrobial Metabolism in Cystic Fibrosis Airways: Pseudomonas aeruginosa Metabolism and Physiology Are Influenced by Rothia mucilaginosa-Derived Metabolites. Msphere 2018, 3, e00151-18. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.W.; Schmieder, R.; Haynes, M.; Furlan, M.; Matthews, T.D.; Whiteson, K.; Poole, S.J.; Hayes, C.S.; Low, D.A.; Maughan, H.; et al. Mechanistic Model of Rothia mucilaginosa Adaptation toward Persistence in the CF Lung, Based on a Genome Reconstructed from Metagenomic Data. PLoS ONE 2013, 8, e64285. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.C.; Freedman, R.; Law, A.J.; Clark, A.M.; Hunter, S.K. Maternal nutrients and effects of gestational COVID-19 infection on fetal brain development. Clin. Nutr. ESPEN 2021, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Indirect Effect | Pathway | |||||
|---|---|---|---|---|---|---|
| Birth Event (X) | Mediator1 (M1) | Mediator2 (M2) | Beta-Coefficiency (95% CI) | Path | Mediator | Beta-Coefficiency (95% CI) |
| CS-labor | Increased dose of | Galactose | −0.17 (−0.34, −0.05) | a | M1 | −0.48 (−0.79, −0.16) |
| breast milk | a′ | M2 | 0.07 (−1.63, 1.78) | |||
| d | M1-M2 | 1.62 (1.03, 2.21) | ||||
| b | M1 | 1.80 (1.17, 2.43) | ||||
| b′ | M2 | 0.22 (0.10, 0.33) | ||||
| c | CS-labor | −0.49 (−2.25, 1.26) | ||||
| CS-labor | Increased dose of | Fucose | −0.13 (−0.35, −0.03) | a | M1 | −0.42 (−0.74, −0.10) |
| breast milk | a′ | M2 | −1.60 (−4.74, 1.54) | |||
| d | M1-M2 | 2.34 (1.28, 3.40) | ||||
| b | M1 | 1.83 (1.21, 2.45) | ||||
| b′ | M2 | 0.13 (0.07, 0.20) | ||||
| c | CS-labor | −0.40 (−2.19, 1.39) | ||||
| CS-labor | Increased dose of | 4Aminobutyrate | −0.09 (−0.23, −0.01) | a | M1 | −0.45 (−0.80, −0.10) |
| breast milk | (GABA) | a′ | M2 | −0.39 (−1.21, 0.42) | ||
| d | M1-M2 | 0.48 (0.21, 0.75) | ||||
| b | M1 | 2.03 (1.36, 2.69) | ||||
| b′ | M2 | 0.41 (0.12, 0.69) | ||||
| c | CS-labor | −0.87 (−2.83, 1.09) | ||||
| CS-labor | Increased dose of | Choline | −0.10 (−0.27, 0.00) | a | M1 | −0.23 (−0.46, 0.00) |
| breast milk | a′ | M2 | 0.00 (−0.09, 0.09) | |||
| d | M1-M2 | 0.11 (0.08, 0.15) | ||||
| b | M1 | 1.98 (1.29, 2.68) | ||||
| b′ | M2 | 3.87 (2.15, 5.59) | ||||
| c | CS-labor | −0.85 (−2.59, 0.88) | ||||
| CS-labor | unclassified | Pyruvate | 0.20 (0.06, 0.45) | a | M1 | 0.64 (0.24, 1.04) |
| Enterobacteriaceae | a′ | M2 | −0.56 (−1.35, 0.23) | |||
| d | M1-M2 | 0.45 (0.28, 0.63) | ||||
| b | M1 | 0.64 (0.26, 1.03) | ||||
| b′ | M2 | 0.68 (0.49, 0.87) | ||||
| c | CS-labor | −1.60 (−3.32, 0.12) | ||||
| CS-labor | unclassified | 1,2-propanediol | 0.15 (0.04, 0.33) | a | M1 | 0.63 (0.21, 1.04) |
| Enterobacteriaceae | a′ | M2 | −1.43 (−3.09, 0.24) | |||
| d | M1-M2 | 0.93 (0.56, 1.30) | ||||
| b | M1 | 0.71 (0.22, 1.20) | ||||
| b′ | M2 | 0.26 (0.14, 0.38) | ||||
| c | CS-labor | −2.06 (−4.22, 0.10) | ||||
| CS-labor | unclassified | Choline | 0.15 (0.04, 0.33) | a | M1 | 0.64 (0.24, 1.05) |
| Enterobacteriaceae | a′ | M2 | −0.05 (−0.15, 0.04) | |||
| d | M1-M2 | 0.05 (0.03, 0.07) | ||||
| b | M1 | 0.70 (0.30, 1.10) | ||||
| b′ | M2 | 4.57 (2.84, 6.30) | ||||
| c | CS-labor | −1.74 (−3.53, 0.04) | ||||
| CS-labor | unclassified | Lactate | 0.09 (0.02, 0.19) | a | M1 | 0.60 (0.20, 0.99) |
| Enterobacteriaceae | a′ | M2 | −5.94 (−12.12, 0.24) | |||
| d | M1-M2 | 2.63 (1.26, 4.00) | ||||
| b | M1 | 0.78 (0.35, 1.22) | ||||
| b′ | M2 | 0.06 (0.03, 0.09) | ||||
| c | CS-labor | −1.77 (−3.70, 0.17) | ||||
| CS-labor | C. difficile | 1,2-propanediol | −0.12 (−0.30, −0.03) | a | M1 | 4.16 (1.78, 6.53) |
| a′ | M2 | −0.09 (−1.96, 1.77) | ||||
| d | M1-M2 | −0.11 (−0.18, −0.03) | ||||
| b | M1 | −0.05 (−0.15, 0.04) | ||||
| b′ | M2 | 0.28 (0.16, 0.40) | ||||
| c | CS-labor | −1.1 (−3.51, 1.31) | ||||
| CS-labor | C. difficile | Pyruvate | −0.12 (−0.31, −0.02) | a | M1 | 3.73 (1.44, 6.02) |
| a′ | M2 | 0.05 (−0.87, 0.97) | ||||
| d | M1-M2 | −0.04 (−0.08, −0.01) | ||||
| b | M1 | −0.05 (−0.13, 0.03) | ||||
| b′ | M2 | 0.73 (0.53, 0.93) | ||||
| c | CS-labor | −0.91 (−2.89, 1.08) | ||||
| CS-labor | C. difficile | Choline | −0.12 (−0.27, −0.02) | a | M1 | 3.52 (1.25, 5.80) |
| a′ | M2 | 0.00 (−0.10, 0.11) | ||||
| d | M1-M2 | −0.01 (−0.01, 0.00) | ||||
| b | M1 | −0.04 (−0.12, 0.04) | ||||
| b′ | M2 | 5.20 (3.40, 7.00) | ||||
| c | CS-labor | −0.99 (−3.03, 1.05) | ||||
| CS-labor | C. difficile | Lactate | −0.09 (−0.20, −0.01) | a | M1 | 3.10 (0.81, 5.38) |
| a′ | M2 | −1.74 (−8.70, 5.22) | ||||
| d | M1-M2 | −0.44 (−0.72, −0.17) | ||||
| b | M1 | −0.05 (−0.14, 0.04) | ||||
| b′ | M2 | 0.06 (0.03, 0.09) | ||||
| c | CS-labor | −1.02 (−3.23, 1.19) | ||||
| CS-labor | Streptococcus | Lactate | 0.09 (0.00, 0.23) | a | M1 | 0.47 (0.03, 0.91) |
| a′ | M2 | −5.85 (−11.94, 0.24) | ||||
| d | M1-M2 | 3.13 (1.93, 4.33) | ||||
| b | M1 | 0.20 (−0.19, 0.60) | ||||
| b′ | M2 | 0.06 (0.04, 0.09) | ||||
| c | CS-labor | −1.37 (−3.32, 0.58) | ||||
| CS-no labor | unclassified | Pyruvate | 0.15 (0.01, 0.39) | a | M1 | 0.48 (0.10, 0.86) |
| Enterobacteriaceae | a′ | M2 | −0.61 (−1.36, 0.15) | |||
| d | M1-M2 | 0.45 (0.28, 0.63) | ||||
| b | M1 | 0.64 (0.26, 1.03) | ||||
| b′ | M2 | 0.68 (0.49, 0.87) | ||||
| c | CS-no labor | 0.29 (−1.35, 1.93) | ||||
| CS-no labor | unclassified | Choline | 0.11 (0.01, 0.26) | a | M1 | 0.46 (0.08, 0.84) |
| Enterobacteriaceae | a′ | M2 | −0.08 (−0.16, 0.01) | |||
| d | M1-M2 | 0.05 (0.03, 0.07) | ||||
| b | M1 | 0.70 (0.30, 1.10) | ||||
| b′ | M2 | 4.57 (2.84, 6.30) | ||||
| c | CS-no labor | 0.27 (−1.41, 1.95) | ||||
| CS-no labor | unclassified | 1,2-propanediol | 0.09 (0.01, 0.26) | a | M1 | 0.40 (0.04, 0.76) |
| Enterobacteriaceae | a′ | M2 | −1.38 (−2.84, 0.08) | |||
| d | M1-M2 | 0.93 (0.56, 1.30) | ||||
| b | M1 | 0.71 (0.22, 1.20) | ||||
| b′ | M2 | 0.26 (0.14, 0.38) | ||||
| c | CS-no labor | 0.14 (−1.75, 2.03) | ||||
| CS-no labor | unclassified | Lactate | 0.07 (0.01, 0.16) | a | M1 | 0.46 (0.10, 0.81) |
| Enterobacteriaceae | a′ | M2 | −6.22 (−11.82, −0.61) | |||
| d | M1-M2 | 2.63 (1.26, 4.00) | ||||
| b | M1 | 0.78 (0.35, 1.22) | ||||
| b′ | M2 | 0.06 (0.03, 0.09) | ||||
| c | CS-no labor | −0.22 (−1.98, 1.53) | ||||
| CS-no labor | C. difficile | 1,2-propanediol | −0.11 (−0.26, −0.02) | a | M1 | 3.68 (1.66, 5.71) |
| a′ | M2 | −0.44 (−2.03, 1.15) | ||||
| d | M1-M2 | −0.11 (−0.18, −0.03) | ||||
| b | M1 | −0.05 (−0.15, 0.04) | ||||
| b′ | M2 | 0.28 (0.16, 0.40) | ||||
| c | CS-no labor | 0.58 (−1.47, 2.64) | ||||
| CS-no labor | C. difficile | Pyruvate | −0.11 (−0.28, −0.01) | a | M1 | 3.38 (1.30, 5.45) |
| a′ | M2 | −0.30 (−1.14, 0.54) | ||||
| d | M1-M2 | −0.04 (−0.08, −0.01) | ||||
| b | M1 | −0.05 (−0.13, 0.03) | ||||
| b′ | M2 | 0.73 (0.53, 0.93) | ||||
| c | CS-no labor | 0.84 (−0.97, 2.64) | ||||
| CS-no labor | C. difficile | Choline | −0.11 (−0.26, −0.02) | a | M1 | 3.36 (1.31, 5.42) |
| a′ | M2 | −0.02 (−0.12, 0.07) | ||||
| d | M1-M2 | −0.01 (−0.01, 0.00) | ||||
| b | M1 | −0.04 (−0.12, 0.04) | ||||
| b′ | M2 | 5.20 (3.40, 7.00) | ||||
| c | CS-no labor | 0.76 (−1.08, 2.61) | ||||
| CS-no labor | C. difficile | Lactate | −0.09 (−0.19, −0.02) | a | M1 | 3.17 (1.20, 5.15) |
| a′ | M2 | −3.29 (−9.33, 2.76) | ||||
| d | M1-M2 | −0.44 (−0.72, −0.17) | ||||
| b | M1 | −0.05 (−0.14, 0.04) | ||||
| b′ | M2 | 0.06 (0.03, 0.09) | ||||
| c | CS-no labor | 0.27 (−1.65, 2.19) | ||||
| CS-no labor | Streptococcus | Lactate | 0.11 (0.04, 0.24) | a | M1 | 0.58 (0.17, 0.98) |
| a′ | M2 | −6.82 (−12.36, −1.27) | ||||
| d | M1-M2 | 3.13 (1.93, 4.33) | ||||
| b | M1 | 0.20 (−0.19, 0.60) | ||||
| b′ | M2 | 0.06 (0.04, 0.09) | ||||
| c | CS-no labor | 0.04 (−1.74, 1.82) | ||||
| Vaginal-IAP | Rothia | Pyruvate | 0.12 (0.01, 0.33) | a | M1 | 0.77 (0.09, 1.45) |
| a′ | M2 | −0.45 (−1.09, 0.19) | ||||
| d | M1-M2 | 0.23 (0.15, 0.32) | ||||
| b | M1 | 0.28 (0.09, 0.46) | ||||
| b′ | M2 | 0.68 (0.49, 0.88) | ||||
| c | Vaginal-IAP | −0.51 (−1.91, 0.89) | ||||
| Vaginal-IAP | Rothia | Lactate | 0.08 (0.01, 0.17) | a | M1 | 0.77 (0.08, 1.46) |
| a′ | M2 | −6.75 (−11.74, −1.76) | ||||
| d | M1-M2 | 1.76 (1.13, 2.38) | ||||
| b | M1 | 0.35 (0.14, 0.55) | ||||
| b′ | M2 | 0.06 (0.03, 0.08) | ||||
| c | Vaginal-IAP | −0.56 (−2.15, 1.04) | ||||
| Vaginal-IAP | Rothia | 1,2-propanediol | 0.05 (0.00, 0.15) | a | M1 | 0.72 (−0.01, 1.45) |
| a′ | M2 | −0.58 (−1.93, 0.78) | ||||
| d | M1-M2 | 0.26 (0.09, 0.44) | ||||
| b | M1 | 0.38 (0.16, 0.60) | ||||
| b′ | M2 | 0.27 (0.15, 0.38) | ||||
| c | Vaginal-IAP | −0.83 (−2.55, 0.89) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.Y.; Tun, H.M.; Field, C.J.; Mandhane, P.J.; Moraes, T.J.; Simons, E.; Turvey, S.E.; Subbarao, P.; Scott, J.A.; Kozyrskyj, A.L. Impact of Cesarean Delivery and Breastfeeding on Secretory Immunoglobulin A in the Infant Gut Is Mediated by Gut Microbiota and Metabolites. Metabolites 2023, 13, 148. https://doi.org/10.3390/metabo13020148
Chen YY, Tun HM, Field CJ, Mandhane PJ, Moraes TJ, Simons E, Turvey SE, Subbarao P, Scott JA, Kozyrskyj AL. Impact of Cesarean Delivery and Breastfeeding on Secretory Immunoglobulin A in the Infant Gut Is Mediated by Gut Microbiota and Metabolites. Metabolites. 2023; 13(2):148. https://doi.org/10.3390/metabo13020148
Chicago/Turabian StyleChen, Yuan Yao, Hein M. Tun, Catherine J. Field, Piushkumar J. Mandhane, Theo J. Moraes, Elinor Simons, Stuart E. Turvey, Padmaja Subbarao, James A. Scott, and Anita L. Kozyrskyj. 2023. "Impact of Cesarean Delivery and Breastfeeding on Secretory Immunoglobulin A in the Infant Gut Is Mediated by Gut Microbiota and Metabolites" Metabolites 13, no. 2: 148. https://doi.org/10.3390/metabo13020148
APA StyleChen, Y. Y., Tun, H. M., Field, C. J., Mandhane, P. J., Moraes, T. J., Simons, E., Turvey, S. E., Subbarao, P., Scott, J. A., & Kozyrskyj, A. L. (2023). Impact of Cesarean Delivery and Breastfeeding on Secretory Immunoglobulin A in the Infant Gut Is Mediated by Gut Microbiota and Metabolites. Metabolites, 13(2), 148. https://doi.org/10.3390/metabo13020148