
Comparative Analysis of the Temporal Impacts of Corticosterone and Simulated Production Stressors on the Metabolome of Broiler Chickens

 , , , and
, , , and 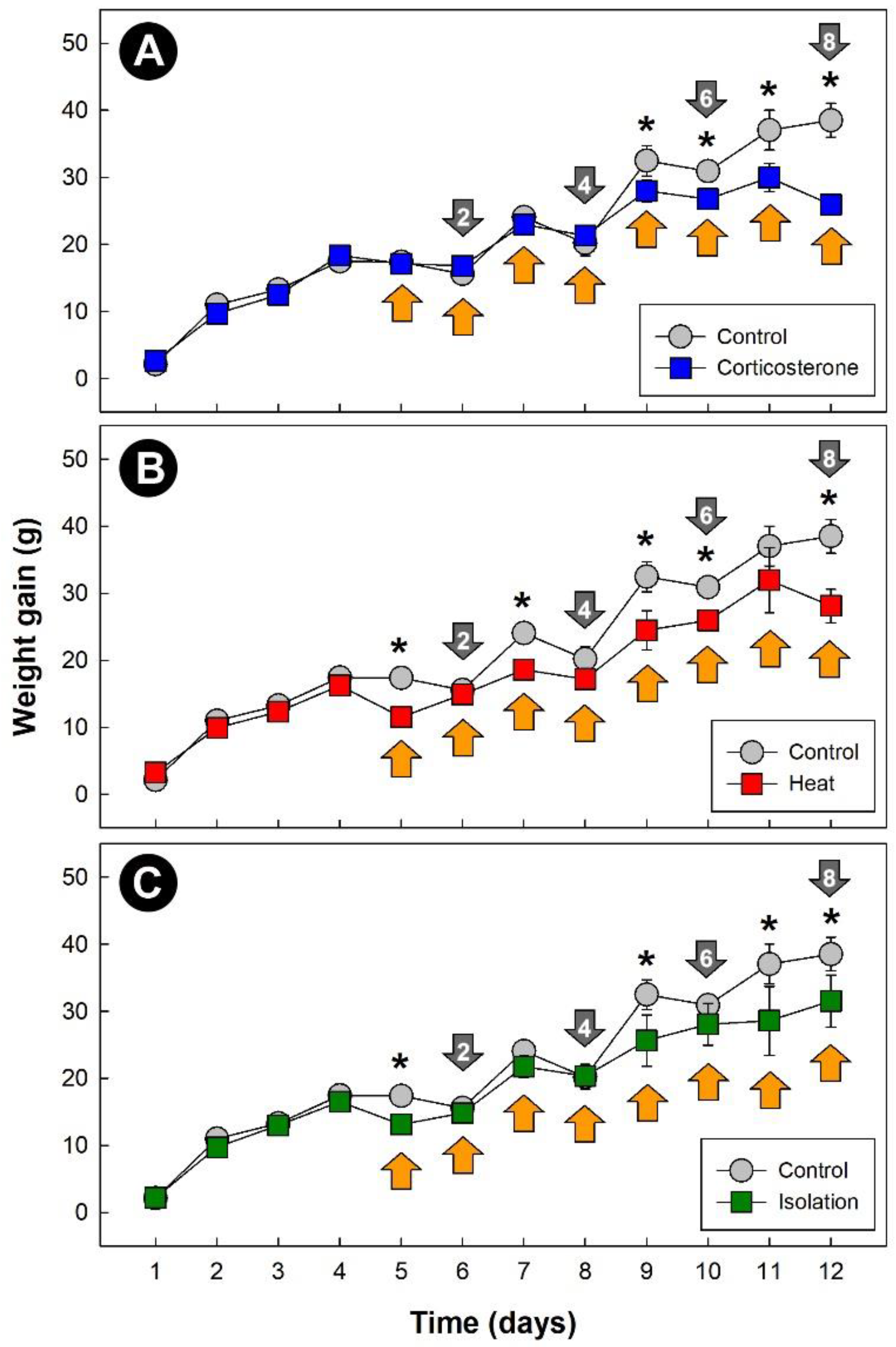{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Bird Husbandry
2.3. Enteric Microbiota Establishment
2.4. Stressors
2.5. Sample Collection
2.6. Sample Preparation for NMR Spectroscopy
2.7. NMR Spectroscopy Data Acquisition and Processing
2.8. Statistical Analyses
3. Results
3.1. Stressors Altered Weight Gain in Broiler Chickens
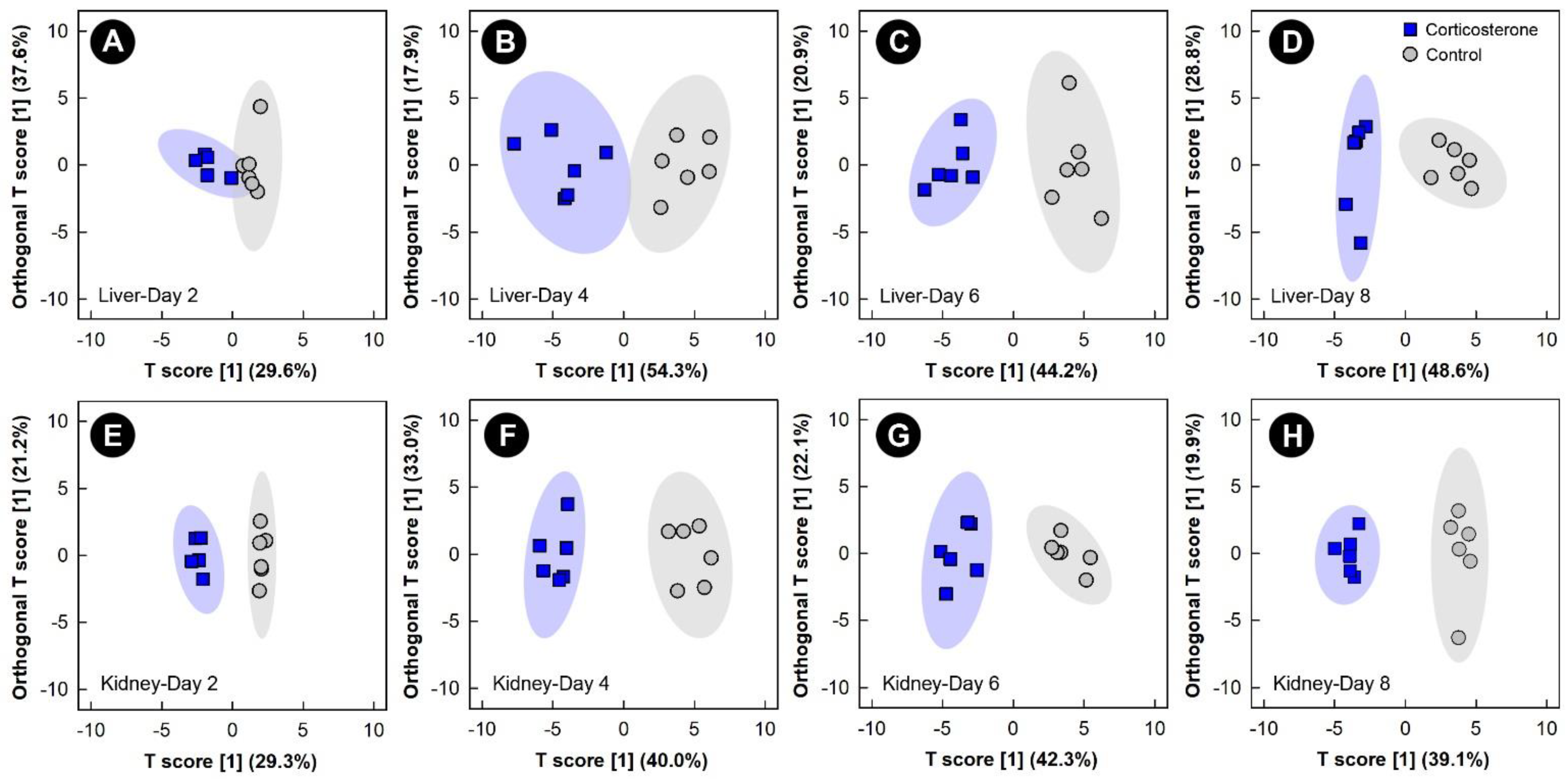
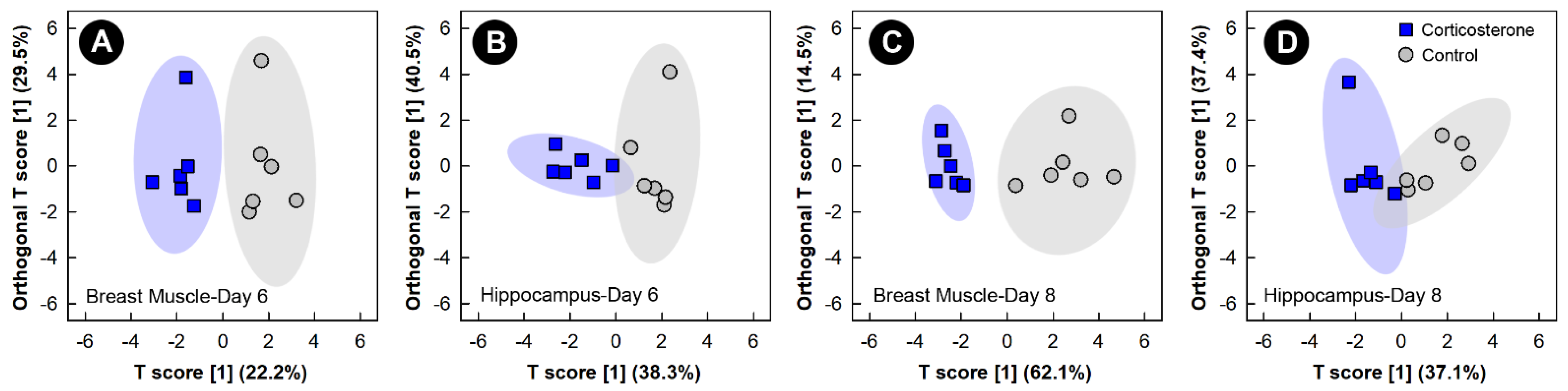
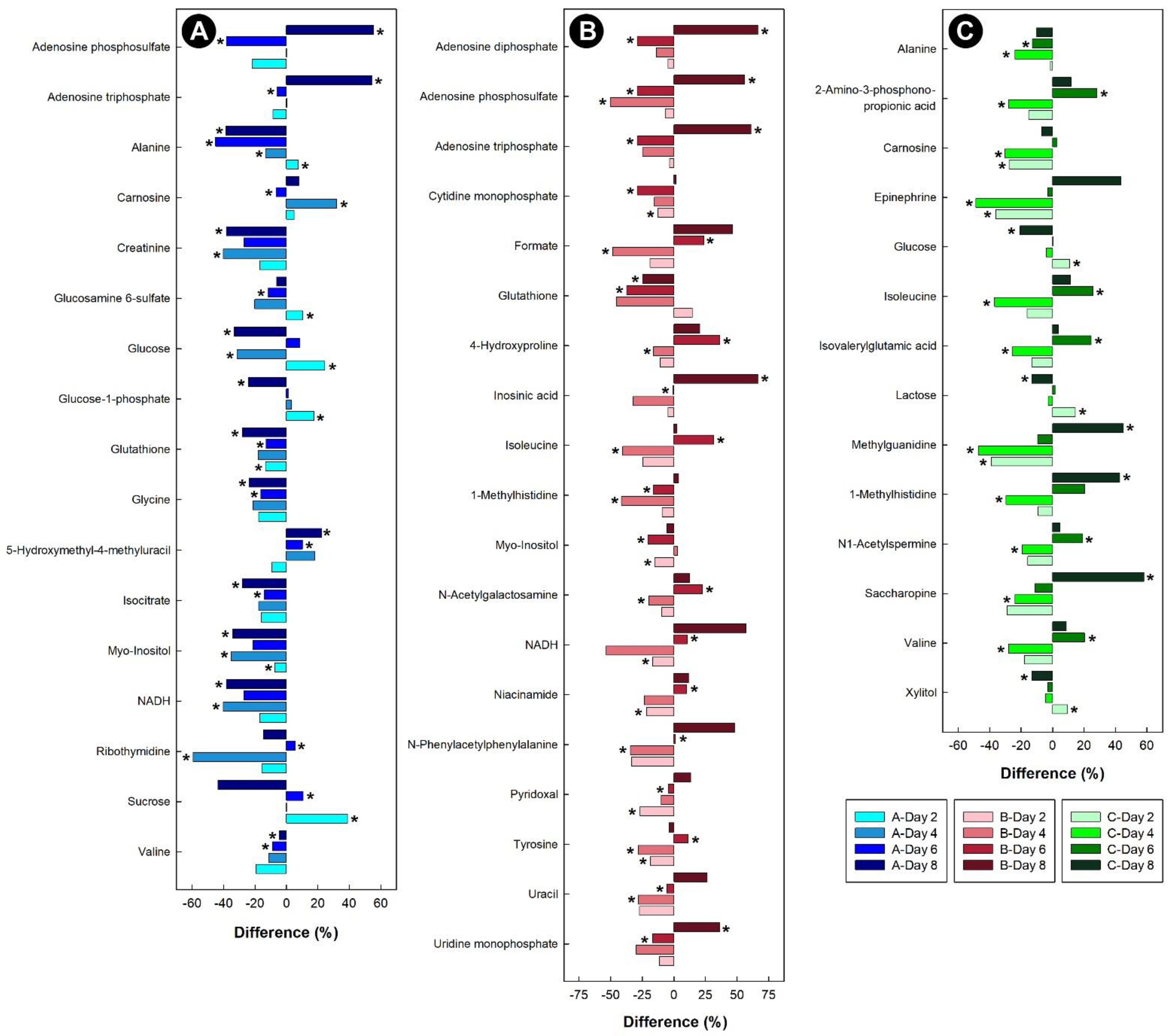
3.2. Corticosterone Administration Substantively Altered the Liver, Kidney, Breast Muscle, and Hippocampus Metabolome
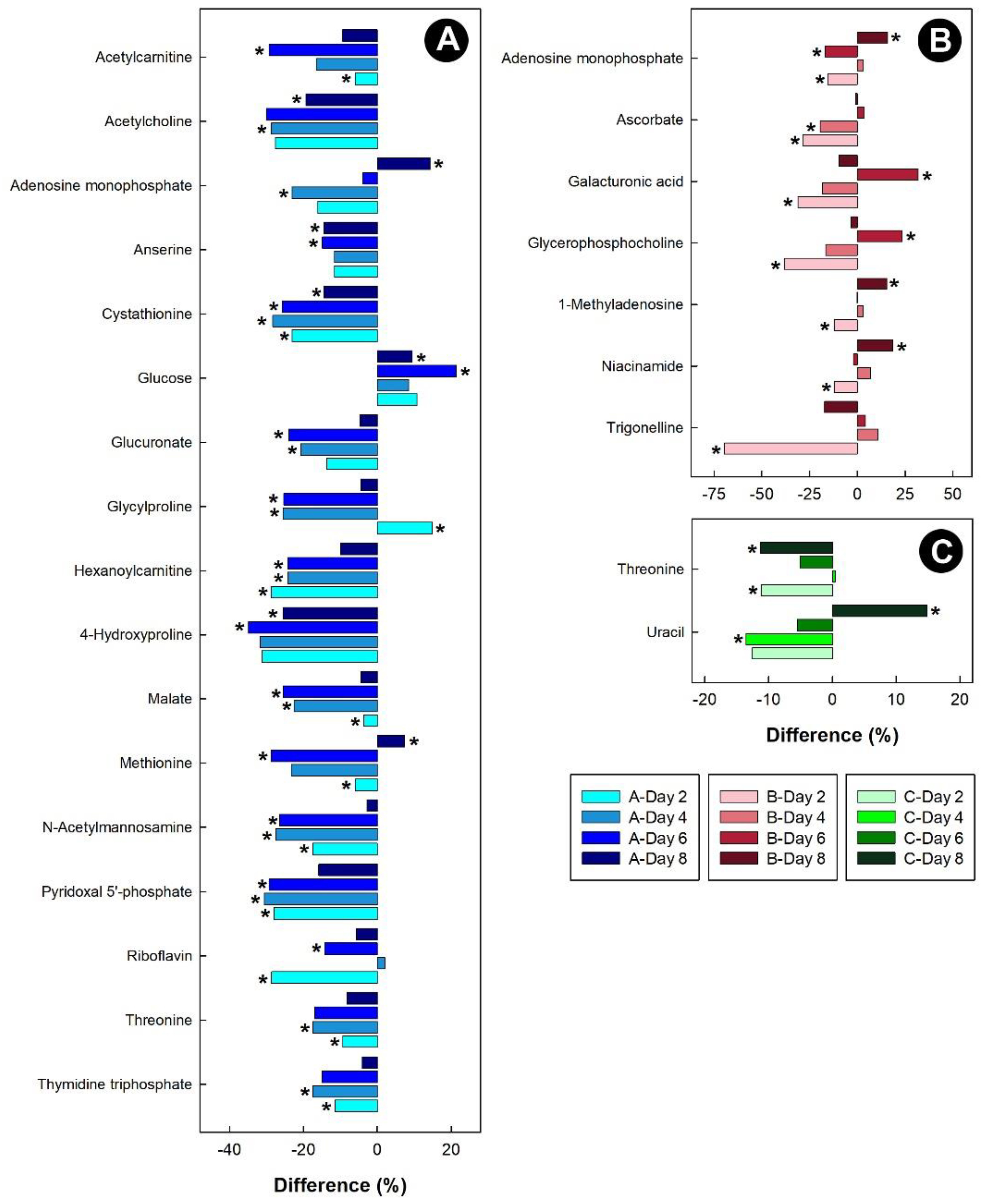
3.3. Heat Stress Altered the Liver, Kidney, and Hippocampus Metabolome, but Not the Breast Muscle Metabolome
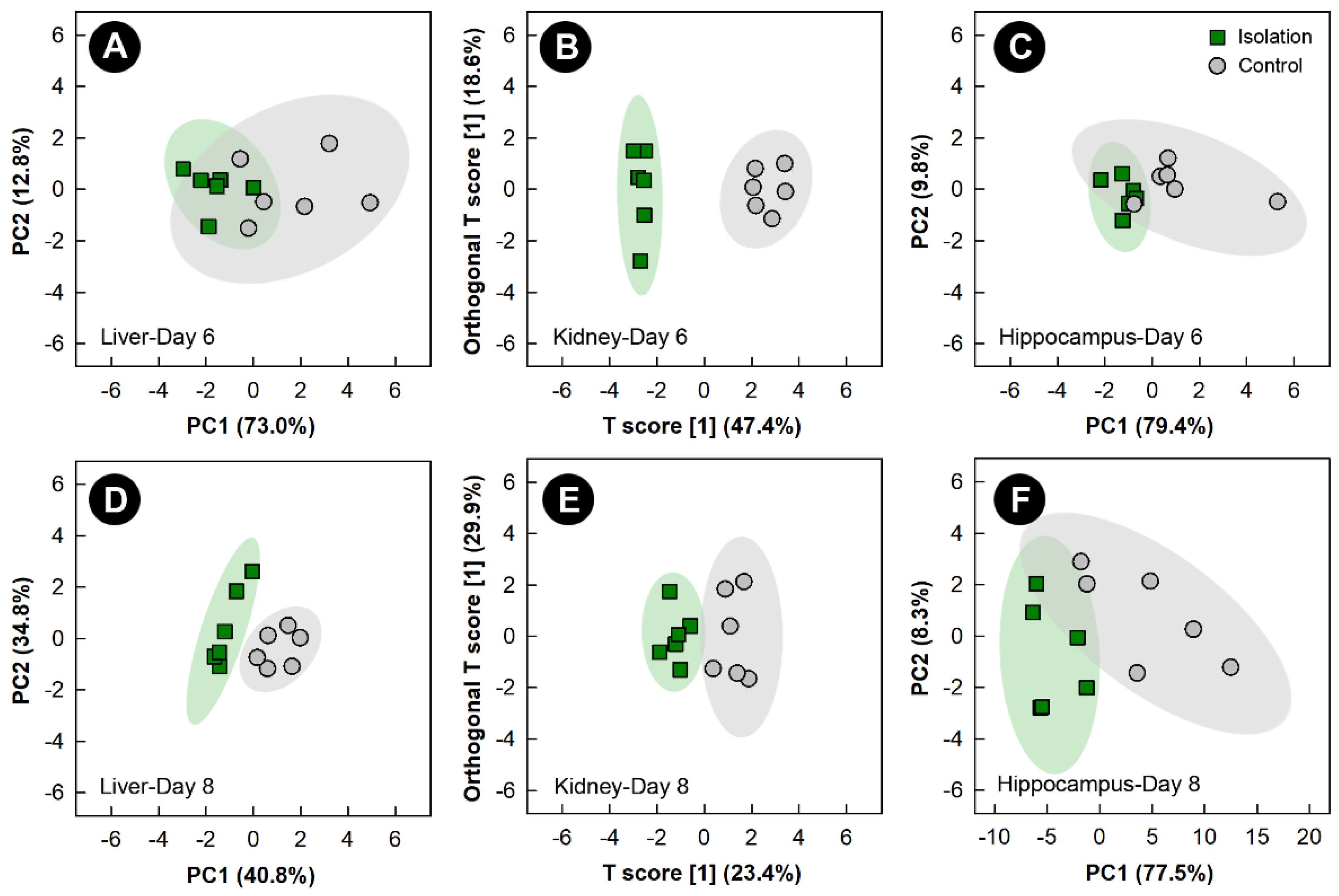
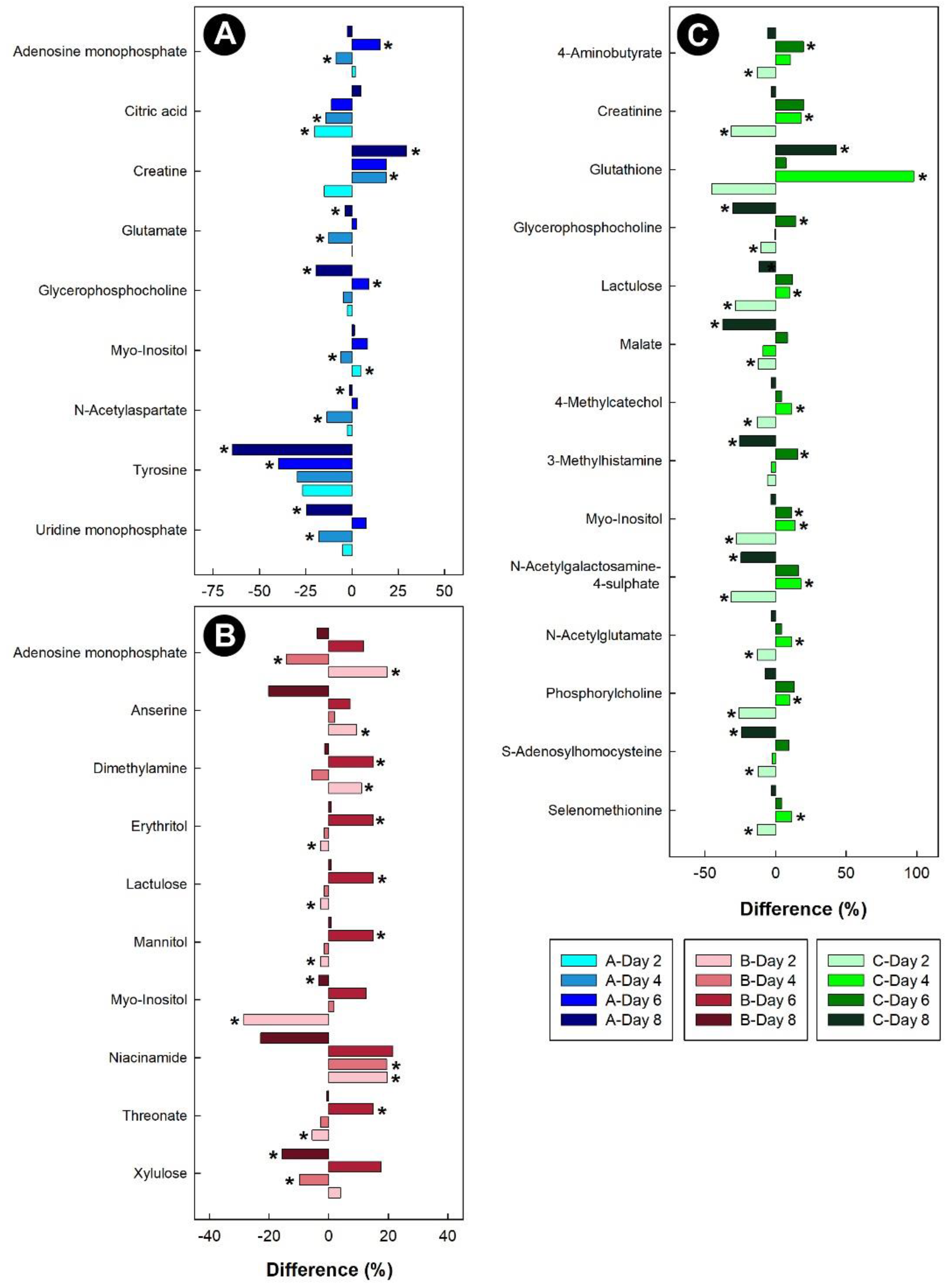
3.4. Isolation Stress Altered the Liver, Kidney, and Hippocampus Metabolome, but Not the Breast Muscle Metabolome
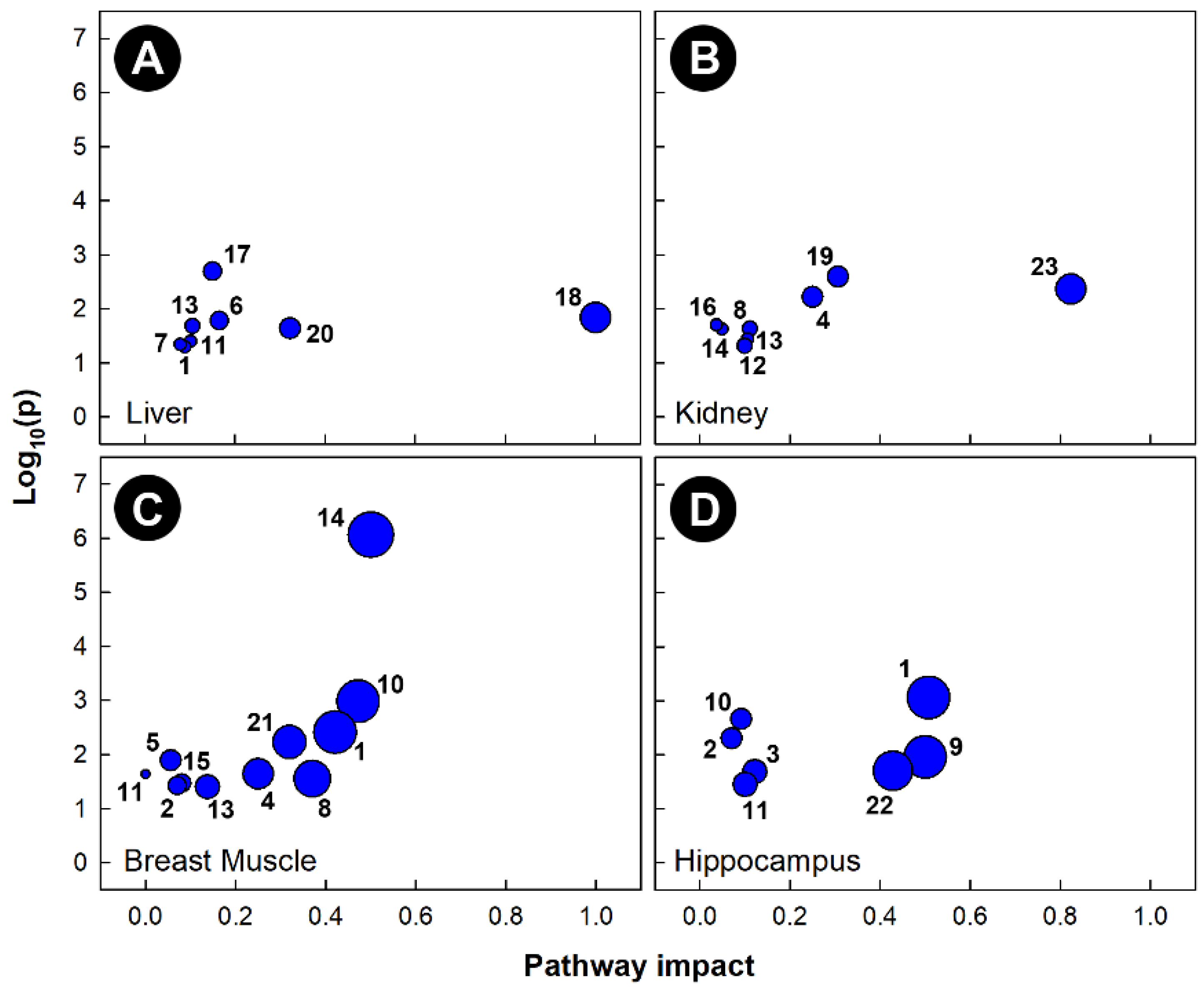
3.5. Different Metabolic Pathways Were Altered by Stressors
3.6. Stressors Altered Relative Metabolite Concentrations in Tissues
4. Discussion
4.1. Overview
4.2. Glycine
4.3. Alanine
4.4. Glutathione
4.5. Threonine
4.6. Betaine
4.7. Breast Muscle Metabolome
4.8. Hippocampus Metabolome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Virden, W.S.; Kidd, M.T. Physiological stress in broilers: Ramifications on nutrient digestibility and responses. J. Appl. Poult. Res. 2009, 18, 338–347. [Google Scholar] [CrossRef]
- Jastrebski, S.F.; Lamont, S.J.; Schmidt, C.J. Chicken hepatic response to chronic heat stress using integrated transcriptome and metabolome analysis. PLoS ONE 2017, 12, e0181900. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Shini, A.; Huff, G.R. Effects of chronic and repeated corticosterone administration in rearing chickens on physiology, the onset of lay and egg production of hens. Physiol. Behav. 2009, 98, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Sandercock, D.A.; Hunter, R.R.; Nute, G.R.; Mitchell, M.A.; Hocking, P.M. Acute heat stress-induced alterations in blood acid-base status and skeletal muscle membrane integrity in broiler chickens at two ages: Implications for meat quality. Poult. Sci. 2001, 80, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef]
- Zaytsoff, S.J.M.; Brown, C.L.J.; Montina, T.; Metz, G.A.S.; Abbott, D.W.; Uwiera, R.R.E.; Inglis, G.D. Corticosterone-mediated physiological stress modulates hepatic lipid metabolism, metabolite profiles, and systemic responses in chickens. Sci. Rep. 2019, 9, 19225–19238. [Google Scholar] [CrossRef]
- El-Lethey, H.; Huber-Eicher, B.; Jungi, T.W. Exploration of stress-induced immunosuppression in chickens reveals both stress-resistant and stress-susceptible antigen responses. Vet. Immunol. Immunopathol. 2003, 95, 91–101. [Google Scholar] [CrossRef]
- Dai, S.F.; Wang, L.K.; Wen, A.Y.; Wang, L.X.; Jin, G.M. Dietary glutamine supplementation improves growth performance, meat quality and colour stability of broilers under heat stress. Br. Poult. Sci. 2009, 50, 333–340. [Google Scholar] [CrossRef]
- Dong, H.; Lin, H.; Jiao, H.C.; Song, Z.G.; Zhao, J.P.; Jiang, K.J. Altered development and protein metabolism in skeletal muscles of broiler chickens (Gallus gallus domesticus) by corticosterone. Comp. Biochem. Physiol. A. Physiol. 2007, 147, 189–195. [Google Scholar] [CrossRef]
- Zaytsoff, S.J.M.; Lyons, S.M.; Garner, A.M.; Uwiera, R.R.E.; Zandberg, W.F.; Abbott, D.W.; Inglis, G.D. Host responses to Clostridium perfringens challenge in a chicken model of chronic stress. Gut Pathog. 2020, 12, 12–24. [Google Scholar] [CrossRef]
- Zaytsoff, S.J.M.; Boras, V.F.; Uwiera, R.R.E.; Inglis, G.D. A stress-induced model of acute necrotic enteritis in broiler chickens using dietary corticosterone administration. Poult. Sci. 2022, 101, 101726. [Google Scholar] [CrossRef]
- Guabiraba-Brito, R.; Schouler, C. Avian colibacillosis: Still many black holes. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef]
- Weimer, S.L.; Wideman, R.F.; Scanes, C.G.; Mauromoustakos, A.; Christensen, K.D.; Vizzier-Thaxton, Y. An evaluation of methods for measuring stress in broiler chickens. Poult. Sci. 2018, 97, 3381–3389. [Google Scholar] [CrossRef]
- Nwaigwe, C.U.; Ihedioha, J.I.; Shoyinka, S.V.; Nwaigwe, C.O. Evaluation of the hematological and clinical biochemical markers of stress in broiler chickens. Vet. World 2020, 13, 2294–2300. [Google Scholar] [CrossRef]
- Bortolotti, G.R.; Marchant, T.A.; Blas, J.; German, T. Corticosterone in feathers is a long-term, integrated measure of avian stress physiology. Funct. Ecol. 2008, 22, 494–500. [Google Scholar] [CrossRef]
- Blas, J. Stress in birds. In Sturkie’s Avian Physiology, 6th ed.; Scanes, Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 769–810. [Google Scholar]
- Alm, M.; Holm, L.; Tauson, R.; Wall, H. Corticosterone metabolites in laying hen droppings—Effects of fiber enrichment, genotype, and daily variations. Poult. Sci. 2014, 93, 2615–2621. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Coleman, G.J.; Barnett, J.L.; Jones, R.B. Behavioral responses to humans and the productivity of commercial broiler chickens. Appl. Anim. Behav. Sci. 1994, 41, 101–114. [Google Scholar] [CrossRef]
- Kannan, G.; Mench, J.A. Influence of different handling methods and crating periods on plasma corticosterone concentrations in broilers. Br. Poult. Sci. 1996, 37, 21–31. [Google Scholar] [CrossRef]
- Mormède, P.; Andanson, S.; Aupérin, B.; Beerda, B.; Guémené, D.; Malmkvist, J.; Manteca, X.; Manteuffel, G.; Prunet, P.; Van Reenen, C.G.; et al. Exploration of the hypothalamic–pituitary–adrenal function as a tool to evaluate animal welfare. Physiol. Behav. 2007, 92, 317–339. [Google Scholar] [CrossRef]
- Chloupek, P.; Bedanova, I.; Chloupek, J.; Vecerek, V. Changes in selected biochemical indices resulting from various pre-sampling handling techniques in broilers. Acta Vet. Scand. 2011, 53, 31. [Google Scholar] [CrossRef]
- Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/lymphocyte ratio as a measure of stress in chickens. Avian Dis. 1983, 27, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, N.G.C.; Da Silva, L.P.; Fonseca, C.; Soares, A.M.V.M.; Loureiro, S. Habitats: Can they stress birds? An example of the use of biomarkers as an evaluation tool. Integr. Environ. Assess. Manag. 2010, 6, 779–780. [Google Scholar] [CrossRef] [PubMed]
- Karu, N.; Deng, L.; Slae, M.; Guo, A.C.; Sajed, T.; Huynh, H.; Wine, E.; Wishart, D.S. A review on human fecal metabolomics: Methods, applications and the human fecal metabolome database. Anal. Chim. Acta 2018, 1030, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Le Roy, C.I.; Mappley, L.J.; La Ragione, R.M.; Woodward, M.J.; Claus, S.P. NMR-based metabolic characterization of chicken tissues and biofluids: A model for avian research. Metabolomics 2016, 12, 157. [Google Scholar] [CrossRef] [PubMed]
- Abasht, B.; Mutryn, M.F.; Michalek, R.D.; Lee, W.R. Oxidative stress and metabolic perturbations in wooden breast disorder in chickens. PLoS ONE 2016, 11, e0153750. [Google Scholar] [CrossRef]
- Beauclercq, S.; Nadal-Desbarats, L.; Hennequet-Antier, C.; Collin, A.; Tesseraud, S.; Bourin, M.; Le Bihan-Duval, E.; Berri, C. Serum and muscle metabolomics for the prediction of ultimate pH, a key factor for chicken-meat quality. J. Proteome Res. 2016, 15, 1168–1178. [Google Scholar] [CrossRef]
- Beauclercq, S.; Nadal-Desbarats, L.; Hennequet-Antier, C.; Gabriel, I.; Tesseraud, S.; Calenge, F.; Le Bihan-Duval, E.; Mignon-Grasteau, S. Relationships between digestive efficiency and metabolomic profiles of serum and intestinal contents in chickens. Sci. Rep. 2018, 8, 6678–6689. [Google Scholar] [CrossRef]
- Brown, C.L.J.; Inglis, G.D.; Montina, T. Feather pulp: A novel sample for proton nuclear magnetic resonance spectroscopy metabolomics for biomarker discovery. Poult. Sci. 2021, 101, 101866. [Google Scholar] [CrossRef]
- Brown, C.L.J.; Scott, H.; Mulik, C.; Freund, A.S.; Opyr, M.P.; Metz, G.A.S.; Inglis, G.D.; Montina, T. Fecal 1H-NMR metabolomics: A comparison of sample preparation methods for NMR and novel in silico baseline correction. Metabolites 2022, 12, 148. [Google Scholar] [CrossRef]
- Brown, C.L.J.; Zaytsoff, S.J.M.; Montina, T.; Inglis, G.D. Corticosterone-mediated physiological stress alters liver, kidney, and breast muscle metabolomic profiles in chickens. Animals 2021, 11, 3056. [Google Scholar] [CrossRef]
- Inglis, G.D.; Wright, B.D.; Sheppard, S.A.; Abbott, D.W.; Oryschak, M.A.; Montina, T. Expeller-pressed canola (Brassica napus) meal modulates the structure and function of the cecal microbiota, and alters the metabolome of the pancreas, liver, and breast muscle of broiler chickens. Animals 2021, 11, 577. [Google Scholar] [CrossRef]
- Post, J.; Rebel, J.M.; ter Huurne, A.A. Physiological effects of elevated plasma corticosterone concentrations in broiler chickens. An alternative means by which to assess the physiological effects of stress. Poult. Sci. 2003, 82, 1313–1318. [Google Scholar] [CrossRef]
- Virden, W.S.; Thaxton, J.P.; Corzo, A.; Dozier, W.A.; Kidd, M.T. Evaluation of models using corticosterone and adrenocorticotropin to induce conditions mimicking physiological stress in commercial broilers. Poult. Sci. 2007, 86, 2485–2491. [Google Scholar] [CrossRef]
- Shini, S.; Kaiser, P. Effects of stress, mimicked by administration of corticosterone in drinking water, on the expression of chicken cytokine and chemokine genes in lymphocytes. Stress 2009, 12, 388–399. [Google Scholar] [CrossRef]
- Günal, M. The effects of early-age thermal manipulation and daily short-term fasting on performance and body temperatures in broiler exposed to heat stress. J. Anim. Physiol. Anim. Nutr. 2012, 97, 854–860. [Google Scholar] [CrossRef]
- Hamissou Maman, A.; Özlü, S.; Uçar, A.; Elibol, O. Effect of chick body temperature during post-hatch handling on broiler live performance. Poult. Sci. 2019, 98, 244–250. [Google Scholar] [CrossRef]
- Cramer, T.; Rosenberg, T.; Kisliouk, T.; Meiri, N. PARP inhibitor affects long-term heat-stress response via changes in DNA methylation. Neuroscience 2019, 399, 65–76. [Google Scholar] [CrossRef]
- Zaboli, G.-R.; Rahimi, S.; Shariatmadari, F.; Torshizi, M.A.K.; Baghbanzadeh, A.; Mehr, M. Thermal manipulation during Pre and Post-Hatch on thermotolerance of male broiler chickens exposed to chronic heat stress. Poult. Sci. 2017, 96, 478–485. [Google Scholar] [CrossRef]
- Yossifoff, M.; Kisliouk, T.; Meiri, N. Dynamic changes in DNA methylation during thermal control establishment affect CREB binding to the brain-derived neurotrophic factor promoter. Eur. J. Neurosci. 2008, 28, 2267–2277. [Google Scholar] [CrossRef]
- Zaytsoff, S.J.M.; Montina, T.; Boras, V.F.; Brassard, J.; Moote, P.E.; Uwiera, R.R.E.; Inglis, G.D. Microbiota transplantation in day-old broiler chickens ameliorates necrotic enteritis via modulation of the intestinal microbiota and host immune responses. Pathogens 2022, 11, 972. [Google Scholar] [CrossRef]
- Goerlich, V.C.; Nätt, D.; Elfwing, M.; Macdonald, B.; Jensen, P. Transgenerational effects of early experience on behavioral, hormonal and gene expression responses to acute stress in the precocial chicken. Horm. Behav. 2012, 61, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Striedter, G.F. Evolution of the hippocampus in reptiles and birds. J. Comp. Neurol. 2016, 524, 496–517. [Google Scholar] [CrossRef] [PubMed]
- Puelles, L.; Martinez-de-la-Torre, M.; Martinez, S.; Watson, C.; Paxinos, G. The Chick Brain in Stereotaxic Coordinates and Alternate Stains: Featuring Neuromeric Divisions and Mammalian Homologies; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Tiziani, S.; Emwas, A.-H.; Lodi, A.; Ludwig, C.; Bunce, C.M.; Viant, M.R.; Günther, U.L. Optimized metabolite extraction from blood serum for 1H nuclear magnetic resonance spectroscopy. Anal. Biochem. 2008, 377, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Paxman, E.J.; Boora, N.S.; Kiss, D.; Laplante, D.P.; King, S.; Montina, T.; Metz, G.A.S. Prenatal maternal stress from a natural disaster alters urinary metabolomic profiles in project ice storm participants. Sci. Rep. 2018, 8, 12932–12944. [Google Scholar] [CrossRef] [PubMed]
- Kiss, D.; Ambeskovic, M.; Montina, T.; Metz, G.A.S. Stress transgenerationally programs metabolic pathways linked to altered mental health. Cell. Mol. Life Sci. 2016, 73, 4547–4557. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.E.; Mahle, D.A.; Doom, T.E.; Reo, N.V.; Delraso, N.J.; Raymer, M.L. Dynamic adaptive binning: An improved quantification technique for NMR spectroscopic data. Metabolomics 2011, 7, 179–190. [Google Scholar] [CrossRef]
- Goodpaster, A.M.; Romick-Rosendale, L.E.; Kennedy, M.A. Statistical significance analysis of nuclear magnetic resonance-based metabonomics data. Anal. Biochem. 2010, 401, 134–143. [Google Scholar] [CrossRef]
- Yun, Y.-H.; Liang, F.; Deng, B.-C.; Lai, G.-B.; Vicente Gonçalves, C.M.; Lu, H.-M.; Yan, J.; Huang, X.; Yi, L.-Z.; Liang, Y.-Z. Informative metabolites identification by variable importance analysis based on random variable combination. Metabolomics 2015, 11, 1539–1551. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.J.; van Duijnhoven, J.P.M.; van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. MetPA: A web-based metabolomics tool for pathway analysis and visualization. Bioinformatics 2010, 26, 2342–2344. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Markley, J.L.; Bruschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr. Opin. Biotechnol. 2017, 43, 34–40. [Google Scholar] [CrossRef]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR Spectroscopy for Metabolomics Research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef]
- Emwas, A.-H.M.; Salek, R.M.; Griffin, J.L.; Merzaban, J. NMR-based metabolomics in human disease diagnosis: Applications, limitations, and recommendations. Metabolomics 2013, 9, 1048–1072. [Google Scholar] [CrossRef]
- Emwas, A.H. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods Mol. Biol. 2015, 1277, 161–193. [Google Scholar] [CrossRef]
- Hazard, D.; Fernandez, X.; Pinguet, J.; Chambon, C.; Letisse, F.; Portais, J.-C.; Wadih-Moussa, Z.; Rémignon, H.; Molette, C. Functional genomics of the muscle response to restraint and transport in chickens. J. Anim. Sci. 2011, 89, 2717–2730. [Google Scholar] [CrossRef]
- Tomonaga, S.; Okuyama, H.; Tachibana, T.; Makino, R. Effects of high ambient temperature on plasma metabolomic profiles in chicks. Anim. Sci. J. 2018, 89, 448–455. [Google Scholar] [CrossRef]
- Kang, D.R.; Shim, K.S. Proteomic analysis of the protective effect of early heat exposure against chronic heat stress in broilers. Animals 2020, 10, 2365. [Google Scholar] [CrossRef]
- Taylor, E.J.; Nott, H.M.; Earle, K.E. Dietary glycine: Its importance in growth and development of the budgerigar (Melopsittacus undulatus). J. Nutr. 1994, 124, 2555S–2558S. [Google Scholar] [CrossRef] [PubMed]
- Niknafs, S.; Fortes, M.R.S.; Cho, S.; Black, J.L.; Roura, E. Alanine-specific appetite in slow growing chickens is associated with impaired glucose transport and TCA cycle. BMC Genom. 2022, 23, 393. [Google Scholar] [CrossRef] [PubMed]
- Ngo, A.; Coon, C.N.; Beecher, G.R. Dietary glycine requirements for growth and cellular development in chicks. J. Nutr. 1977, 107, 1800–1808. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino Acids Biochemistry and Nutrition, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Knabe, D.A.; Li, P.; Li, X.; Mcknight, J.R.; Satterfield, M.C.; et al. Proline and hydroxyproline metabolism: Implications for animal and human nutrition. Amino Acids 2011, 40, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef]
- Klandorf, H.; Probert, I.L.; Iqbal, M. In the defence against hyperglycaemia: An avian strategy. Worlds Poult. Sci. J. 1999, 55, 251–268. [Google Scholar] [CrossRef]
- Stinefelt, B.; Leonard, S.S.; Blemings, K.P.; Shi, X.L.; Klandorf, H. Free radical scavenging, DNA protection, and inhibition of lipid peroxidation mediated by uric acid. Ann. Clin. Lab. Sci. 2005, 35, 37–45. [Google Scholar]
- Simoyi, M.F.; Falkenstein, E.; Van Dyke, K.; Blemings, K.P.; Klandorf, H. Allantoin, the oxidation product of uric acid is present in chicken and turkey plasma. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 135, 325–335. [Google Scholar] [CrossRef]
- Salway, J.G. The Krebs uric acid cycle: A forgotten Krebs cycle. Trends Biochem. Sci. 2018, 43, 847–849. [Google Scholar] [CrossRef]
- He, W.; Li, P.; Wu, G. Amino acid nutrition and metabolism in chickens. In Advances in Experimental Medicine and Biology; Springer International Publishing: Berlin/Heidelberg, Germany, 2021; pp. 109–131. [Google Scholar] [CrossRef]
- Emmanuel, B.; Howard, B.R. Endogenous uric acid and urea metabolism in the chicken. Br. Poult. Sci. 1978, 19, 295–301. [Google Scholar] [CrossRef]
- Malheiros, R.D.; Moraes, V.M.B.; Collin, A.; Decuypere, E.; Buyse, J. Free diet selection by broilers as influenced by dietary macronutrient ratio and corticosterone supplementation. 1. Diet selection, organ weights, and plasma metabolites. Poult. Sci. 2003, 82, 123–131. [Google Scholar] [CrossRef]
- Phang, J.M.; Liu, W.E.I.; Zabirnyk, O. Proline metabolism and microenvironmental stress. Ann. Rev. Nutr. 2010, 30, 441–463. [Google Scholar] [CrossRef]
- Phang, J.M.; Pandhare, J.; Liu, Y. The metabolism of proline as microenvironmental stress substrate. J. Nutr. 2008, 138, 2008S–2015S. [Google Scholar] [CrossRef]
- Liu, S.Y.; Macelline, S.P.; Chrystal, P.V.; Selle, P.H. Progress towards reduced-crude protein diets for broiler chickens and sustainable chicken-meat production. J. Anim. Sci. Biotechnol. 2021, 12, 20. [Google Scholar] [CrossRef]
- Greenhalgh, S.; Chrystal, P.V.; Selle, P.H.; Liu, S.Y. Reduced-crude protein diets in chicken-meat production: Justification for an imperative. Worlds Poult. Sci. J. 2020, 76, 537–548. [Google Scholar] [CrossRef]
- Xue, G.-D.; Wu, S.-B.; Choct, M.; Swick, R.A. The role of supplemental glycine in establishing a subclinical necrotic enteritis challenge model in broiler chickens. Anim. Nutr. 2017, 3, 266–270. [Google Scholar] [CrossRef]
- Rahimi, J.; Zendehdel, M.; Khodadadi, M. Mediatory role of the dopaminergic system through D1 receptor on glycine-induced hypophagia in neonatal broiler-type chickens. Amino Acids 2021, 53, 461–470. [Google Scholar] [CrossRef]
- De Koning, D.-J.; Dominguez-Gasca, N.; Fleming, R.H.; Gill, A.; Kurian, D.; Law, A.; McCormack, H.A.; Morrice, D.; Sanchez-Rodriguez, E.; Rodriguez-Navarro, A.B.; et al. An eQTL in the cystathionine beta synthase gene is linked to osteoporosis in laying hens. Genet. Sel. Evol. 2020, 52, 13. [Google Scholar] [CrossRef]
- Maidin, M.B.M.; McCormack, H.A.; Wilson, P.W.; Caughey, S.D.; Whenham, N.; Dunn, I.C. Dietary betaine reduces plasma homocysteine concentrations and improves bone strength in laying hens. Br. Poult. Sci. 2021, 62, 573–578. [Google Scholar] [CrossRef]
- Watford, M. Gluconeogenesis in the chicken: Regulation of phosphoenolpyruvate carboxykinase gene expression. Fed. Proc. 1985, 44, 2469–2474. [Google Scholar] [PubMed]
- Zhang, S.-S.; Su, H.-G.; Zhou, Y.; Li, X.-M.; Feng, J.-H.; Zhang, M.-H. Effects of sustained cold and heat stress on energy intake, growth and mitochondrial function of broiler chickens. J. Integr. Agric. 2016, 15, 2336–2342. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, L.; Li, J.; Xing, T.; Jiang, Y.; Gao, F. Heat stress alters muscle protein and amino acid metabolism and accelerates liver gluconeogenesis for energy supply in broilers. Poult. Sci. 2021, 100, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Garriga, C.; Hunter, R.R.; Amat, C.; Planas, J.M.; Mitchell, M.A.; Moreto, M. Heat stress increases apical glucose transport in the chicken jejunum. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R195–R201. [Google Scholar] [CrossRef]
- Garcia-Palmer, F.J.; Pons, A.; Alemany, M.; Palou, A. Patterns of amino acid enzyme in domestic fowl breast and leg muscle during development. Comp. Biochem. Physiol. B Comp. Biochem. 1985, 82, 143–146. [Google Scholar] [CrossRef]
- Chen, R.; Wen, C.; Gu, Y.; Wang, C.; Chen, Y.; Zhuang, S.; Yanmin, Z. Dietary betaine supplementation improves meat quality of transported broilers through altering muscle anaerobic glycolysis and antioxidant capacity. J. Sci. Food Agric. 2020, 100, 2656–2663. [Google Scholar] [CrossRef]
- Alanazi, A.M.; Mostafa, G.A.E.; Al-Badr, A.A. Glutathione. In Profiles of Drug Substances, Excipients, and Related Methodology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 40, pp. 43–158. [Google Scholar]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef]
- Castro, F.L.d.S.; Kim, W.K. Secondary functions of arginine and sulfur amino acids in poultry health: Review. Animals 2020, 10, 2106. [Google Scholar] [CrossRef]
- Kidd, M.T. Nutritional considerations concerning threonine in broilers. Worlds Poult. Sci. J. 2000, 56, 139–151. [Google Scholar] [CrossRef]
- Khan, A.; Nawaz, H.; Zahoor, I. Effect of different levels of digestible threonine on growth performance of broiler chicks. J. Anim. Plant Sci. 2006, 16, 8–11. [Google Scholar]
- Kheiri, F.; Alibeyghi, M. Effect of different levels of lysine and threonine on carcass characteristics, intestinal microflora and growth performance of broiler chicks. Ital. J. Anim. Sci. 2017, 16, 580–587. [Google Scholar] [CrossRef]
- Yang, J.; Liu, L.; Sheikhahmadi, A.; Wang, Y.; Li, C.; Jiao, H.; Lin, H.; Song, Z. Effects of corticosterone and dietary energy on immune function of broiler chickens. PLoS ONE 2015, 10, e0119750. [Google Scholar] [CrossRef]
- Boehm, G.; Cervantes, H.; Georgi, G.; Jelinek, J.; Sawatzki, G.; Wermuth, B.; Colombo, J.-P. Effect of increasing dietary threonine intakes on amino acid metabolism of the central nervous system and peripheral tissues in growing rats. Pediatr. Res. 1998, 44, 900–906. [Google Scholar] [CrossRef]
- Smulders, T.V. The avian hippocampal formation and the stress response. Brain Behav. Evol. 2017, 90, 81–91. [Google Scholar] [CrossRef]
- Yang, M.; Chen, R.; Song, Y.D.; Zhou, Y.M.; Liu, Q.; Zhuang, S. Effects of dietary betaine supplementation on growth performance, meat quality, muscle fatty acid composition and antioxidant ability in slow-growing broiler chickens. Br. Poult. Sci. 2022, 63, 351–359. [Google Scholar] [CrossRef]
- Nasiroleslami, M.; Torki, M.; Saki, A.A.; Abdolmohammadi, A.R. Effects of dietary guanidinoacetic acid and betaine supplementation on performance, blood biochemical parameters and antioxidant status of broilers subjected to cold stress. J. Appl. Anim. Res. 2018, 46, 1016–1022. [Google Scholar] [CrossRef]
- Shakeri, M.; Cottrell, J.; Wilkinson, S.; Ringuet, M.; Furness, J.; Dunshea, F. Betaine and antioxidants improve growth performance, breast muscle development and ameliorate thermoregulatory responses to cyclic heat exposure in broiler chickens. Animals 2018, 8, 162. [Google Scholar] [CrossRef]
- Wen, C.; Chen, R.; Chen, Y.; Ding, L.; Wang, T.; Zhou, Y. Betaine improves growth performance, liver health, antioxidant status, breast meat yield, and quality in broilers fed a mold-contaminated corn-based diet. Anim. Nutr. 2021, 7, 661–666. [Google Scholar] [CrossRef]
- Wen, C.; Chen, Y.; Leng, Z.; Ding, L.; Wang, T.; Zhou, Y. Dietary betaine improves meat quality and oxidative status of broilers under heat stress. J. Sci. Food Agric. 2019, 99, 620–623. [Google Scholar] [CrossRef]
- Chowdhury, V.S. Heat stress biomarker amino acids and neuropeptide afford thermotolerance in chicks. J. Poult. Sci. 2019, 56, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, V.S.; Han, G.; Eltahan, H.M.; Haraguchi, S.; Gilbert, E.R.; Cline, M.A.; Cockrem, J.F.; Bungo, T.; Furuse, M. Potential role of amino acids in the adaptation of chicks and market-age broilers to heat stress. Front. Vet. Sci. 2021, 7, 610541. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.A.K.; Kikusato, M.; Maekawa, T.; Shirakawa, H.; Toyomizu, M. Metabolic characteristics and oxidative damage to skeletal muscle in broiler chickens exposed to chronic heat stress. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 155, 401–406. [Google Scholar] [CrossRef]
- Eltahan, H.M.; Bahry, M.A.; Yang, H.; Han, G.; Nguyen, L.T.N.; Ikeda, H.; Ali, M.N.; Amber, K.A.; Furuse, M.; Chowdhury, V.S. Central NPY-Y5 sub-receptor partially functions as a mediator of NPY-induced hypothermia and affords thermotolerance in heat-exposed fasted chicks. Physiol. Rep. 2017, 5, e13511. [Google Scholar] [CrossRef]
- Ito, K.; Bahry, M.A.; Hui, Y.; Furuse, M.; Chowdhury, V.S. Acute heat stress up-regulates neuropeptide Y precursor mRNA expression and alters brain and plasma concentrations of free amino acids in chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 187, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Yang, H.; Bungo, T.; Ikeda, H.; Wang, Y.; Nguyen, L.T.N.; Eltahan, H.M.; Furuse, M.; Chowdhury, V.S. In ovo L-leucine administration stimulates lipid metabolisms in heat-exposed male, but not female, chicks to afford thermotolerance. J. Therm. Biol. 2018, 71, 74–82. [Google Scholar] [CrossRef]
- Chowdhury, V.S.; Tomonaga, S.; Nishimura, S.; Tabata, S.; Furuse, M. Physiological and behavioral responses of young chicks to high ambient temperature. J. Poult. Sci. 2012, 49, 212–218. [Google Scholar] [CrossRef]
- Kamboh, A.A.; Hang, S.Q.; Bakhetgul, M.; Zhu, W.-Y. Effects of genistein and hesperidin on biomarkers of heat stress in broilers under persistent summer stress. Poult. Sci. 2013, 92, 2411–2418. [Google Scholar] [CrossRef]
- Bartov, I. Effects of dietary protein concentration and corticosterone injections on energy and nitrogen balances and fat deposition in broiler chicks. Br. Poult. Sci. 1985, 26, 311–324. [Google Scholar] [CrossRef]
- Hayashi, K.; Nagai, Y.; Ohstuka, A.; Tomita, Y. Effects of dietary corticosterone and trilostane on growth and skeletal muscle protein turnover in broiler cockerels. Br. Poult. Sci. 1994, 35, 789–798. [Google Scholar] [CrossRef]
- Fairchild, B.D.; Ritz, C.W. Poultry Drinking Water Primer; University of Georgia: Athens, GA, USA, 2015; Available online: https://extension.uga.edu/publications/detail.html?number=B1301 (accessed on 16 December 2022).
- Gerbens-Leenes, P.W.; Mekonnen, M.M.; Hoekstra, A.Y. The water footprint of poultry, pork and beef: A comparative study in different countries and production systems. Water Res. Ind. 2013, 1–2, 25–36. [Google Scholar] [CrossRef]
- Mekonnen, M.M.; Hoekstra, A.Y. A global assessment of the water footprint of farm animal products. Ecosystems 2012, 15, 401–415. [Google Scholar] [CrossRef]
- Rensel, M.A.; Schlinger, B.A. Determinants and significance of corticosterone regulation in the songbird brain. Gen. Comp. Endocrinol. 2016, 227, 136–142. [Google Scholar] [CrossRef]
- Breuner, C.W.; Orchinik, M. Seasonal regulation of membrane and intracellular corticosteroid receptors in the house sparrow brain. J. Neuroendocrinol. 2001, 13, 412–420. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Song, G.; Lei, F.; Li, D.; Qu, Y. Avian corticosteroid-binding globulin: Biological function and regulatory mechanisms in physiological stress responses. Front. Zool. 2021, 18, 22. [Google Scholar] [CrossRef]
- Rosner, W. The functions of corticsteroid-binding globuline and sex hormone-binding globulin: Recent advances. Endocr. Rev. 1990, 11, 80–91. [Google Scholar] [CrossRef]
- Breuner, C.W.; Orchinik, M. Plasma binding proteins as mediators of corticosteroid action in vertebrates. J. Endocrinol. 2002, 175, 99–112. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, C.L.J.; Zaytsoff, S.J.M.; Iwaniuk, A.N.; Metz, G.A.S.; Montina, T.; Inglis, G.D. Comparative Analysis of the Temporal Impacts of Corticosterone and Simulated Production Stressors on the Metabolome of Broiler Chickens. Metabolites 2023, 13, 144. https://doi.org/10.3390/metabo13020144
Brown CLJ, Zaytsoff SJM, Iwaniuk AN, Metz GAS, Montina T, Inglis GD. Comparative Analysis of the Temporal Impacts of Corticosterone and Simulated Production Stressors on the Metabolome of Broiler Chickens. Metabolites. 2023; 13(2):144. https://doi.org/10.3390/metabo13020144
Chicago/Turabian StyleBrown, Catherine L. J., Sarah J. M. Zaytsoff, Andrew N. Iwaniuk, Gerlinde A. S. Metz, Tony Montina, and G. Douglas Inglis. 2023. "Comparative Analysis of the Temporal Impacts of Corticosterone and Simulated Production Stressors on the Metabolome of Broiler Chickens" Metabolites 13, no. 2: 144. https://doi.org/10.3390/metabo13020144
APA StyleBrown, C. L. J., Zaytsoff, S. J. M., Iwaniuk, A. N., Metz, G. A. S., Montina, T., & Inglis, G. D. (2023). Comparative Analysis of the Temporal Impacts of Corticosterone and Simulated Production Stressors on the Metabolome of Broiler Chickens. Metabolites, 13(2), 144. https://doi.org/10.3390/metabo13020144