

Lipid and Amino Acid Pathway Metabolites Contribute to Cold Tolerance in Quercus wutaishanica


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Analysis of Physiological Parameters
2.3. Untargeted Metabolomics Analysis
2.4. Statistical Analysis
3. Results
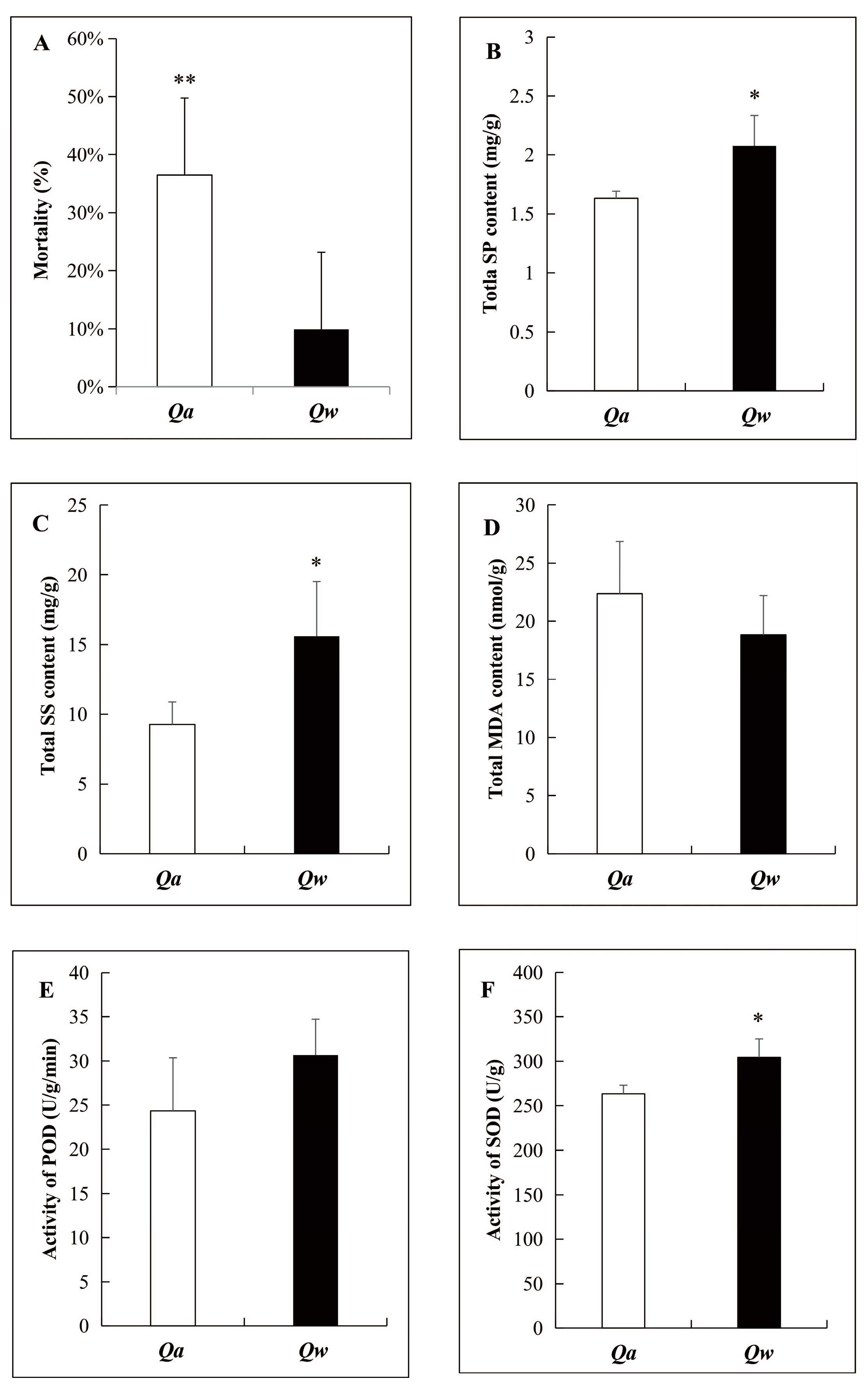
3.1. Overwintering Mortality Ratio and Physiological Responses of Q. acutissima and Q. wutaishanica to LTs
3.2. Detection of Metabolites by LC-MS/MS
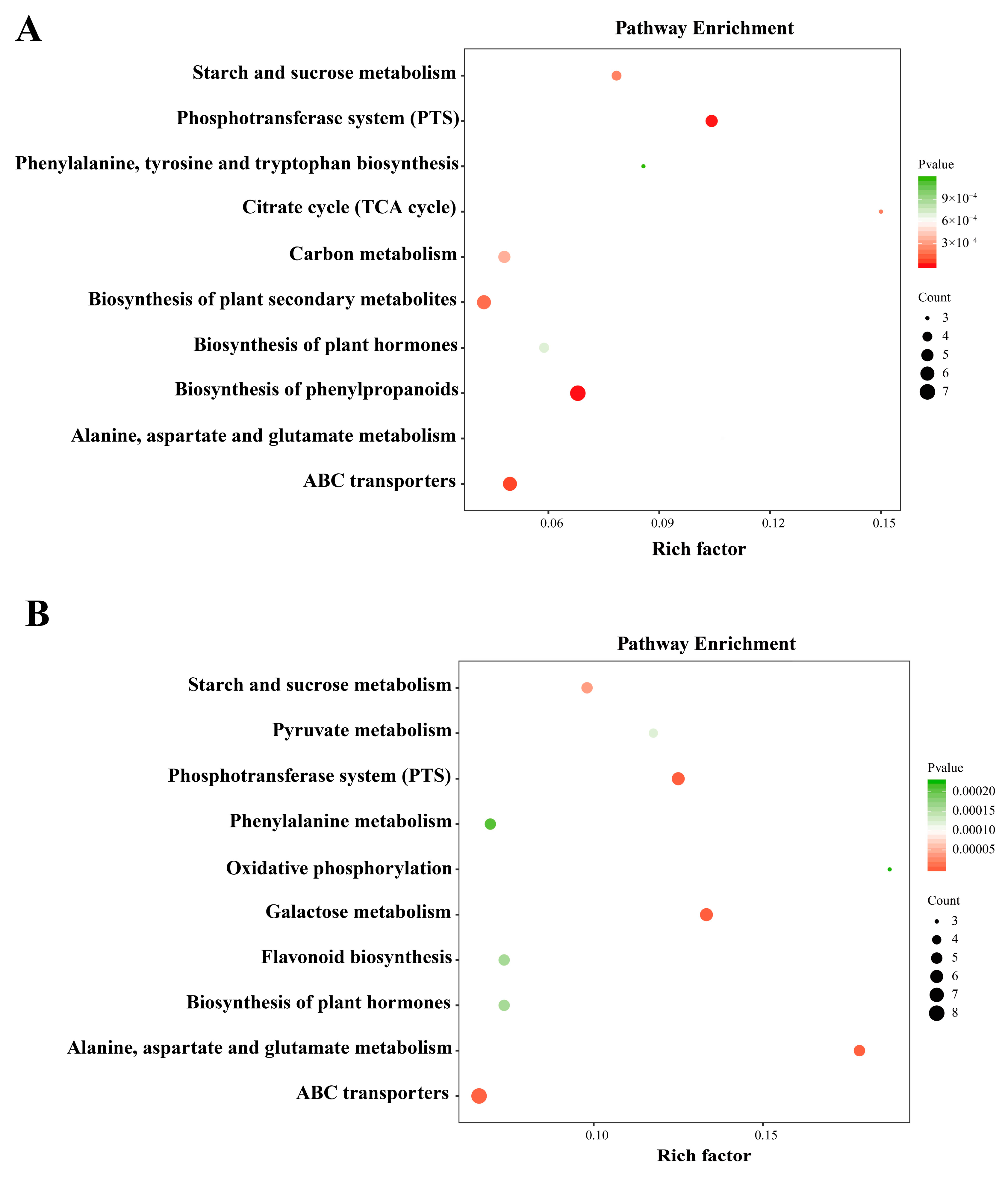
3.3. Pathway Enrichment and Topological Analysis of Differentially Accumulated Metabolites in the Two Oak Species
4. Discussion
4.1. Q. wutaishanica has Higher Cold Tolerance Than Q. accutissima
4.2. Universal Cold Resistance Mechanisms of Oaks
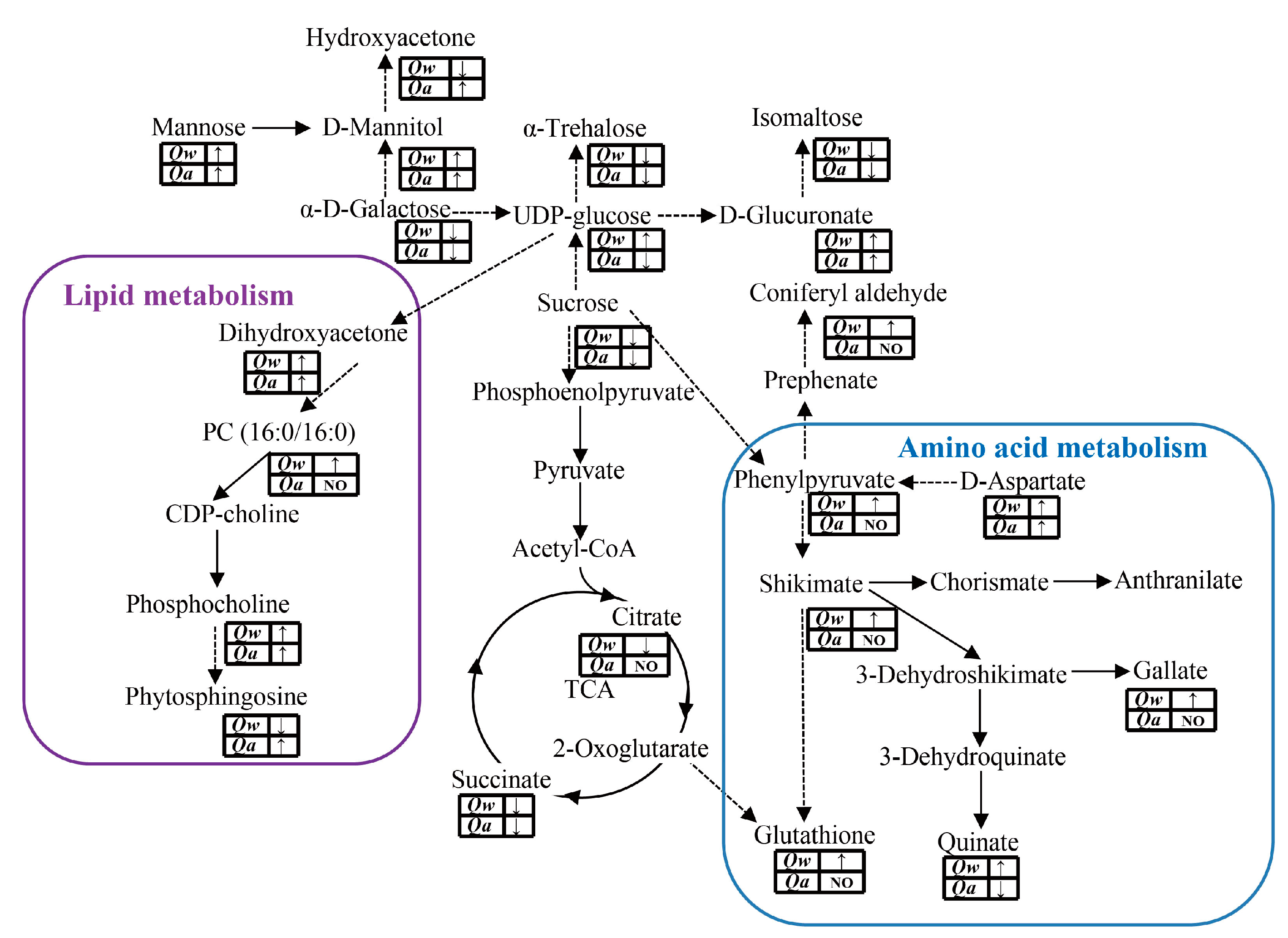
4.3. Lipid and Amino Acid Pathway Metabolites May Contribute to Cold Tolerance in Q. wutaishanica
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nixon, K.C. Quercus Linnaeus; Flora of North America Editorial Committee, Ed.; Flora of North America: North of Mexico City, Mexico; Oxford University Press: New York, NY, USA, 1997; pp. 431–506. [Google Scholar]
- Sánchez, J.A.B.; Ramírez, M.V.M.; Larrosa, M.M.; Gallegos, J.A.I.; González, R.F.L.; Medina, L.T.; Rocha, N.E.G. Antioxidant, antimicrobial, antitopoisomerase and gastroprotective effect of herbal infusions from four Quercus species. Ind. Crops Prod. 2012, 42, 57–62. [Google Scholar] [CrossRef]
- Barta, C.E.; Bolander, B.; Bilby, S.R.; Brown, J.H.; Brown, R.N.; Duryee, A.M.; Edelman, D.R.; Gray, C.E.; Gossett, C.; Haddock, A.G.; et al. In situ dark adaptation enhances the efficiency of DNA extraction from Mature Pin Oak (Quercus palustris) leaves, facilitating the identification of partial sequences of the 18S rRNA and isoprene synthase (IspS) genes. Plants 2017, 6, 52. [Google Scholar] [CrossRef]
- Kroeger, T.; Escobedo, F.J.; Hernandez, J.L.; Varela, S.; Delphin, S.; Fisher, J.R.B.; Waldron, J. Reforestation as a novel abatement and compliance measure for ground-level ozone. Proc. Natl. Acad. Sci. USA 2014, 111, E4204–E4213. [Google Scholar] [CrossRef]
- Qin, L.; Li, S.Y. Sericulture of Chinese Oak Silkworm; China Agricultural Press: Beijing, China, 2017; pp. 35–36. (In Chinese) [Google Scholar]
- Wang, L.; Lang, Q.; Xia, X.; Sun, J.; Gao, W.; Wang, F.; Na, K. Investigation and protective measure of oaks frost damage in Dandong in the Spring of 2008. North Seric. 2009, 30, 27–30. [Google Scholar]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Peñuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef]
- Szuba, A.; Kalemba, E.M.; Wawrzyniak, M.K.; Suszka, J.; Cheielarz, P. Deterioration in the quality of recalcitrant Quercus robur seeds during six months of storage at subzero temperatures: Ineffective activation of prosurvival mechanisms and evidence of freezing stress from an untargeted metabolomic study. Metabolites 2022, 12, 756. [Google Scholar] [CrossRef]
- Guerrero-Sánchez, V.M.; Lόpez-Hidalgo, C.; Rey, M.; Castillejo, M.Á.; Jorrín-Novo, J.V.; Escandón, M. Multiomic data integration in the analysis of drought-responsive mechanisms in Quercus ilex seedlings. Plants 2022, 11, 3067. [Google Scholar] [CrossRef]
- Wani, M.A.; Jan, N.; Qazi, H.A.; Andrabi, K.I.; John, R. Cold stress induces biochemical changes, fatty acid profile, antioxidant system and gene expression in Capsella bursa pastoris L. Acta Physiol. Plant 2018, 40, 167. [Google Scholar] [CrossRef]
- Peng, M.; Chang, Y.; Chu, G.; Wang, M. Low-temperature tolerance and transcriptome analyses during seed germination of Anabasis aphylla. J. Plant Interact. 2019, 14, 254–264. [Google Scholar] [CrossRef]
- Guy, C.L. Cold acclimation and freezing stress tolerance: Role of protein metabolism. Annu. Rev. Plant Phys. 1990, 41, 187–223. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, F.; Guy, C.L. β-Amylase induction and the protective role of maltose during temperature shock. Plant Physiol. 2004, 135, 1674–1684. [Google Scholar] [CrossRef]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.M.; Qian, P.; Xin, W.; Li, H.Y.; Burritt, D.J.; Fujita, M.; Tran, L.P. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef]
- Han, C.; Jin, P.; Li, M.; Wang, L.; Zheng, Y. Physiological and transcriptomic analysis validates previous findings of changes in primary metabolism for the production of phenolic antioxidants in wounded carrots. J. Agric. Food Chem. 2017, 65, 7159–7167. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araujo, W.L.; Braun, H.P. Amino acid catabolism in plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Bocian, A.; Zwierzykowski, Z.; Rapaca, M.; Koczyk, G.; Ciesiolka, D.; Kosmala, A. Metabolite profiling during cold acclimation of Lolium perenne genotypes distinct in the level of frost tolerance. J. Appl. Genet. 2015, 56, 439–449. [Google Scholar] [CrossRef]
- Oberschelp, G.P.J.; Guarnascheli, A.B.; Teson, N.; Harrand, L.; Podestá, F.E.; Margarit, E. Cold acclimation and freezing tolerance in three Eucalyptus species: A metabolomic and proteomic approach. Plant Physiol. Biochem. 2020, 154, 316–327. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Miguel, C.; Chaves, I.; António, C. Mass spectrometry-based forest tree metabolomics. Mass Spectrom. Rev. 2019, 40, 126–157. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Liao, W.J.; Petit, R.J.; Zhang, D.Y. Geographic variation in the structure of oak hybrid zones provides insights into the dynamics of speciation. Mol. Ecol. 2011, 20, 4995–5011. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, L.I.; Fang, Y. Geographical distribution and prediction of potential ranges of Quercus acutissima in China. Acta Bot. Boreali Occident. Sin. 2014, 34, 1685–1692. [Google Scholar]
- Xiong, S.; Wang, Y.; Chen, Y.; Gao, M.; Zhao, Y.; Wu, L. Effects of drought stress and rehydration on physiological and biochemical properties of four oak species in China. Plants 2022, 11, 679. [Google Scholar] [CrossRef]
- Adusumilli, R.; Mallick, P. Data conversion with proteowizard msconvert. Methods Mol. Biol. 2017, 1550, 339–368. [Google Scholar] [CrossRef] [PubMed]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, P.C. 1H nuclear magnetic resonance (NMR)-based metabolome diversity of Seabuckthorn (H. rhamnoides L.) berries originating from two geographical regions of Indian Himalayas. Food Anal. Method 2022, 15, 157–171. [Google Scholar] [CrossRef]
- Gemperlin, E.; Keller, C.; Li, L. Mass spectrometry in plant-omics. Anal. Chem. 2016, 88, 3422–3434. [Google Scholar] [CrossRef]
- Yang, M.; Yang, J.; Su, L.; Sun, K.; Li, D.; Liu, Y.; Wang, H.; Chen, Z.; Guo, T. Metabolic profile analysis and identification of key metabolites during rice seed germination under low-temperature stress. Plant Sci. 2019, 289, 110282. [Google Scholar] [CrossRef]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0-making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef]
- Shen, W.; Nada, K.; Tachibana, S. Involvement of polyamines in the chilling tolerance of cucumber cultivars. Plant Physiol. 2000, 124, 431–439. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Du, S.; Cui, M.; Cai, Y.; Xue, A.; Hao, Y.; Huang, X.; Liu, L.; Luo, L. Metabolomic analysis of chilling response in rice (Oryza sativa L.) seedlings by extractive electrospray ionization mass spectrometry. Environ. Exp. Bot. 2020, 180, 104231. [Google Scholar] [CrossRef]
- He, J.; Yao, L.; Pecoraro, L.; Liu, C.; Wang, J.; Huang, L.; Gao, W. Cold stress regulates accumulation of flavonoids and terpenoids in plants by phytohormone, transcription process, functional enzyme, and epigenetics. Crit. Rev. Biotechnol. 2022, 43, 680–697. [Google Scholar] [CrossRef]
- Morin, X.; Améglio, T.; Ahas, R.; Kurz-besson, C.; Lanta, V.; Lebourgeois, F.; Miglietta, F.; Chuine, I. Variation in cold hardiness and carbohydrate concentration from dormancy induction to bud burst among provenances of three European oak species. Tree Physiol. 2007, 27, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Stoop, J.M.H.; Williamson, J.D.; Pharr, D.M. Mannitol metabolism in plants: A method for coping with stress. Trends Plant Sci. 1996, 1, 139–144. [Google Scholar] [CrossRef]
- Cao, X.C. Responses of Sucrose Metabolism and Antioxidant Enzymes of Dendrobium Catenatum to Stress; Zhejiang A and F University: Hangzhou, China, 2017. [Google Scholar]
- Zhang, J.; Li, J.Q.; Wang, X.C.; Chen, J. OVP1, a vacuolar H+-translocation inorganic pyrophosphatase (V-PPase), overexpression improved rice cold tolerance. Plant Physiol. Biochem. 2011, 49, 33–38. [Google Scholar] [CrossRef]
- Song, S.Y.; Chen, Y.; Chen, J.; Dai, X.Y.; Zhang, W.H. Physiological mechanisms underlying Os Nac5-dependent tolerance of rice plants to abiotic stress. Planta 2011, 234, 331–345. [Google Scholar] [CrossRef]
- Benina, M.; Obata, T.; Mehterov, N.; Ivanov, I.; Petrov, V.; Toneva, V.; Fernie, A.R.; Gechev, T.S. Comparative metabolic profiling of Haberlea rhodopensis, Thellungiella halophyla, and Arabidopsis thaliana exposed to low temperature. Front. Plant Sci. 2013, 4, 499. [Google Scholar] [CrossRef]
- Korn, M.; Gärtner, T.; Erban, A.; Kopka, J.; Selbig, J.; Hincha, D.K. Predicting Arabidopsis freezing tolerance and heterosis in freezing tolerance from metabolite composition. Mol. Plant 2010, 3, 224–235. [Google Scholar] [CrossRef]
- Pagter, M.; Alpers, J.; Erban, A.; Kopka, J.; Zuther, E.; Hincha, D. Rapid transcriptional and metabolic regulation of the deacclimation process in cold acclimated Arabidopsis thaliana. BMC Genom. 2017, 18, 731. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Yu, M.Y.; Han, Y.; Tai, B.; Ni, S.F.; Ji, R.F.; Pu, C.J.; Chen, K.; Li, F.Q.; Xiao, H.; et al. Comparative transcriptomics and metabolites analysis of two closely related Euphorbia species reveal environmental adaptation mechanism and active ingredients difference. Front. Plant Sci. 2022, 13, 905275. [Google Scholar] [CrossRef]
- Gao, C.; Mumtaz, M.A.; Zhou, Y.; Yang, Z.; Shu, H.; Zhu, J.; Bao, W.; Cheng, S.; Yin, L.; Huang, J.; et al. Integrated transcriptomic and metabolomics analyses of cold-tolerant and cold-sensitive pepper species reveal key genes and essential metabolic pathways involved in response to cold stress. Int. J. Mol. Sci. 2022, 23, 6683. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.; O’Neill, L.A.J. Succinate: A metabolic signal in inflammation. Trends Cell Biol. 2014, 24, 313–320. [Google Scholar] [CrossRef]
- Pellegrini, E.; Hoshika, Y.; Dusart, N.; Cotrozzi, L.; Gérard, J.; Nali, C.; Vaultier, M.; Jolivet, Y.; Lorenzini, G.; Paoletti, E. Antioxidative responses of three oak species under ozone and water stress conditions. Sci. Total Environ. 2019, 647, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, C.; Zhu, Y.; Zhang, L.; Chen, T.; Zhou, F.; Chen, H.; Lin, Y. The calcium-dependent kinase OsCPK24 functions in cold stress responses in rice. J. Integr. Plant Biol. 2018, 60, 173–188. [Google Scholar] [CrossRef]
- Chen, Y.P.; Jia, J.F.; Yue, M. Effect of CO2 laser radiation on physiological tolerance of wheat seedlings exposed to chilling stress. Photochem. Photobiol. 2010, 86, 600–605. [Google Scholar] [CrossRef]
- Ribeiro, P.R.; Femandez, L.G.; de Castro, R.D.; Ligterind, W.; Hilhorst, H.W.M. Physiological and biochemical responses of Ricinus communis seedlings to different temperatures: A metabolomics approach. BMC Plant Biol. 2014, 14, 223–237. [Google Scholar] [CrossRef]
- Sun, C.X.; Gao, X.X.; Li, M.Q.; Fu, J.Q.; Zhang, Y.L. Plastic responses in the metabolome and functional traits of maize plants to temperature variations. Plant Biol. 2016, 18, 249–261. [Google Scholar] [CrossRef]
- Daglia, M.; Lorenzo, A.D.; Nabavi, S.F.; Talas, Z.S.; Nabavi, S.M. Polyphenols: Well beyond the antioxidant capacity: Gallic acid and related compounds as neuroprotective agents: You are what you eat! Curr. Pharm. Biotechnol. 2014, 15, 362–372. [Google Scholar] [CrossRef]
- Świeca, M. Elicitation with abiotic stresses improves pro-health constituents, antioxidant potential and nutritional quality of lentil sprouts. Saudi. J. Biol. Sci. 2015, 22, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Widhalm, J.R.; Qian, Y.; Maeda, H.; Cooper, B.R.; Jannasch, A.S.; Gonda, I.; Lewinsohn, E.; Rhodes, D.; Dudareva, N. An alternative pathway contributes to phenylalanine biosynthesis in plants via a cytosolic tyrosine: Phenylpyruvate aminotransferase. Nat. Commun. 2013, 4, 2833. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Kim, J.K.; Wu, Q.; Park, S.U. Effects of cold stress on transcripts and metabolites in tartary buckwheat (Fagopyrum tataricum). Environ. Exp. Bot. 2018, 155, 488–496. [Google Scholar] [CrossRef]
- Tasseva, G.; Richard, L.; Zachowski, A. Regulation of phosphatidylcholine biosynthesis under salt stress involves choline kinases in Arabidopsis thaliana. FEBS Lett. 2004, 566, 115–120. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Xu, Y.; Liu, Y.-Q.; Qin, L. Lipid and Amino Acid Pathway Metabolites Contribute to Cold Tolerance in Quercus wutaishanica. Metabolites 2023, 13, 1094. https://doi.org/10.3390/metabo13101094
Li Q, Xu Y, Liu Y-Q, Qin L. Lipid and Amino Acid Pathway Metabolites Contribute to Cold Tolerance in Quercus wutaishanica. Metabolites. 2023; 13(10):1094. https://doi.org/10.3390/metabo13101094
Chicago/Turabian StyleLi, Qun, Yang Xu, Yan-Qun Liu, and Li Qin. 2023. "Lipid and Amino Acid Pathway Metabolites Contribute to Cold Tolerance in Quercus wutaishanica" Metabolites 13, no. 10: 1094. https://doi.org/10.3390/metabo13101094
APA StyleLi, Q., Xu, Y., Liu, Y.-Q., & Qin, L. (2023). Lipid and Amino Acid Pathway Metabolites Contribute to Cold Tolerance in Quercus wutaishanica. Metabolites, 13(10), 1094. https://doi.org/10.3390/metabo13101094