

Citronellal as a Promising Candidate for Alzheimer’s Disease Treatment: A Comprehensive Study on In Silico and In Vivo Anti-Acetylcholine Esterase Activity

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drug Likeliness and ADME/T Analysis
2.2. Molecular Docking
2.3. Molecular Dynamics
2.4. Chemicals and Software
2.5. Animals and Treatment
2.6. Morris Water Maze (MWM) Test
2.7. Cook’s Pole Climbing (CPC) Test
2.8. Collection of Brain Samples
2.9. Brain Chemical Parameters
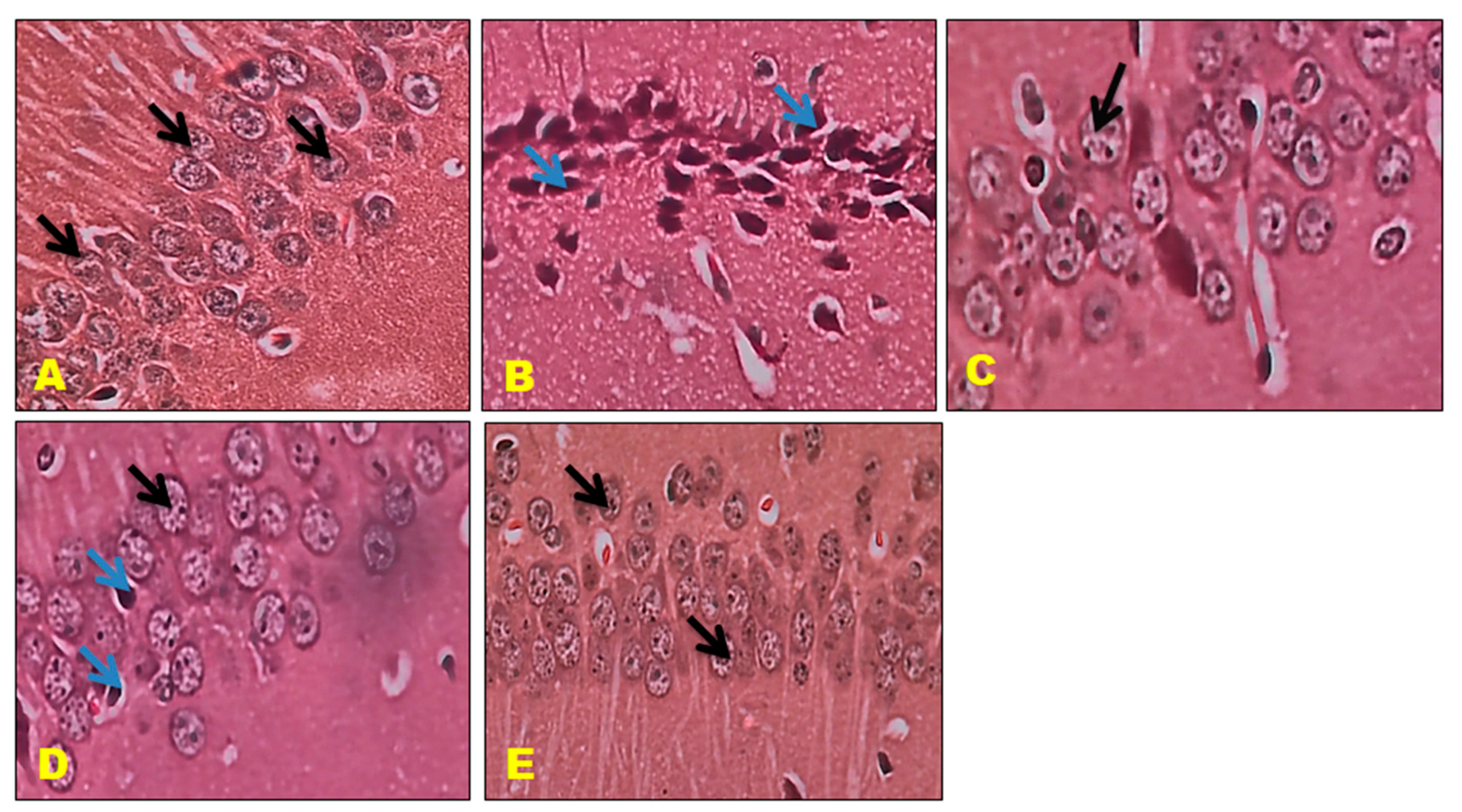
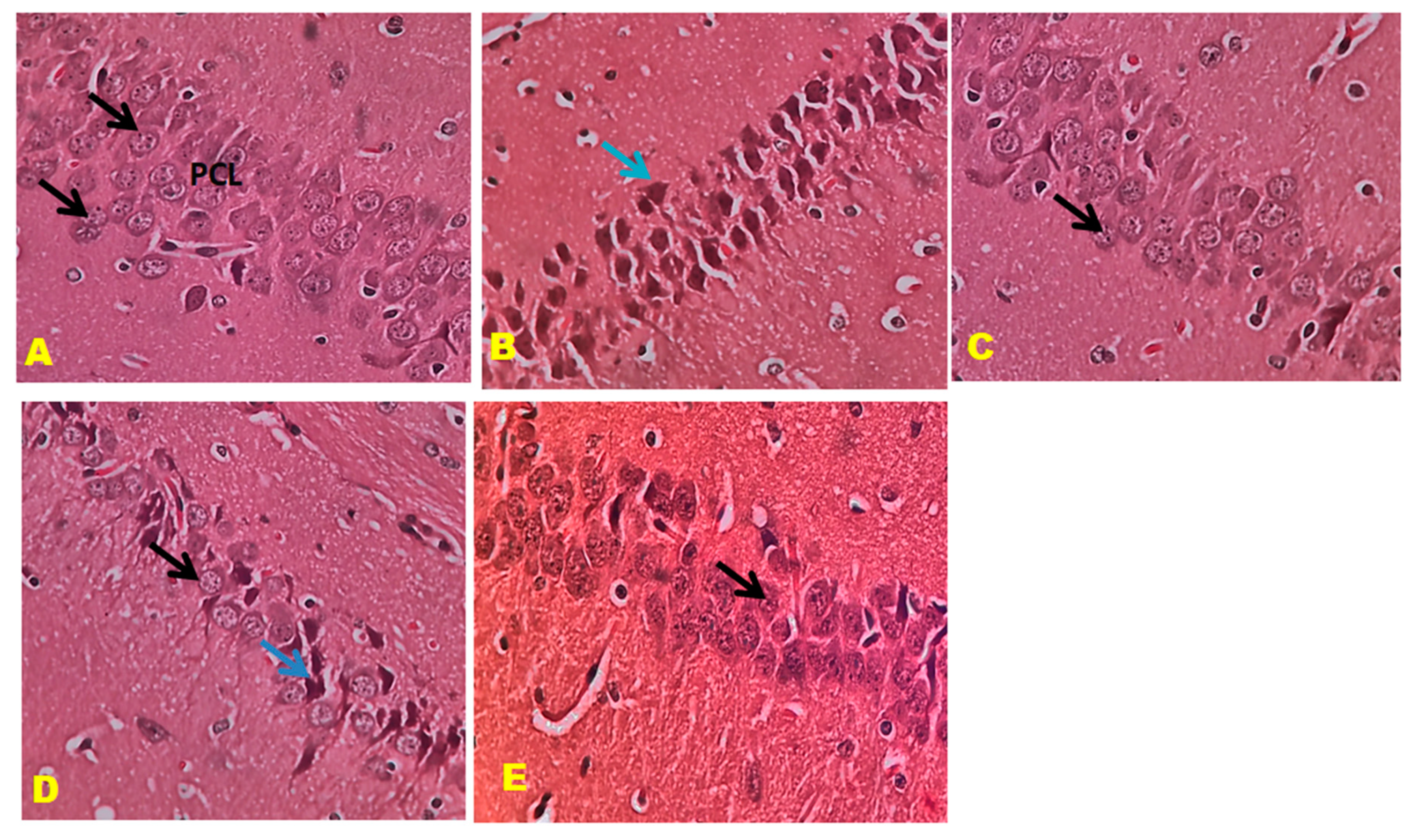
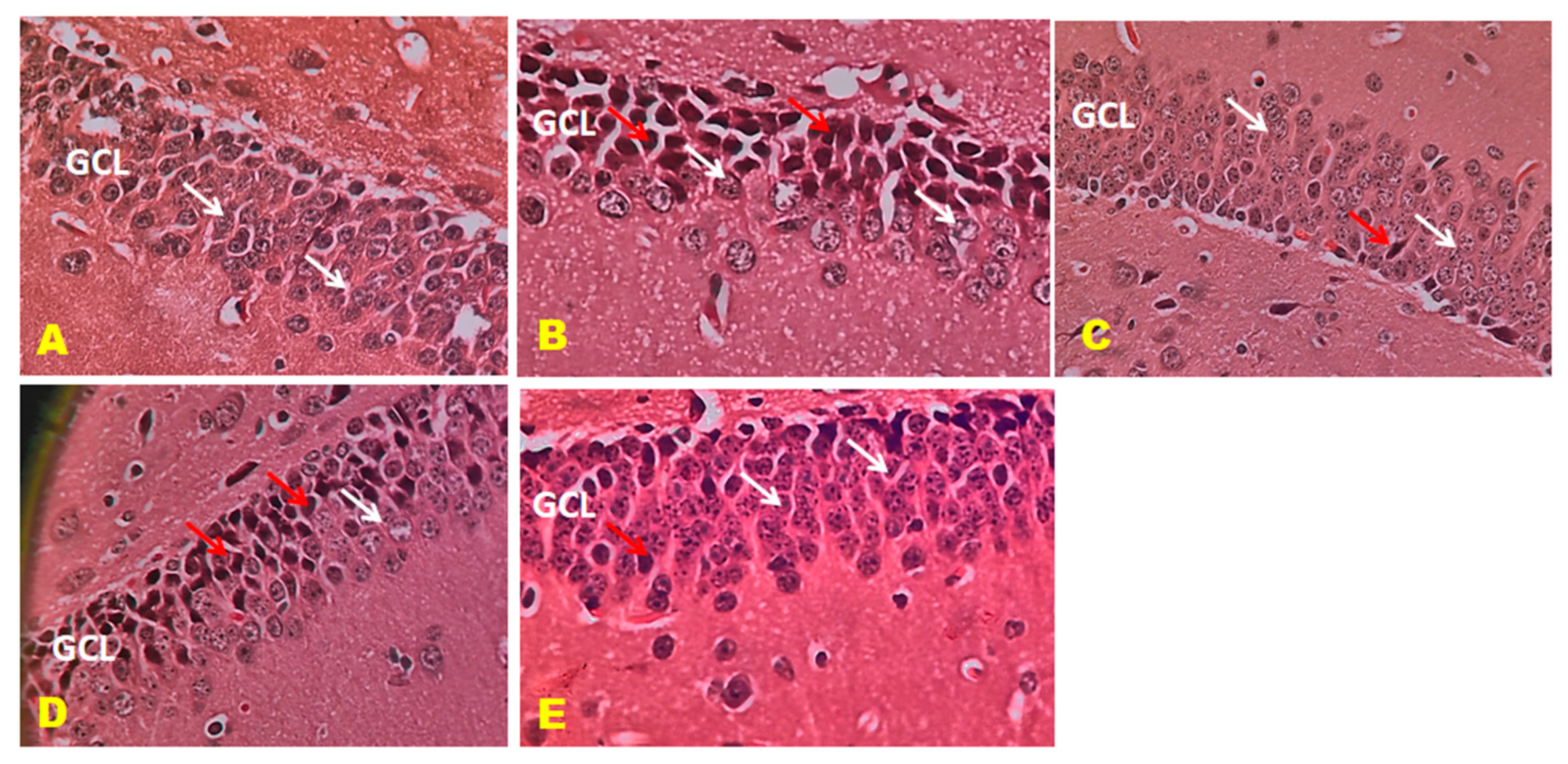
2.10. Histopathological Examination
2.11. Statistical Analysis
3. Results
3.1. Drug Likeliness and ADMET Analysis
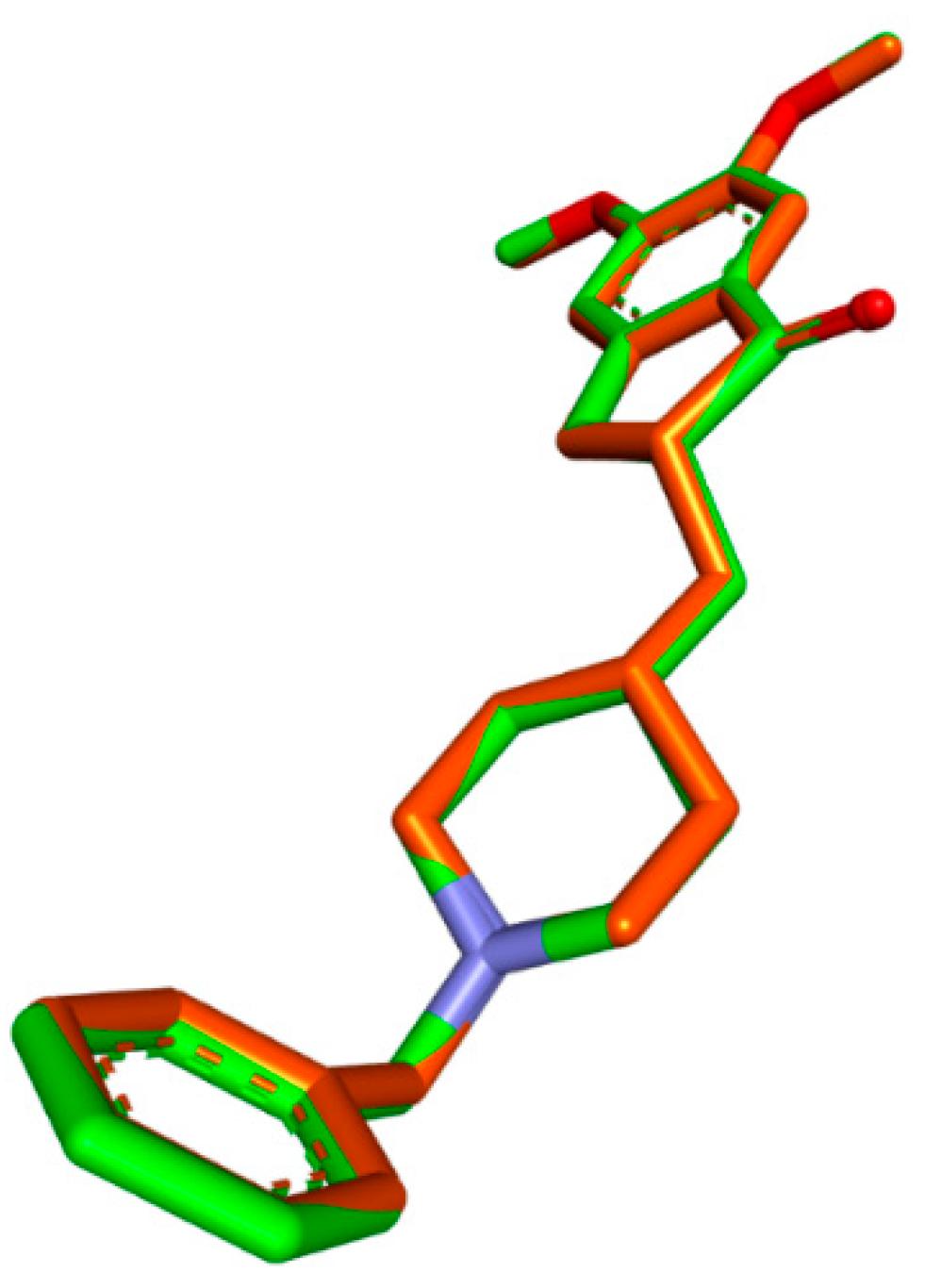
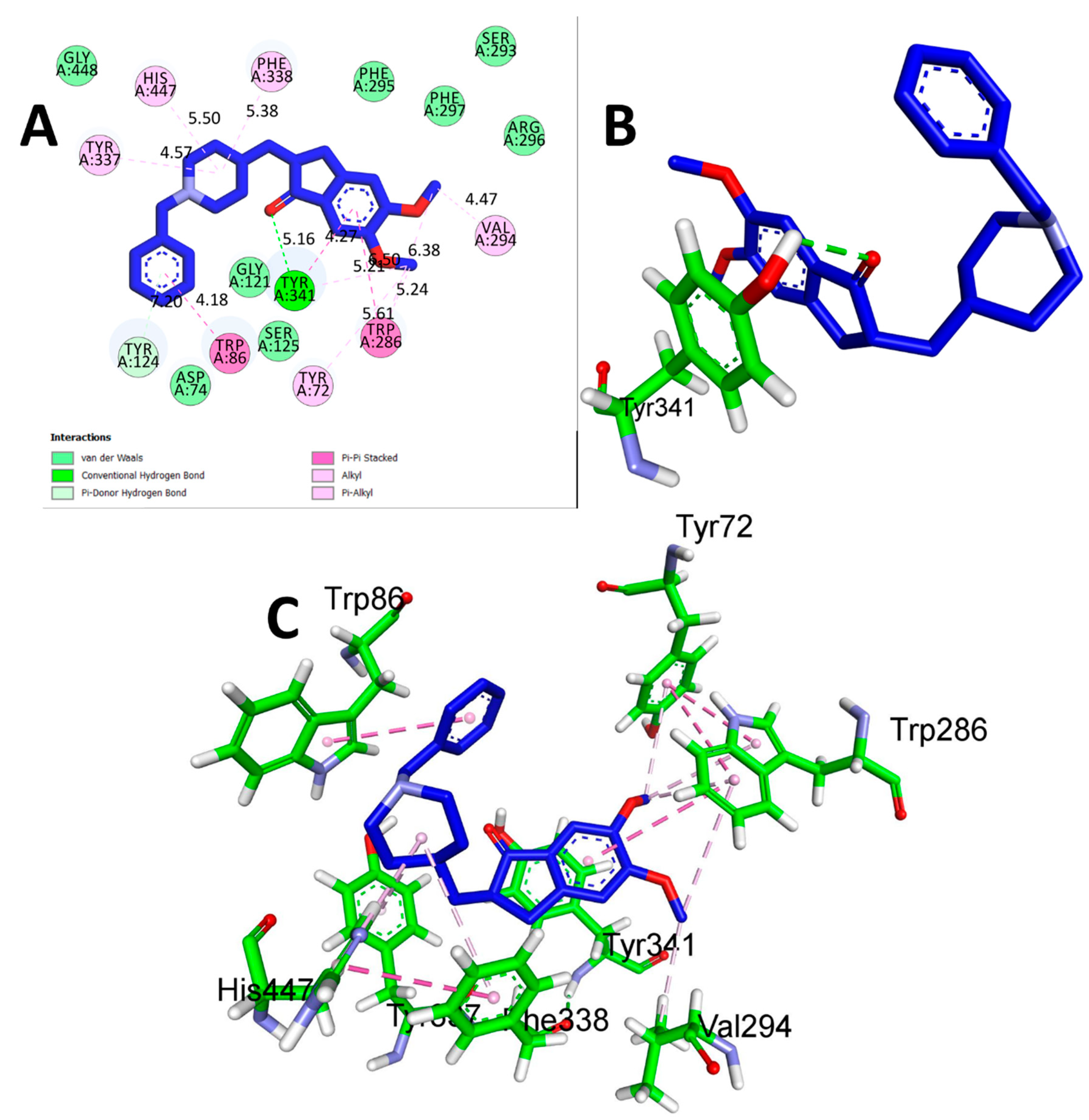
3.2. Molecular Docking
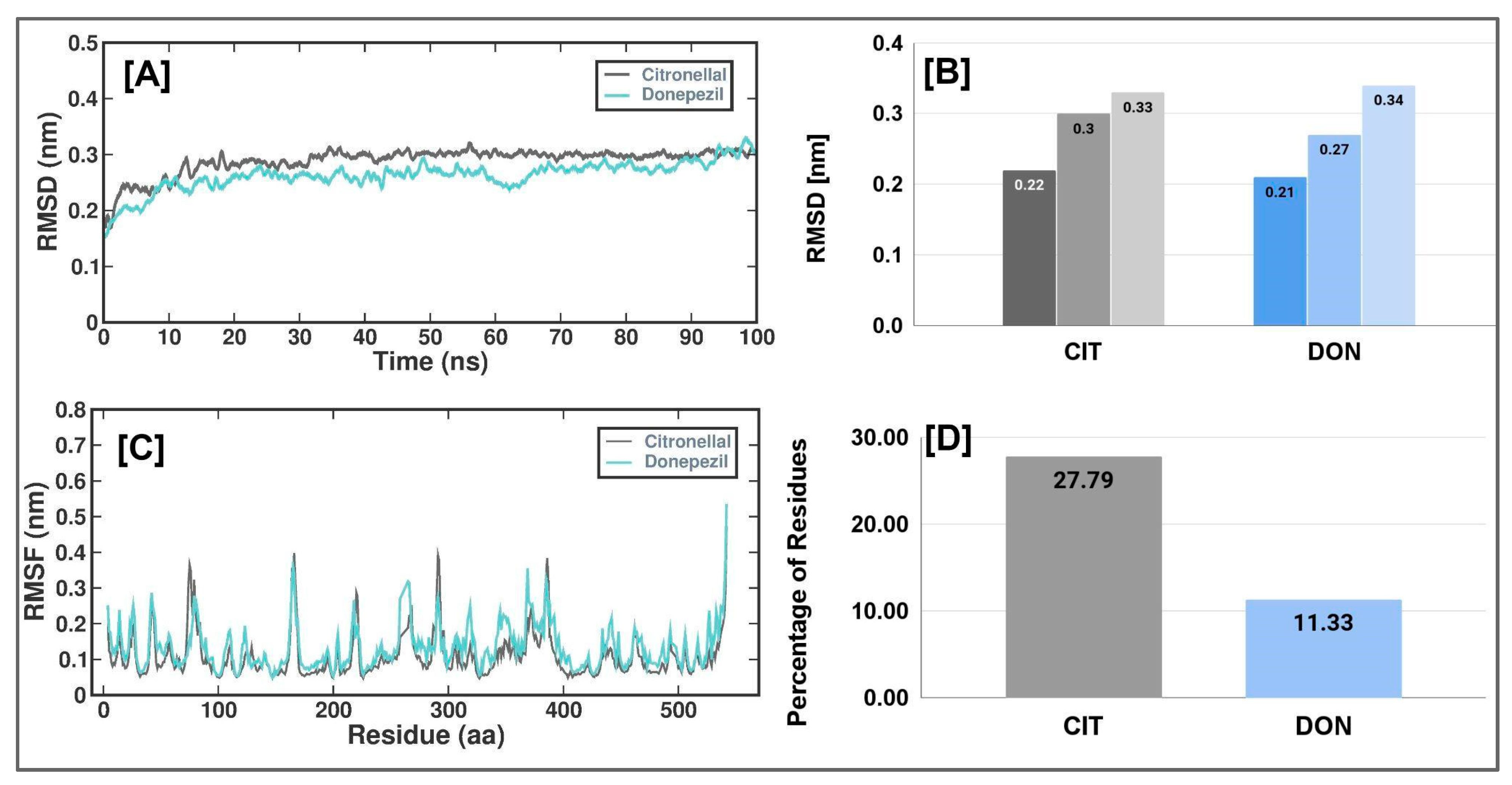
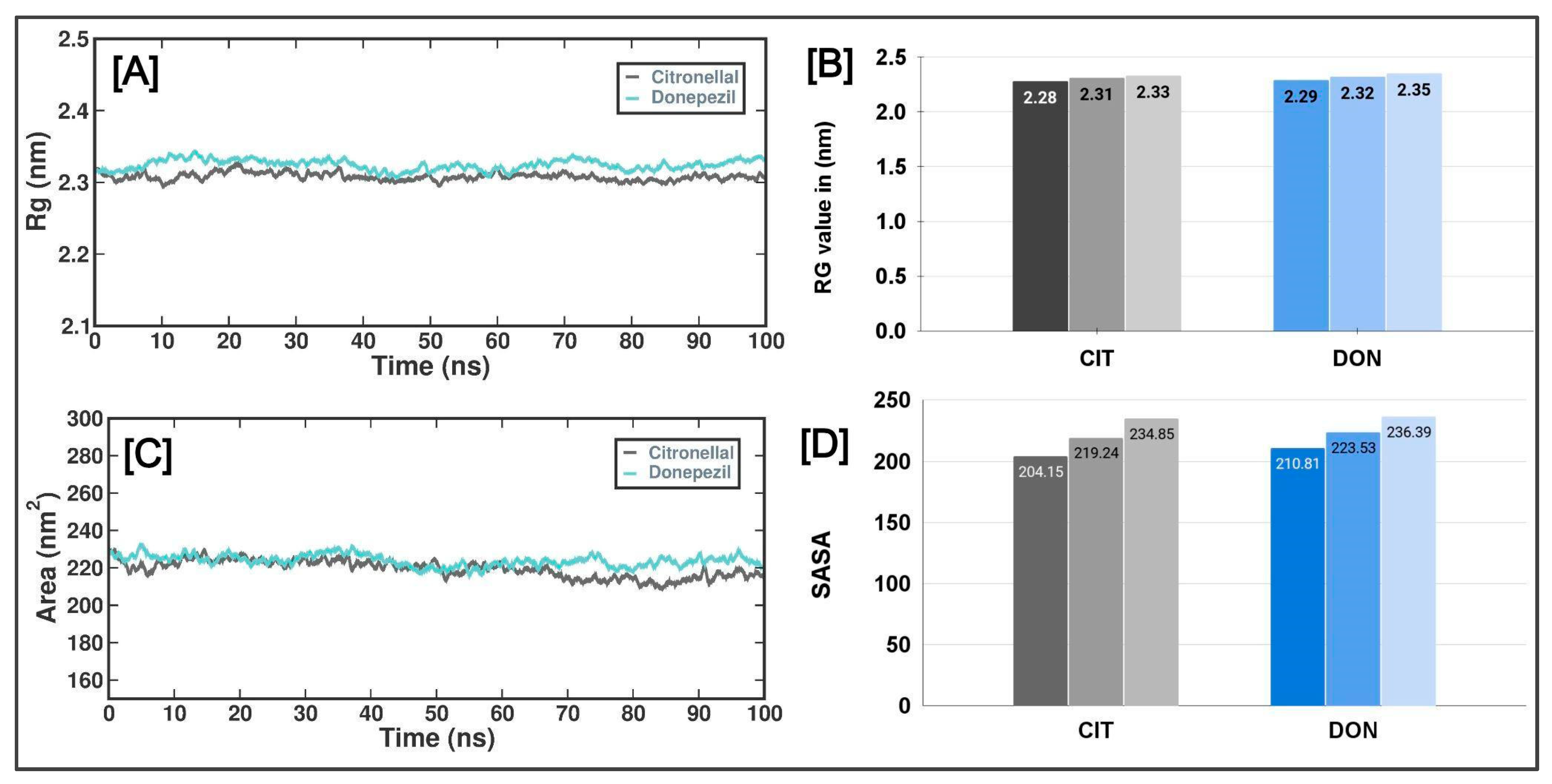
3.3. Molecular Simulation Studies
Induction of AD
3.4. Behaviours Analysis
3.4.1. MWM Test
3.4.2. CPC Test
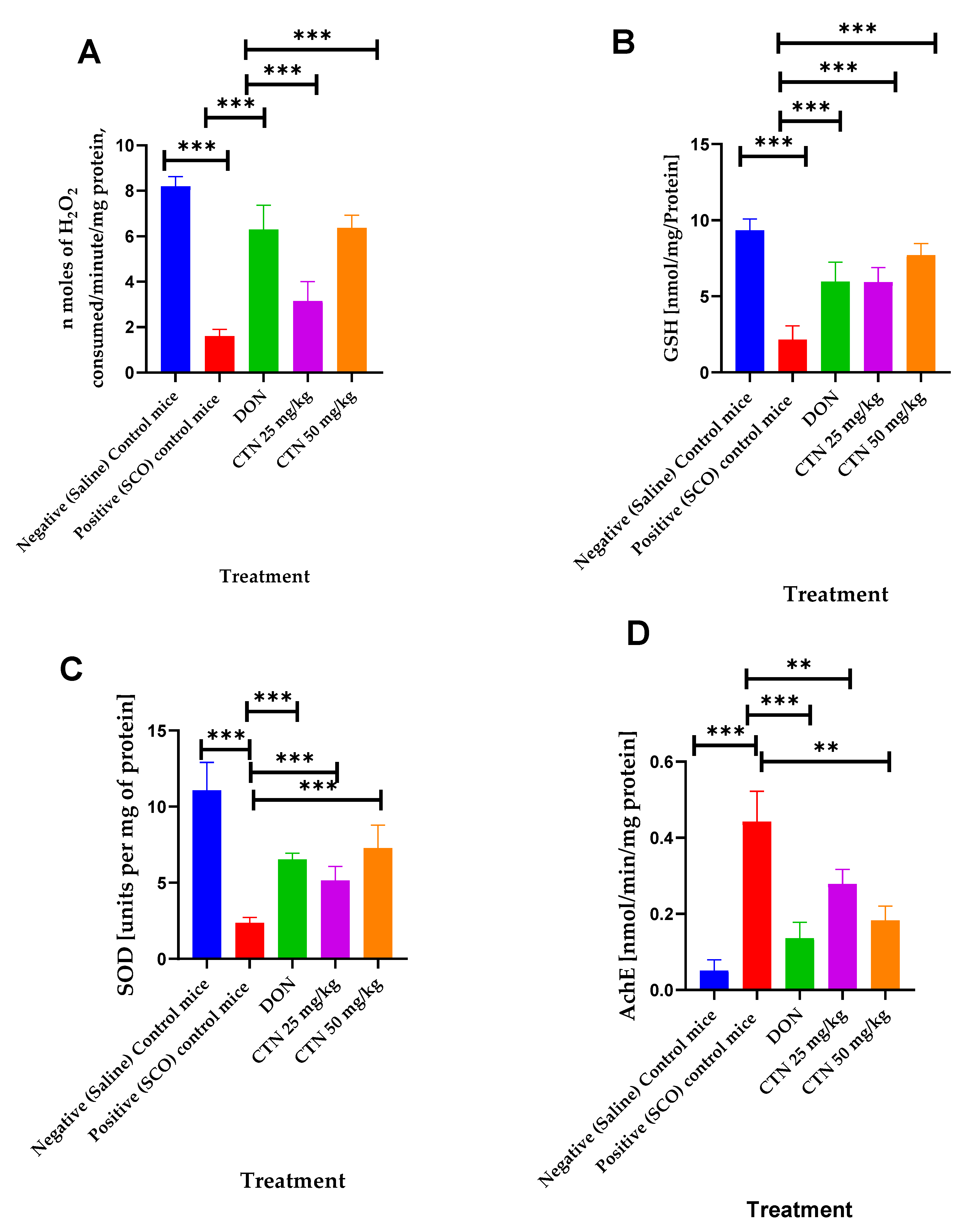
3.5. Brain Biochemical Analysis
3.6. Histopathology Studies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogbodo, J.O.; Agbo, C.P.; Njoku, U.O.; Ogugofor, M.O.; Egba, S.I.; Ihim, S.A.; Echezona, A.C.; Brendan, K.C.; Upaganlawar, A.B.; Upasani, C.D. Alzheimer’s Disease: Pathogenesis and Therapeutic Interventions. Curr. Aging Sci. 2022, 15, 2–25. [Google Scholar] [CrossRef] [PubMed]
- World Alzheimer Report 2019. Available online: https://www.alzint.org/resource/world-alzheimer-report-2019/ (accessed on 11 September 2023).
- Savelieff, M.G.; Lee, S.; Liu, Y.; Lim, M.H. Untangling Amyloid-β, Tau, and Metals in Alzheimer’s Disease. ACS Chem. Biol. 2013, 8, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Trippier, P.C.; Jansen Labby, K.; Hawker, D.D.; Mataka, J.J.; Silverman, R.B. Target- and Mechanism-Based Therapeutics for Neurodegenerative Diseases: Strength in Numbers. J. Med. Chem. 2013, 56, 3121–3147. [Google Scholar] [CrossRef] [PubMed]
- Agis-Torres, A.; Sollhuber, M.; Fernandez, M.; Sanchez-Montero, J.M. Multi-Target-Directed Ligands and other Therapeutic Strategies in the Search of a Real Solution for Alzheimer’s Disease. Curr. Neuropharmacol. 2014, 12, 2–36. [Google Scholar] [CrossRef] [PubMed]
- Blaikie, L.; Kay, G.; Lin, P.K.T. Current and emerging therapeutic targets of alzheimer’s disease for the design of multi-target directed ligands. MedChemComm 2019, 10, 2052–2072. [Google Scholar] [CrossRef] [PubMed]
- Petralia, M.C.; Battaglia, G.; Bruno, V.; Pennisi, M.; Mangano, K.; Lombardo, S.D.; Fagone, P.; Cavalli, E.; Saraceno, A.; Nicoletti, F.; et al. The Role of Macrophage Migration Inhibitory Factor in Alzheimer′s Disease: Conventionally Pathogenetic or Unconventionally Protective? Molecules 2020, 25, 291. [Google Scholar] [CrossRef]
- Petralia, M.C.; Mazzon, E.; Fagone, P.; Basile, M.S.; Lenzo, V.; Quattropani, M.C.; Di Nuovo, S.; Bendtzen, K.; Nicoletti, F. The cytokine network in the pathogenesis of major depressive disorder. Close to translation? Autoimmun. Rev. 2020, 19, 102504. [Google Scholar] [CrossRef]
- Scarpini, E.; Scheltens, P.; Feldman, H. Treatment of Alzheimer’s disease: Current status and new perspectives. Lancet Neurol. 2003, 2, 539–547. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Dinamarca, M.C.; Alvarez, A. Amyloid-cholinesterase interactions. Implications for Alzheimer’s disease. FEBS J. 2008, 275, 625–632. [Google Scholar] [CrossRef]
- Pohanka, M. Acetylcholinesterase inhibitors: A patent review (2008–present). Expert Opin. Ther. Pat. 2012, 22, 871–886. [Google Scholar] [CrossRef]
- Kelly, C.A.; Harvey, R.J.; Cayton, H. Drug treatments for Alzheimer’s disease. BMJ 1997, 314, 693–694. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, P.J. Cholinergic therapy in dementia. Acta Neurol. Scand. 1993, 88, 42–45. [Google Scholar] [CrossRef]
- Gottwald, M.D.; Rozanski, R.I. Rivastigmine, a brain-region selective acetylcholinesterase inhibitor for treating Alzheimer’s disease: Review and current status. Expert. Opin. Investig. Drugs 1999, 8, 1673–1682. [Google Scholar] [CrossRef]
- Scott, L.J.; Goa, K.L. Galantamine. Drugs 2000, 60, 1095–1122. [Google Scholar] [CrossRef] [PubMed]
- Crismon, M.L. Tacrine: First drug approved for Alzheimer’s disease. Ann. Pharmacother. 1994, 28, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zhang, J.; Wang, Y.; Cui, K.; Cao, Y.; Wang, L.; Wu, Y. Linarin improves the dyskinesia recovery in Alzheimer’s disease zebrafish by inhibiting the acetylcholinesterase activity. Life Sci. 2019, 222, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.P.; Or, T.C.; Ip, N.Y. Plant alkaloids as drug leads for Alzheimer’s disease. Neurochem. Int. 2015, 89, 260–270. [Google Scholar] [CrossRef]
- Dash, U.C.; Kanhar, S.; Dixit, A.; Dandapat, J.; Sahoo, A.K. Isolation, identification, and quantification of Pentylcurcumene from Geophila repens: A new class of cholinesterase inhibitor for Alzheimer’s disease. Bioorg. Chem. 2019, 88, 102947. [Google Scholar] [CrossRef]
- Shrivastava, S.K.; Sinha, S.K.; Srivastava, P.; Tripathi, P.N.; Sharma, P.; Tripathi, M.K.; Tripathi, A.; Choubey, P.K.; Waiker, D.K.; Aggarwal, L.M.; et al. Design and development of novel p-aminobenzoic acid derivatives as potential cholinesterase inhibitors for the treatment of Alzheimer’s disease. Bioorg. Chem. 2019, 82, 211–223. [Google Scholar] [CrossRef]
- Yu, S.E.; Mwesige, B.; Yi, Y.S.; Yoo, B.C. Ginsenosides: The need to move forward from bench to clinical trials. J. Ginseng Res. 2019, 43, 361–367. [Google Scholar] [CrossRef]
- Saklani, A.; Kutty, S.K. Plant-derived compounds in clinical trials. Drug Discov. Today 2008, 13, 161–171. [Google Scholar] [CrossRef]
- Kaur, H.; Bhardwaj, U.; Kaur, R. Cymbopogon nardus essential oil: A comprehensive review on its chemistry and bioactivity. J. Essent. Oil Res. 2021, 33, 205–220. [Google Scholar] [CrossRef]
- Trindade, L.A.; Cordeiro, L.V.; de Figuerêdo Silva, D.; Figueiredo, P.T.R.; de Pontes, M.L.C.; de Oliveira Lima, E.; de Albuquerque Tavares Carvalho, A. The antifungal and antibiofilm activity of Cymbopogon nardus essential oil and citronellal on clinical strains of Candida albicans. Braz. J. Microbiol. 2022, 53, 1231–1240. [Google Scholar] [CrossRef]
- Victoria, F.N.; Anversa, R.; Penteado, F.; Castro, M.; Lenardão, E.J.; Savegnago, L. Antioxidant and antidepressant-like activities of semi-synthetic α-phenylseleno citronellal. Eur. J. Pharmacol. 2014, 742, 131–138. [Google Scholar] [CrossRef]
- Waliwitiya, R.; Isman, M.B.; Vernon, R.S.; Riseman, A. Insecticidal Activity of Selected Monoterpenoids and Rosemary Oil to Agriotes obscurus (Coleoptera: Elateridae). J. Econ. Entomol. 2005, 98, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.S.; Guimares, A.G.; Santana, M.F.; Siqueira, R.S.; De Lima, A.D.C.B.; Dias, A.S.; Santos, M.R.V.; Onofre, A.S.C.; Quintans, J.S.S.; De Sousa, D.P.; et al. Anti-inflammatory and redox-protective activities of citronellal. Biol. Res. 2011, 44, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Melo, M.S.; Sena, L.C.S.; Barreto, F.J.N.; Bonjardim, L.R.; Almeida, J.R.G.S.; Lima, J.T.; De Sousa, D.P.; Quintans-Júnior, L.J. Antinociceptive effect of citronellal in mice. Pharm. Biol. 2010, 48, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Stojanović, N.M.; Mladenović, M.Z.; Randjelović, P.J.; Radulović, N.S. The potential of lemon balm (Melissa officinalis L.) essential oil as an anti-anxiety agent—Is the citronellal the activity carrier? J. Ethnopharmacol. 2023, 314, 116661. [Google Scholar] [CrossRef]
- Lu, J.-X.; Qiu, Y.; Guo, L.-J.; Song, P.; Xu, J.; Wan, G.-R.; Wang, S.-X.; Yin, Y.-L.; Li, P. Potential Therapeutic Effect of Citronellal on Diabetic Cardiomyopathy in Experimental Rats. Evid. Based Complement. Altern. Med. 2021, 2021, 9987531. [Google Scholar] [CrossRef]
- Maurya, A.K.; Aggarwal, G.; Vashisath, S.; Kumar, V.; Agnihotri, V.K. Chemodiversity and α-Glucosidase Activity of Eucalyptus Species from Northwestern Himalaya, India. Chem. Biodivers. 2023, 20, e202300223. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.; Rudkowska, M.; Kasprzak-Drozd, K.; Oniszczuk, A.; Borowicz-Reutt, K. Activity of selected group of monoterpenes in alzheimer’s disease symptoms in experimental model studies—A non-systematic review. Int. J. Mol. Sci. 2021, 22, 7366. [Google Scholar] [CrossRef]
- Campora, M.; Canale, C.; Gatta, E.; Tasso, B.; Laurini, E.; Relini, A.; Pricl, S.; Catto, M.; Tonelli, M. Multitarget Biological Profiling of New Naphthoquinone and Anthraquinone-Based Derivatives for the Treatment of Alzheimer’s Disease. ACS Chem. Neurosci. 2021, 12, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Kongsune, P.; Hannongbua, S. The role of conserved QXG and binding affinity of S23G & S26G receptors on avian H5, swine H1 and human H1 of influenza A virus hemagglutinin. J. Mol. Graph. Model. 2018, 82, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.J.; Roy, A.; Jung, H.J.; Ali, M.Y.; Min, B.S.; Park, C.H.; Yokozawa, T.; Fan, T.P.; Choi, J.S.; Jung, H.A. BACE1 molecular docking and anti-Alzheimer’s disease activities of ginsenosides. J. Ethnopharmacol. 2016, 190, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Abbasi, M.A.; Aziz Ur, R.; Siddiqui, S.Z.; Hussain, G.; Shah, S.A.A.; Shahid, M.; Seo, S.Y. Exploration of synthetic multifunctional amides as new therapeutic agents for Alzheimer’s disease through enzyme inhibition, chemoinformatic properties, molecular docking and dynamic simulation insights. J. Theor. Biol. 2018, 458, 169–183. [Google Scholar] [CrossRef]
- Ntie-Kang, F.; Lifongo, L.L.; Mbah, J.A.; Owono Owono, L.C.; Megnassan, E.; Mbaze, L.M.; Judson, P.N.; Sippl, W.; Efange, S.M. In silico drug metabolism and pharmacokinetic profiles of natural products from medicinal plants in the Congo basin. Silico Pharmacol. 2013, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lou, C.; Sun, L.; Li, J.; Cai, Y.; Wang, Z.; Li, W.; Liu, G.; Tang, Y. admetSAR 2.0: Web-service for prediction and optimization of chemical ADMET properties. Bioinformatics 2019, 35, 1067–1069. [Google Scholar] [CrossRef]
- Killari, K.N.; Polimati, H.; Prasanth, D.; Singh, G.; Panda, S.P.; Vedula, G.S.; Tatipamula, V.B. Salazinic acid attenuates male sexual dysfunction and testicular oxidative damage in streptozotocin-induced diabetic albino rats. RSC Adv. 2023, 13, 12991–13005. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, D.S.N.B.K.; Singh, G.; Panda, S.P.; Achanti, S.; Soni, H.; Chaudhuri, T.K.; Pawar, H.A.; Jahasultana, M. In Silico Screening of Plant-Derived Anti-virals from Shorea hemsleyana (King) King ex Foxw Against SARS CoV-2 Main Protease. Chem. Afr. 2023, 6, 345–366. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2020, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Gündoğdu, S.; Türkeş, C.; Arslan, M.; Demir, Y.; Beydemir, Ş. New Isoindole-1,3-dione Substituted Sulfonamides as Potent Inhibitors of Carbonic Anhydrase and Acetylcholinesterase: Design, Synthesis, and Biological Evaluation. ChemistrySelect 2019, 4, 13347–13355. [Google Scholar] [CrossRef]
- Visualizer, B.D.S. Dassault Systemes BIOVIA, Discovery Studio Visualizer, Version 17.2; Dassault Systemes: San Diego, CA, USA, 2021.
- Gangadharappa, B.S.; Sharath, R.; Revanasiddappa, P.D.; Chandramohan, V.; Balasubramaniam, M.; Vardhineni, T.P. Structural insights of metallo-beta-lactamase revealed an effective way of inhibition of enzyme by natural inhibitors. J. Biomol. Struct. Dyn. 2020, 38, 3757–3771. [Google Scholar] [CrossRef]
- Kumari, R.; Kumar, R.; Open Source Drug Discovery, C.; Lynn, A. g_mmpbsa—A GROMACS tool for high-throughput MM-PBSA calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef]
- Haider, S.; Tabassum, S.; Perveen, T. Scopolamine-induced greater alterations in neurochemical profile and increased oxidative stress demonstrated a better model of dementia: A comparative study. Brain Res. Bull. 2016, 127, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, L.H.; Jia, W.; Liu, X.M.; Dang, H.X.; Mai, W.L.; Wang, N.; Steinmetz, A.; Wang, Y.Q.; Xu, C.J. Comparison of ginsenosides Rg1 and Rb1 for their effects on improving scopolamine-induced learning and memory impairment in mice. Phytother. Res. 2010, 24, 1748–1754. [Google Scholar] [CrossRef] [PubMed]
- Cook, L.; Weidley, E. Behavioral effects of some psychopharmacological agents. Ann. N. Y. Acad. Sci. 1957, 66, 740–752. [Google Scholar] [CrossRef]
- Goverdhan, P.; Sravanthi, A.; Mamatha, T. Neuroprotective effects of meloxicam and selegiline in scopolamine-induced cognitive impairment and oxidative stress. Int. J. Alzheimers Dis. 2012, 2012, 974013. [Google Scholar] [CrossRef]
- Abo Mansour, H.E.; Elberri, A.I.; Ghoneim, M.E.; Samman, W.A.; Alhaddad, A.A.; Abdallah, M.S.; El-Berri, E.I.; Salem, M.A.; Mosalam, E.M. The Potential Neuroprotective Effect of Thymoquinone on Scopolamine-Induced In Vivo Alzheimer’s Disease-like Condition: Mechanistic Insights. Molecules 2023, 28, 6566. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Nishikimi, M.; Appaji, N.; Yagi, K. The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochem. Biophys. Res. Commun. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Maryland Heights, MO, USA, 2008. [Google Scholar]
- Ahmad, A.; Ramasamy, K.; Jaafar, S.M.; Majeed, A.B.; Mani, V. Total isoflavones from soybean and tempeh reversed scopolamine-induced amnesia, improved cholinergic activities and reduced neuroinflammation in brain. Food Chem. Toxicol. 2014, 65, 120–128. [Google Scholar] [CrossRef]
- Norinder, U.; Bergstrom, C.A. Prediction of ADMET Properties. ChemMedChem 2006, 1, 920–937. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.V.; Thakur, S.; Kumar, N.; Singh, H.; Mithu, V.S.; Singh, H.; Bhagat, K.; Gulati, H.K.; Sharma, A.; Singh, H.; et al. Donepezil-Inspired Multitargeting Indanone Derivatives as Effective Anti-Alzheimer’s Agents. ACS Chem. Neurosci. 2022, 13, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Choudhir, G.; Sharma, S.; Hariprasad, P. A combinatorial approach to screen structurally diverse acetylcholinesterase inhibitory plant secondary metabolites targeting Alzheimer’s disease. J. Biomol. Struct. Dyn. 2022, 40, 11705–11718. [Google Scholar] [CrossRef] [PubMed]
- Garlich, F.M.; Balakrishnan, K.; Shah, S.K.; Howland, M.A.; Fong, J.; Nelson, L.S. Prolonged altered mental status and bradycardia following pediatric donepezil ingestion. Clin. Toxicol. 2014, 52, 291–294. [Google Scholar] [CrossRef]
- McLaren, A.T.; Allen, J.; Murray, A.; Ballard, C.G.; Kenny, R.A. Cardiovascular effects of donepezil in patients with dementia. Dement. Geriatr. Cogn. Disord. 2003, 15, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, D.; Panda, S.P.; Rao, A.L.; Chakravarti, G.; Teja, N.; Vani, V.B.N.; Sandhya, T. In-silico strategies of some selected phytoconstituents from zingiber officinale as SARS CoV-2 main protease (COVID-19) inhibitors. Indian J. Pharm. Educ. Res. 2020, 54, s552–s559. [Google Scholar] [CrossRef]
- Hassan, M.; Raza, H.; Abbasi, M.A.; Moustafa, A.A.; Seo, S.-Y. The exploration of novel Alzheimer’s therapeutic agents from the pool of FDA approved medicines using drug repositioning, enzyme inhibition and kinetic mechanism approaches. Biomed. Pharmacother. 2019, 109, 2513–2526. [Google Scholar] [CrossRef] [PubMed]
- Al-Mustafa, A.; Al-Zereini, W.; Ashram, M.; Al-Sha’er, M.A. Evaluation of antibacterial, antioxidant, cytotoxic, and acetylcholinesterase inhibition activities of novel [1,4] benzoxazepines fused to heterocyclic systems with a molecular modeling study. Med. Chem. Res. 2023, 32, 239–253. [Google Scholar] [CrossRef]
- Ibrar, A.; Khan, A.; Ali, M.; Sarwar, R.; Mehsud, S.; Farooq, U.; Halimi, S.M.A.; Khan, I.; Al-Harrasi, A. Combined in Vitro and in Silico Studies for the Anticholinesterase Activity and Pharmacokinetics of Coumarinyl Thiazoles and Oxadiazoles. Front. Chem. 2018, 6, 61. [Google Scholar] [CrossRef]
- Mohankumar, T.; Chandramohan, V.; Lalithamba, H.S.; Jayaraj, R.L.; Kumaradhas, P.; Sivanandam, M.; Hunday, G.; Vijayakumar, R.; Balakrishnan, R.; Manimaran, D.; et al. Design and Molecular dynamic Investigations of 7,8-Dihydroxyflavone Derivatives as Potential Neuroprotective Agents Against Alpha-synuclein. Sci. Rep. 2020, 10, 599. [Google Scholar] [CrossRef] [PubMed]
- Prasanth, D.A.-O.; Manikanta, M.A.-O.; Chandramohan, V.; Bhavya, G.; Lakshmana Rao, A.; Panda, S.P.; Rao, G.; Chakravarthi, G.; Teja, N.; Suguna Rani, P.; et al. In-silico strategies of some selected phytoconstituents from Melissa officinalis as SARS CoV-2 main protease and spike protein (COVID-19) inhibitors. Mol. Simul. 2021, 47, 457–470. [Google Scholar] [CrossRef]
- Hsieh, M.T.; Hsieh, C.L.; Lin, L.W.; Wu, C.R.; Huang, G.S. Differential gene expression of scopolamine-treated rat hippocampus-application of cDNA microarray technology. Life Sci. 2003, 73, 1007–1016. [Google Scholar] [CrossRef]
- Umukoro, S.; Adewole, F.A.; Eduviere, A.T.; Aderibigbe, A.O.; Onwuchekwa, C. Free radical scavenging effect of donepezil as the possible contribution to its memory enhancing activity in mice. Drug Res. 2014, 64, 236–239. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | MW | logp | Alogp | HBA | HBD | TPSA | AMR |
|---|---|---|---|---|---|---|---|
| CTN | 154.14 | 3.591 | 2.043 | 1 | 0 | 17.07 | 48.38 |
| DON | 349.99 | 2.633 | 0.364 | 4 | 0 | 38.77 | 115.79 |
| Phytocompound | Parameters | Values | |
|---|---|---|---|
| CTN | Swiss ADME | log P o/w | 3.83 |
| Water Solubility | Soluble | ||
| GI Absorption | High | ||
| Lipinski Rule | Yes | ||
| Veber’s Rule | Yes | ||
| PAINS Alert | 0 | ||
| TPSA | 17.07 | ||
| ADMETSAR | HIA | 0.9905 | |
| CaCO2 | 0.7608 | ||
| BBB | 0.9664 | ||
| CYP1A2 | 0.6175 | ||
| CYP2C19 | 0.9168 | ||
| CYP2C9 | 0.9225 | ||
| CYP2D6 | 0.9572 | ||
| PROTOX-II | LD50 (mg/kg) | 2420 (Class 5) | |
| Hepatotoxicity | Inactive | ||
| Carcinogenicity | Inactive | ||
| Immunotoxicity | Inactive | ||
| Mutagenicity | Inactive | ||
| Cytotoxicity | Inactive | ||
| DON | Swiss ADME | log P o/w | 3.92 |
| Water Solubility | Moderately Soluble | ||
| GI Absorption | High | ||
| Lipinski Rule | Yes | ||
| Veber’s Rule | Yes | ||
| PAINS Alert | 0 | ||
| TPSA | 38.77 | ||
| ADMETSAR | HIA | 0.9966 | |
| CaCO2 | 0.7742 | ||
| BBB | 0.9953 | ||
| CYP1A2 | 0.5072 | ||
| CYP2C19 | 0.8356 | ||
| CYP2C9 | 0.8189 | ||
| CYP2D6 | 0.8919 | ||
| PROTOX-II | LD50 (mg/kg) | 505 (Class 4) | |
| Hepatotoxicity | Inactive | ||
| Carcinogenicity | Active | ||
| Immunotoxicity | Active | ||
| Mutagenicity | Inactive | ||
| Cytotoxicity | Active | ||
| Ligands | Binding Affinity, ΔG (Kcal/mol) | Amino Acids Involved and Distance (Å) | |
|---|---|---|---|
| Hydrogen-Bond Interactions | Hydrophobic Interactions | ||
| CTN | −6.5 | TYR A:337 (5.21), TYR A:341 (6.26) | TYR A:124 (4.65), HIS A:447 (4.85), PHE A:338 (3.90), TYR A:337 (6.41), TYR A:341 (4.70,4.56), TRP A:286 (4.44) |
| DON | −9.2 | TYR A:341 (5.16) | TYR A:72 (5.61), ARP A:86 (4.18), TRP A:286 (5.21), TYR A:337 (4.57), HIS A:447 (5.50), PHE A:338 (5.38), VAL A:294 (4.47) |
| S. No. | Protein | RMSD (nm) | RMSF (nm) | Rg (nm) | SASA (nm 2) |
|---|---|---|---|---|---|
| 1 | CTN | 0.3 | 1.11 | 2.31 | 219.2 |
| 2 | DON | 0.27 | 1.31 | 2.32 | 223.1 |
| Binding Site Residues (aa) | kJ/mol | Binding Site Residues (aa) | kJ/mol |
|---|---|---|---|
| TYR-77 | 0.9165 | TYR-72 | −2.2201 |
| PRO-78 | 0.1434 | VAL-73 | −3.3021 |
| GLY-79 | −1.2396 | ASP-74 | −3.4861 |
| THR-83 | 0.5039 | THR-75 | 0.1518 |
| GLU-84 | −3.5967 | LEU-76 | −5.9723 |
| TRP-86 | −2.1014 | TRP-86 | −2.3365 |
| SER-125 | 1.1953 | THR-83 | 0.0273 |
| TRP-286 | −3.5095 | ASN-87 | 4.2641 |
| LEU-289 | −0.361 | GLY-121 | −1.8082 |
| GLN-291 | −0.005 | GLY-122 | 0.0616 |
| GLU-292 | −0.9968 | TYR-124 | −6.2327 |
| PHE-295 | 1.1175 | SER-125 | 1.5916 |
| ARG-296 | 6.215 | TRP-286 | −5.4894 |
| PHE-297 | −1.9667 | LEU-289 | −3.0761 |
| PHE-338 | −2.0933 | PRO-290 | −0.3583 |
| TYR-341 | −1.3473 | GLN-291 | −0.821 |
| GLY-122 | 0.3881 | GLU-292 | −5.3739 |
| PHE-123 | −1.3959 | SER-293 | −1.0867 |
| TYR-124 | −4.0743 | PHE-295 | 0.6986 |
| −12.2078 | ARG-296 | 1.5049 | |
| PHE-297 | −4.7205 | ||
| PHE-338 | −4.8894 | ||
| TYR-341 | 0.8496 | ||
| GLY-342 | −2.329 | ||
| ALA-343 | −2.5718 | ||
| PRO-344 | −1.0723 | ||
| −47.9969 |
| Treatment | CAT (n Moles of H2O2 Consumed/Minute/mg Protein) | GSH (nmol/mg/Protein) | SOD (Units per mg of Protein) | AchE (nmol/min/mg Protein) |
|---|---|---|---|---|
| Negative control | 8.19 ±0.15 | 3.2 ± 0.09 | 11.07 ± 0.64 | 0.051 ± 0.0098 |
| Positive control | 1.6 ± 0.104 *** | 0.34 ± 0.08 *** | 2.36 ± 012 *** | 0.44 ± 0.02 *** |
| DON (0.5 mg/kg, p.o) + SCO (1 mg/kg, i.p, 14 days) | 6.29 ± 0.37 *** | 2.36 ± 0.30 *** | 6.53 ± 0.14 *** | 0.13 ± 0.01 *** |
| CTN (25 mg/kg. I.P) + SCO (1 mg/kg, i.p, 14 days) | 3.15 ± 0.3 *** | 1.65 ± 0.26 *** | 5.15 ± 0.32 *** | 0.27 ± 0.031 ** |
| CTN (50 mg/kg. I.P) + SCO (1 mg/kg, i.p, 14 days) | 6.36 ± 0.19 *** | 2.02 ± 0.29 *** | 7.28 ± 0.53 *** | 0.18 ± 0.01 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
K, P.; Prasanth, D.S.N.B.K.; Shadakshara, M.K.R.; Ahmad, S.F.; Seemaladinne, R.; Rudrapal, M.; Pasala, P.K. Citronellal as a Promising Candidate for Alzheimer’s Disease Treatment: A Comprehensive Study on In Silico and In Vivo Anti-Acetylcholine Esterase Activity. Metabolites 2023, 13, 1133. https://doi.org/10.3390/metabo13111133
K P, Prasanth DSNBK, Shadakshara MKR, Ahmad SF, Seemaladinne R, Rudrapal M, Pasala PK. Citronellal as a Promising Candidate for Alzheimer’s Disease Treatment: A Comprehensive Study on In Silico and In Vivo Anti-Acetylcholine Esterase Activity. Metabolites. 2023; 13(11):1133. https://doi.org/10.3390/metabo13111133
Chicago/Turabian StyleK, Pavani, D S. N. B. K. Prasanth, Murthy K. R. Shadakshara, Sheikh F. Ahmad, Ramanjaneyulu Seemaladinne, Mithun Rudrapal, and Praveen Kumar Pasala. 2023. "Citronellal as a Promising Candidate for Alzheimer’s Disease Treatment: A Comprehensive Study on In Silico and In Vivo Anti-Acetylcholine Esterase Activity" Metabolites 13, no. 11: 1133. https://doi.org/10.3390/metabo13111133
APA StyleK, P., Prasanth, D. S. N. B. K., Shadakshara, M. K. R., Ahmad, S. F., Seemaladinne, R., Rudrapal, M., & Pasala, P. K. (2023). Citronellal as a Promising Candidate for Alzheimer’s Disease Treatment: A Comprehensive Study on In Silico and In Vivo Anti-Acetylcholine Esterase Activity. Metabolites, 13(11), 1133. https://doi.org/10.3390/metabo13111133