
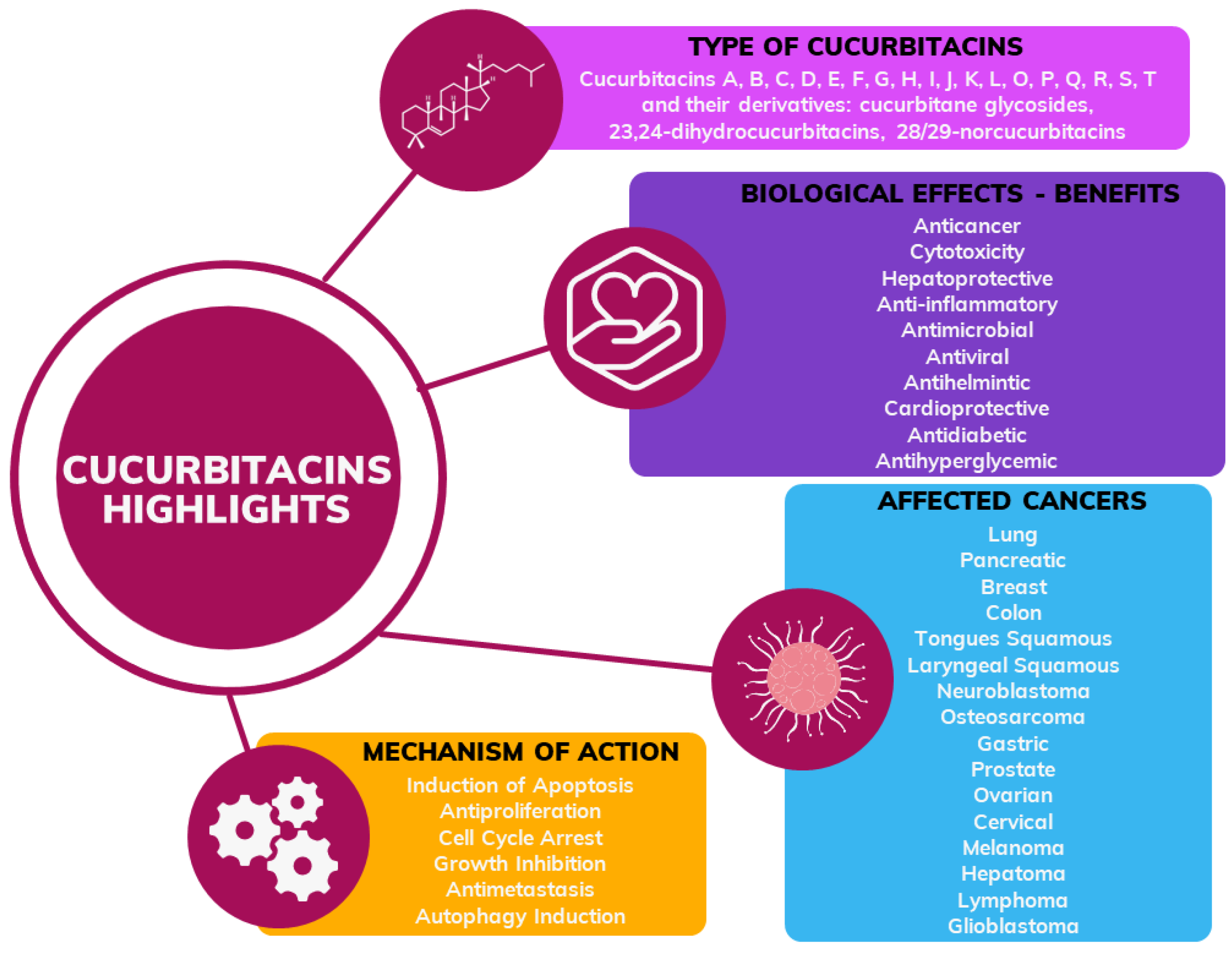
Recent Advances in the Application of Cucurbitacins as Anticancer Agents



Abstract
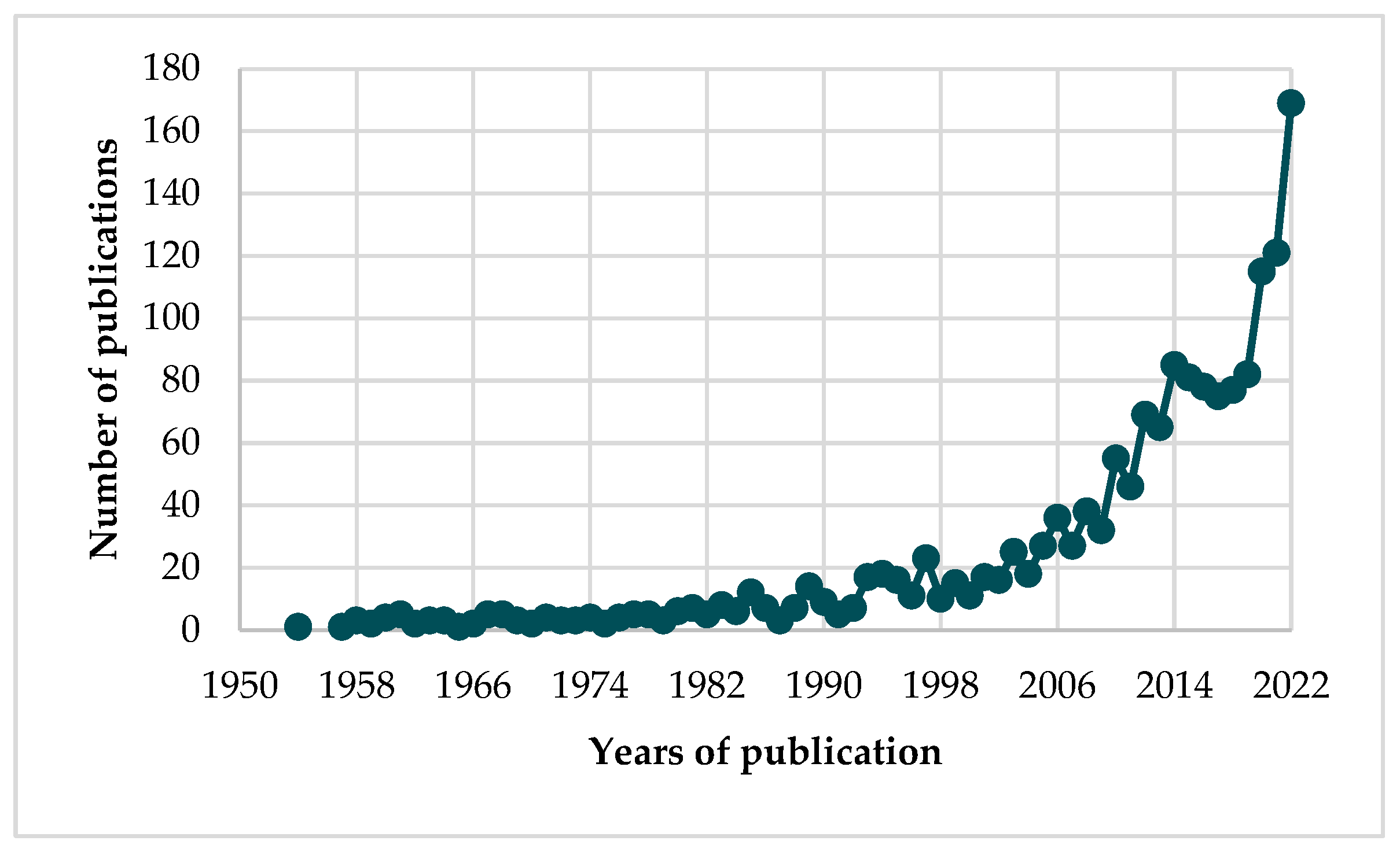
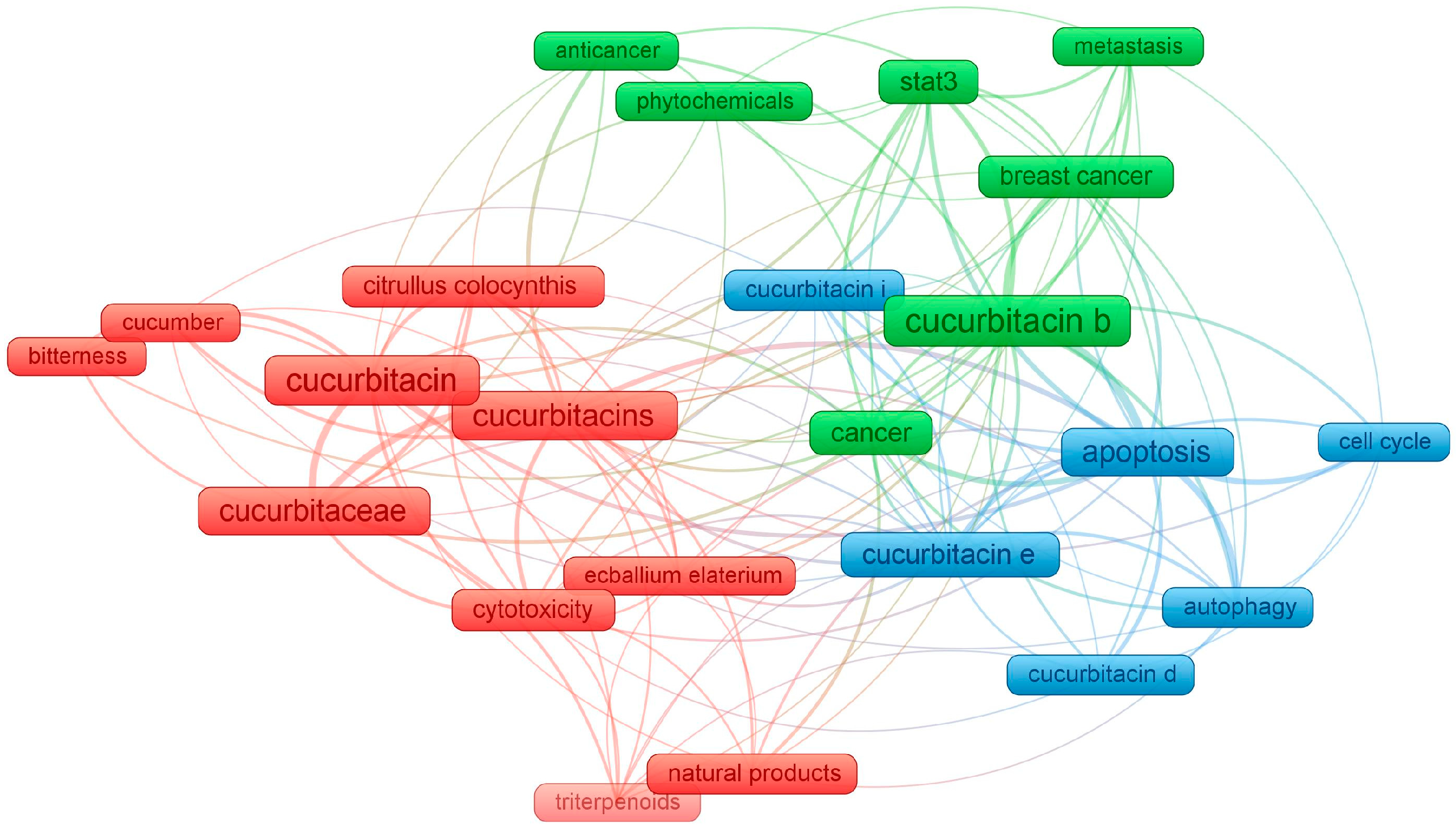
1. Introduction
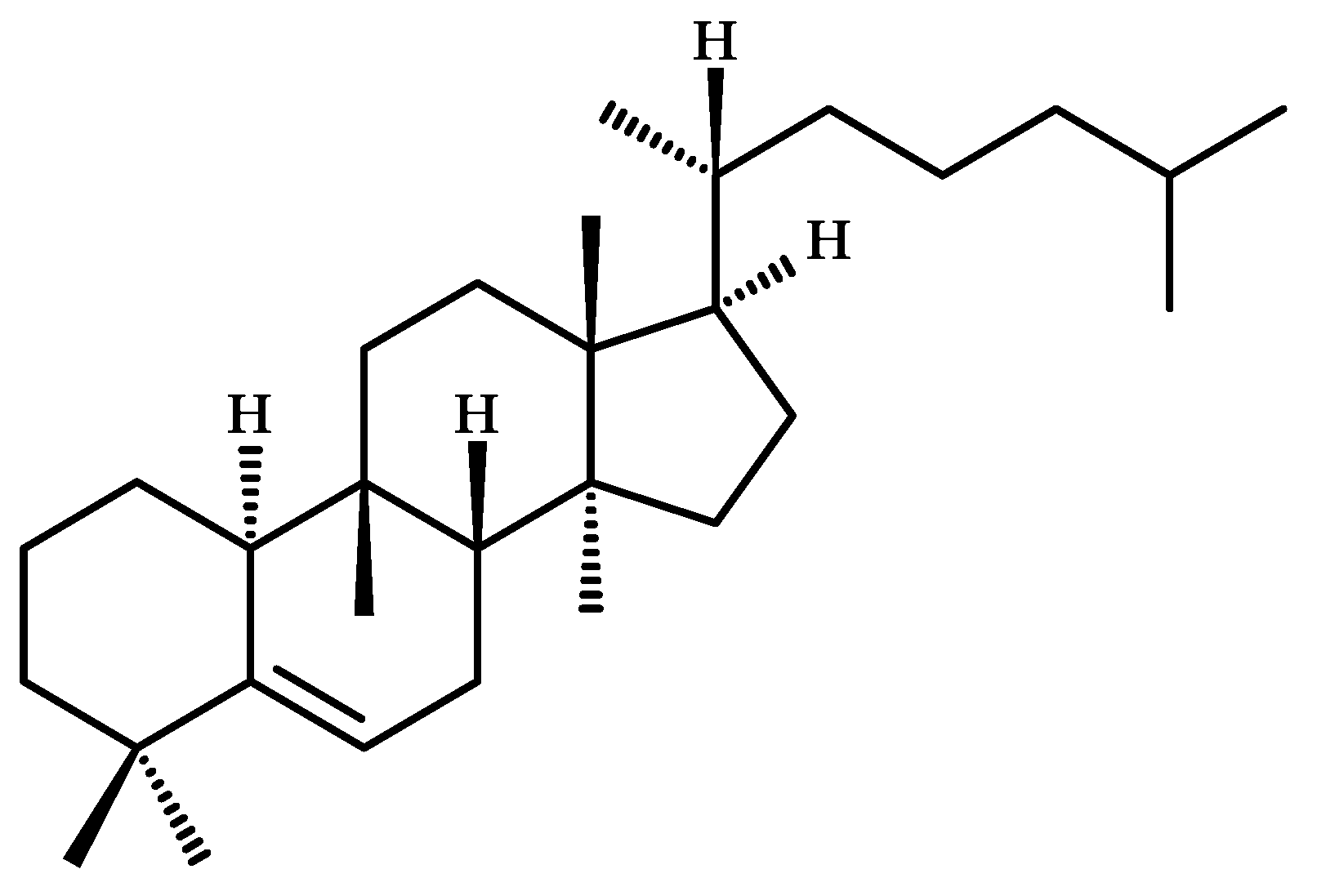
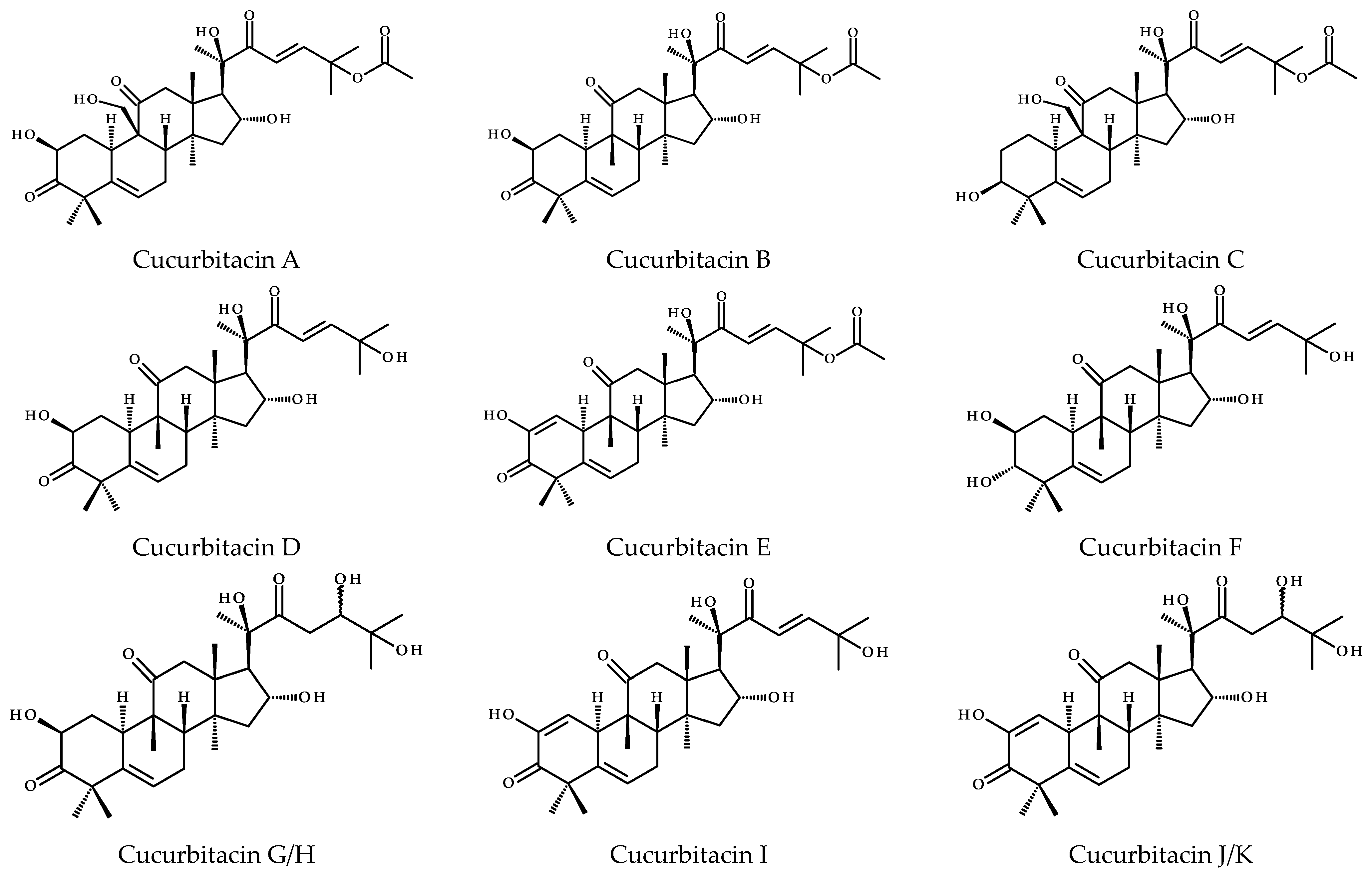
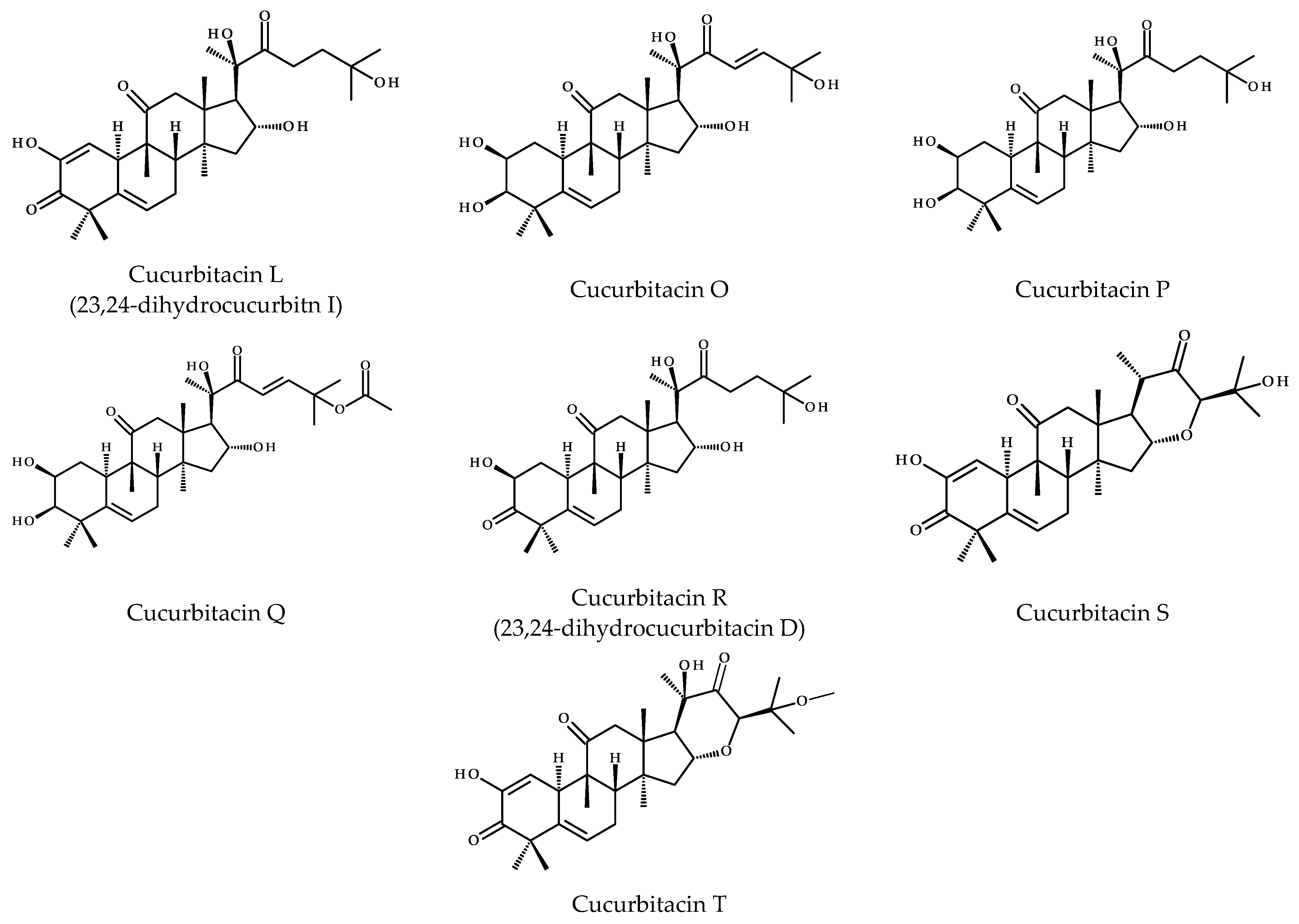
2. Chemical Diversity of Cucurbitacins and Their Occurrence in Various Organisms
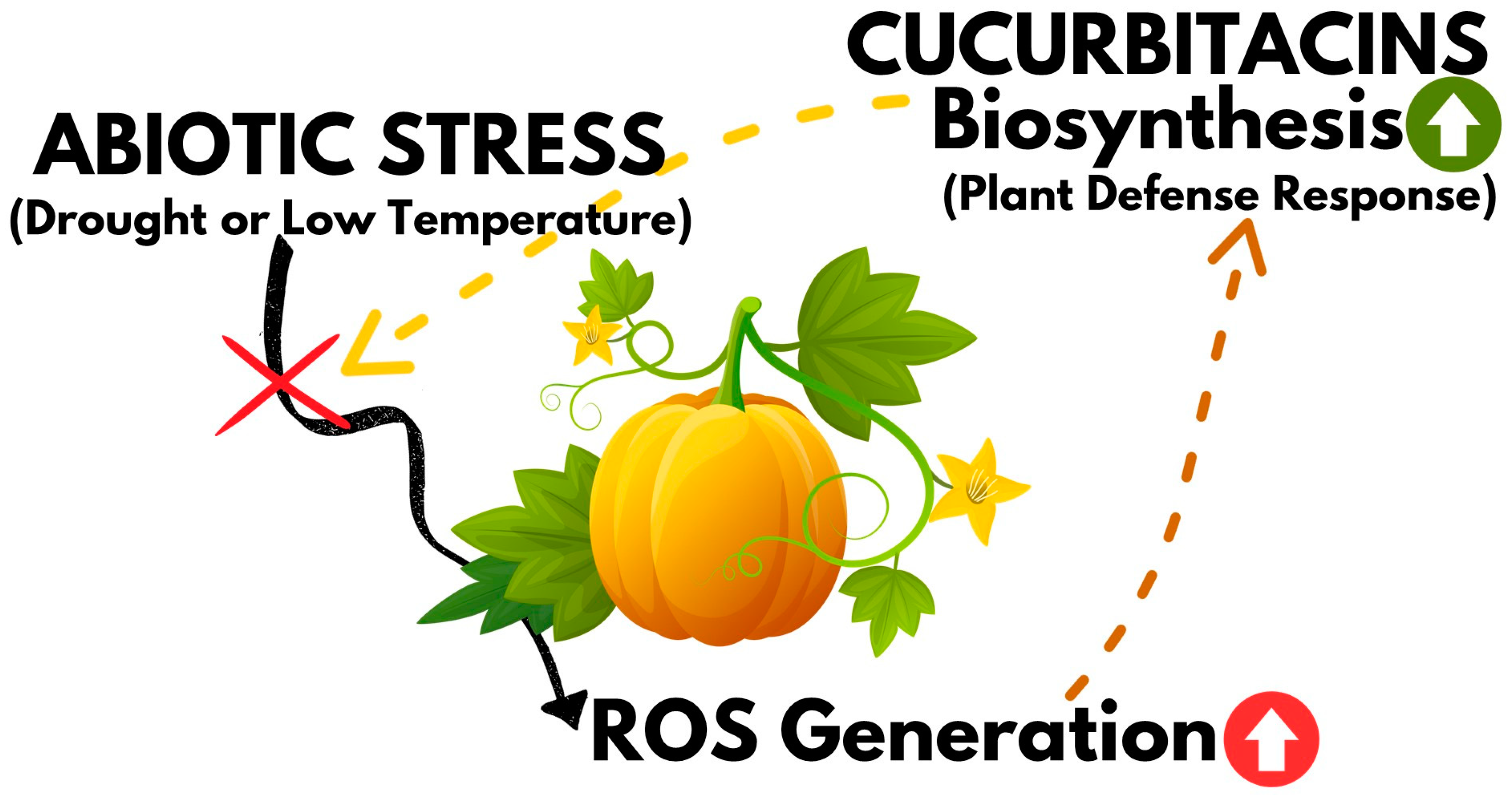
3. Effect of Drought and Temperature on the Biosynthesis of Cucurbitacins
4. The Use of Cucurbitacins in Cancer Research
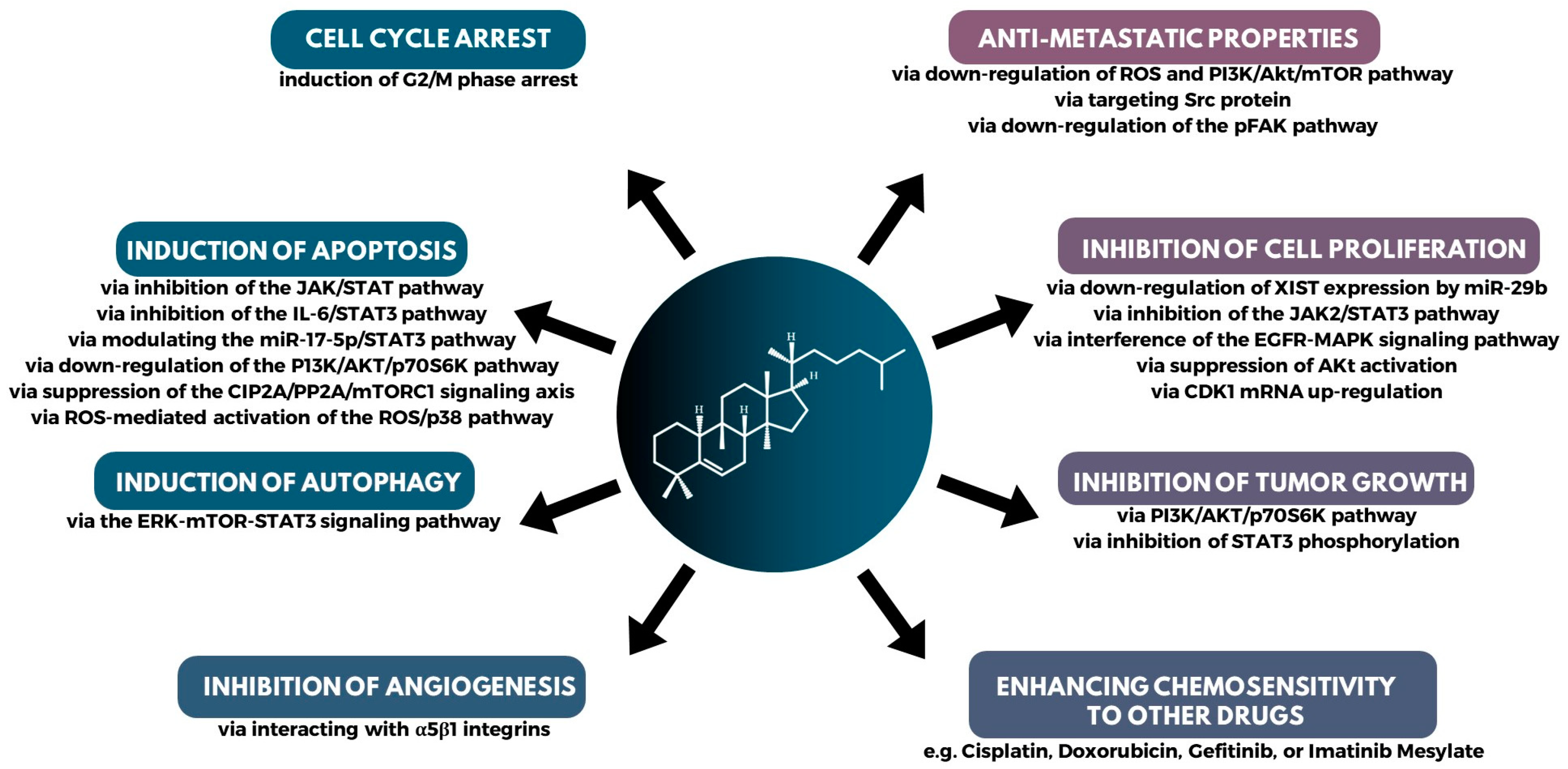
4.1. Biological Activity and Breakthroughs on Cucurbitacin Efficacy
4.2. Mechanism of Action
4.3. Toxicity and Safety Dosage
4.4. Combination Therapy of Cucurbitacins and Other Drugs in Combating Cancer
4.5. Future Directions: Paving the Path Forward
5. Conclusion: The Rising Star of Botanical Therapeutics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Varela, C.; Melim, C.; Neves, B.G.; Sharifi-Rad, J.; Calina, D.; Mamurova, A.; Cabral, C. Cucurbitacins as potential anticancer agents: New insights on molecular mechanisms. J. Transl. Med. 2022, 20, 630. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, Y.; Zeng, J.; Duan, L.; Xue, X.; Wang, H.; Lin, T.; Liu, Z.; Zeng, K.; Zhong, Y.; et al. Convergence and divergence of bitterness biosynthesis and regulation in Cucurbitaceae. Nat. Plants 2016, 2, 16183. [Google Scholar] [CrossRef] [PubMed]
- Alghasham, A.A. Cucurbitacins—A Promising Target for Cancer Therapy. Int. J. Health Sci. 2013, 7, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Cronin, K.A.; Scott, S.; Firth, A.U.; Sung, H.; Henley, S.J.; Sherman, R.L.; Siegel, R.L.; Anderson, R.N.; Kohler, B.A.; Benard, V.B.; et al. Annual report to the nation on the status of cancer, part 1: National cancer statistics. Cancer 2022, 128, 4251–4282. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Jing, S.; Zou, H.; Wu, Z.; Ren, L.; Zhang, T.; Zhang, J.; Wei, Z. Cucurbitacins: Bioactivities and synergistic effect with small-molecule drugs. J. Funct. Foods 2020, 72, 104042. [Google Scholar] [CrossRef]
- Wu, D.; Wang, Z.; Lin, M.; Shang, Y.; Wang, F.; Zhou, J.Y.; Wang, F.; Zhang, X.; Luo, X.; Huang, W. In Vitro and In Vivo Antitumor Activity of Cucurbitacin C, a Novel Natural Product From Cucumber. Front. Pharmacol. 2019, 10, 1287. [Google Scholar] [CrossRef]
- Yuan, R.Q.; Qian, L.; Yun, W.J.; Cui, X.H.; Lv, G.X.; Tang, W.Q.; Cao, R.C.; Xu, H. Cucurbitacins extracted from Cucumis melo L. (CuEC) exert a hypotensive effect via regulating vascular tone. Hypertens. Res. 2019, 42, 1152–1161. [Google Scholar] [CrossRef]
- Chen, C.H.; Kuo, T.C.Y.; Yang, M.H.; Chien, T.Y.; Chu, M.J.; Huang, L.C.; Chen, C.Y.; Lo, H.F.; Jeng, S.T.; Chen, L.F.O. Identification of cucurbitacins and assembly of a draft genome for Aquilaria agallocha. BMC Genom. 2014, 15, 578. [Google Scholar] [CrossRef]
- Kaushik, U.; Aeri, V.; Mir, S.R. Cucurbitacins—An insight into medicinal leads from nature. Pharmacogn. Rev. 2015, 9, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Clericuzio, M.; Mella, M.; Vita-Finzi, P.; Zema, M.; Vidari, G. Cucurbitane Triterpenoids from Leucopaxillus gentianeus. J. Nat. Prod. 2004, 67, 1823–1828. [Google Scholar] [CrossRef] [PubMed]
- Clericuzio, M.; Tabasso, S.; Bianco, M.A.; Pratesi, G.; Beretta, G.; Tinelli, S.; Zunino, F.; Vidari, G. Cucurbitane Triterpenes from the Fruiting Bodies and Cultivated Mycelia of Leucopaxillus gentianeus. J. Nat. Prod. 2006, 69, 1796–1799. [Google Scholar] [CrossRef]
- Enslin, P.R. Bitter principles of the cucurbitaceae. I.—Observations on the chemistry of cucurbitacin A. J. Sci. Food Agric. 1954, 5, 410–416. [Google Scholar] [CrossRef]
- Rolnik, A.; Olas, B. Vegetables from the Cucurbitaceae family and their products: Positive effect on human health. Nutrition 2020, 78, 110788. [Google Scholar] [CrossRef]
- Chen, J.C.; Chiu, M.H.; Nie, R.L.; Cordell, G.A.; Qiu, S.X. Cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Farias, M.R.; Schenkel, E.P.; Mayer, R.; Rucker, G. Cucurbitacins as Constituents of Wilbrandia ebracteata. Planta Med. 1993, 59, 272–275. [Google Scholar] [CrossRef]
- Kawahara, N.; Kurata, A.; Hakamatsuka, T.; Sekita, S.; Satake, M. Two Novel Cucurbitacins, Neocucurbitacins A and B, from the Brazilian Folk Medicine “Buchinha” (Luffa operculata) and Their Effect on PEBP2αA and OCIF Gene Expression in a Human Osteoblast-Like Saos-2 Cell Line. Chem. Pharm. Bull. 2001, 49, 1377–1379. [Google Scholar] [CrossRef][Green Version]
- Salehi, B.; Capanoglu, E.; Adrar, N.; Catalkaya, G.; Shaheen, S.; Jaffer, M.; Giri, L.; Suyal, R.; Jugran, A.K.; Calina, D.; et al. Cucurbits Plants: A Key Emphasis to Its Pharmacological Potential. Molecules 2019, 24, 1854. [Google Scholar] [CrossRef]
- Hylands, P.J.; Magd, M.S. Cucurbitacins from Acanthosicyos horridus. Phytochemistry 1986, 25, 1681–1684. [Google Scholar] [CrossRef]
- Momma, K.; Masuzawa, Y.; Nakai, N.; Chujo, M.; Murakami, A.; Kioka, N.; Kiyama, Y.; Akita, T.; Nagao, M. Direct interaction of Cucurbitacin E isolated from Alsomitra macrocarpa to actin filament. Cytotechnology 2008, 56, 33–39. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tang, Y.; Li, W.; Cao, J.; Li, W.; Zhao, Y. Bioassay-guided isolation and identification of cytotoxic compounds from Bolbostemma paniculatum. J. Ethnopharmacol. 2015, 169, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.A.; Hitotsuyanagi, Y.; Mansour, E.S.S.; Ahmed, A.F.; Gedara, S.; Fukaya, H.; Takeya, K. Cucurbitacins from Bryonia cretica. Phytochem. Lett. 2010, 3, 117–121. [Google Scholar] [CrossRef]
- Matsuda, H.; Nakashima, S.; Abdel-Halim, O.B.; Morikawa, T.; Yoshikawa, M. Cucurbitane-type triterpenes with anti-proliferative effects on U937 cells from an egyptian natural medicine, Bryonia cretica: Structures of new triterpene glycosides, bryoniaosides A and B. Chem. Pharm. Bull. 2010, 58, 747–751. [Google Scholar] [CrossRef][Green Version]
- Dantas, I.N.F.; Gadelha, G.C.M.; Chaves, D.C.; Monte, F.J.Q.; Pessoa, C.; de Moraes, M.O.; Costa-Lotufo, L.V. Studies on the Cytotoxicity of Cucurbitacins Isolated from Cayaponia racemosa (Cucurbitaceae). Z. Naturforsch. 2006, 61, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, H.; Singh, T.; Reynolds, W.F.; McLean, S. Isolation and 13C-NMR Assignments of Cucurbitacins from Cayaponia Angustiloba, Cayaponia racemosa, and Guranias ubumbellata. J. Nat. Prod. 1990, 53, 1600–1605. [Google Scholar] [CrossRef]
- Recio, M.C.; Prieto, M.; Bonucelli, M.; Orsi, C.; Manez, S.; Giner, R.M.; Cerda-Nicolas, M.; Rios, J.L. Anti-inflammatory activity of two cucurbitacins isolated from Cayaponia tayuya roots. Planta Med. 2004, 70, 414–420. [Google Scholar] [CrossRef]
- Gamlath, C.B.; Gunatilaka, A.A.L.; Alvi, K.A.; ur Rahman, A.; Balasubramaniam, S. Cucurbitacins of Colocynthis vulgaris. Phytochemistry 1988, 27, 3225–3229. [Google Scholar] [CrossRef]
- Tannin-Spitz, T.; Grossman, S.; Dovrat, S.; Gottlieb, H.E.; Bergman, M. Growth inhibitory activity of cucurbitacin glucosides isolated from Citrullus colocynthis on human breast cancer cells. Biochem. Pharmacol. 2007, 73, 56–67. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Morikawa, T.; Kobayashi, H.; Nakamura, A.; Matsuhira, K.; Nakamura, S.; Matsuda, H. Bioactive saponins and glycosides. XXVII. Structures of new cucurbitane-type triterpene glycosides and antiallergic constituents from Citrullus colocynthis. Chem. Pharm. Bull. 2007, 55, 428–434. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, G.; Chen, X.; Chen, S.X.; Gan, L.S.; Yuan, T. Colocynthenins A-D, Ring-A seco-Cucurbitane Triterpenoids from the Fruits of Citrullus colocynthis. J. Nat. Prod. 2018, 81, 2115–2119. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Qiang, S.; Lou, L.; Zhao, W. Cucurbitane-type triterpenoids from the stems of Cucumis melo. J. Nat. Prod. 2009, 72, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Afifi, M.S.; Ross, S.A.; ElSohly, M.A.; Naeem, Z.E.; Halaweish, F.T. Cucurbitacins of Cucumis prophetarum and Cucumis prophetarum. J. Chem. Ecol. 1999, 25, 847–859. [Google Scholar] [CrossRef]
- Qing, Z.; Shi, Y.; Han, L.; Li, P.; Zha, Z.; Liu, C.; Liu, X.; Huang, P.; Liu, Y.; Tang, Q.; et al. Identification of seven undescribed cucurbitacins in Cucumis sativus (cucumber) and their cytotoxic activity. Phytochemistry 2022, 197, 113123. [Google Scholar] [CrossRef]
- Halaweish, F.T.; Tallamy, D.W. A new cucurbitacin profile for Cucurbita andreana: A candidate for cucurbitacin tissue culture. J. Chem. Ecol. 1993, 19, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Hutt, T.F.; Herrington, M.E. The determination of bitter principles in zucchinis. J. Sci. Food Agric. 1985, 36, 1107–1112. [Google Scholar] [CrossRef]
- Greige-Gerges, H.; Khalil, R.A.; Mansour, E.A.; Magdalou, J.; Chahine, R.; Ouaini, N. Cucurbitacins from Ecballium elaterium juice increase the binding of bilirubin and ibuprofen to albumin in human plasma. Chem. Biol. Interact. 2007, 169, 53–62. [Google Scholar] [CrossRef]
- Seger, C.; Sturm, S.; Haslinger, E.; Stuppner, H. NMR Signal Assignment of 22-Deoxocucurbitacin D and Cucurbitacin D from Ecballium elaterium L. (Cucurbitaceae). Monatsh. Chem. 2005, 136, 1645–1649. [Google Scholar] [CrossRef]
- Hano, Y.; Shi, Y.Q.; Nomura, T.; Yang, P.Q.; Chang, W.J. Two acetogenins from Hemsleya ellipsoidea. Phytochemistry 1997, 46, 1447–1449. [Google Scholar] [CrossRef]
- Torres-Moreno, H.; Marcotullio, M.C.; Velazquez, C.; Ianni, F.; Garibay-Escobar, A.; Robles-Zepeda, R.E. Cucurbitacin IIb, a steroidal triterpene from Ibervillea sonorae induces antiproliferative and apoptotic effects on cervical and lung cancer cells. Steroids 2020, 157, 108597. [Google Scholar] [CrossRef]
- Attar, U.A.; Ghane, S.G. Optimized extraction of anti-cancer compound—Cucurbitacin I and LC–MS identification of major metabolites from wild Bottle gourd (Lagenaria siceraria (Molina) Standl.). S. Afr. J. Bot. 2018, 119, 181–187. [Google Scholar] [CrossRef]
- Kumar, S.; Sharma, K.; Sahai, M.; Maurya, R. A New Cucurbitacin Glucoside from Luffa graveolense. Chem. Nat. Compd. 2019, 55, 490–492. [Google Scholar] [CrossRef]
- Liu, J.Q.; Chen, J.C.; Wang, C.F.; Qiu, M.H. New Cucurbitane Triterpenoids and Steroidal Glycoside from Momordica charantia. Molecules 2009, 14, 4804–4813. [Google Scholar] [CrossRef] [PubMed]
- Aguiniga-Sanchez, I.; Cadena-Iniguez, J.; Santiago-Osorio, E.; Gomez-Garcia, G.; Mendoza-Nunez, V.M.; Rosado-Perez, J.; Ruiz-Ramos, M.; Cisneros-Solano, V.M.; Ledesma-Martinez, E.; Delgado-Bordonave, A.D. Chemical analyses and in vitro and in vivo toxicity of fruit methanol extract of Sechium edule var. nigrum spinosum. Pharm. Biol. 2017, 55, 1638–1645. [Google Scholar] [CrossRef]
- Kongtun, S.; Juratchariyakul, W.; Kummalue, T.; Tan-ariya, P.; Kunnachak, S.; Frahm, A.W. Cytotoxic properties of root extract and fruit juice of Trichosanthes cucumerina. Planta Med. 2009, 75, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.Y.; Lee, S.H.; Choi, S.U.; Lee, C.O.; No, Z.; Ahn, J.W. Antitumor activity of Trichosanthes kirilowii. Arch. Pharm. Res. 1994, 17, 348–353. [Google Scholar] [CrossRef]
- Oh, H.; Mun, Y.J.; Im, S.J.; Lee, S.Y.; Song, H.J.; Lee, H.S.; Woo, W.H. Cucurbitacins from Trichosanthes kirilowii as the inhibitory components on tyrosinase activity and melanin synthesis of B16/F10 melanoma cells. Planta Med. 2002, 68, 832–833. [Google Scholar] [CrossRef]
- Kanchanapoom, T.; Kasai, R.; Yamasaki, K. Cucurbitane, hexanorcucurbitane and octanorcucurbitane glycosides from fruits of Trichosanthes tricuspidata. Phytochemistry 2002, 59, 215–228. [Google Scholar] [CrossRef]
- Frei, B.; Heinrich, M.; Herrmann, D.; Orjala, J.E.; Schmitt, J.; Sticher, O. Phytochemical and biological investigation of Begonia heracleifolia. Planta Med. 1998, 64, 385–386. [Google Scholar] [CrossRef]
- Wu, P.L.; Lin, F.W.; Wu, T.S.; Kuoh, C.H.; Lee, K.H.; Lee, S.J. Cytotoxic and anti-HIV principles from the rhizomes of Begonia nantoensis. Chem. Pharm. Bull. 2004, 52, 345–349. [Google Scholar] [CrossRef]
- Sachdev-Gupta, K.; Radke, C.D.; Renwick, J.A.A. Antifeedant activity of cucurbitacins from Iberis amara against larvae of Pieris rapae. Phytochemistry 1993, 33, 1385–1388. [Google Scholar] [CrossRef]
- Curtis, P.J.; Meade, P.M. Cuburbitacins from the Cruciferae. Phytochemistry 1971, 10, 3081–3083. [Google Scholar] [CrossRef]
- Sarker, S.D.; Whiting, P.; Lafont, R.; Girault, J.P.; Dinan, L. Cucurbitacin D from Cercidiphyllum japonicum. Biochem. Syst. Ecol. 1997, 25, 79–80. [Google Scholar] [CrossRef]
- Sasamori, H.; Reddy, K.S.; Kirkup, M.P.; Shabanowitz, J.; Lynn, D.G.; Hecht, S.M.; Woode, K.A.; Bryan, R.F.; Campbell, J.; Lynn, W.S.; et al. New cytotoxic principles from Datisca glomerata. J. Chem. Soc. Perkin Trans. 1983, 1, 1333–1347. [Google Scholar] [CrossRef]
- Reddy, K.S.; Amonkar, A.J.; McCloud, T.G.; Chang, C.J.; Cassady, J.M. Spinosides A and B. Two cytotoxic cucurbitacin glycosides from Desfontainia spinosa. Phytochemistry 1988, 27, 3781–3785. [Google Scholar] [CrossRef]
- Oberlies, N.H.; Burgess, J.P.; Navarro, H.A.; Pinos, R.E.; Soejarto, D.D.; Farnsworth, N.R.; Kinghorn, A.D.; Wani, M.C.; Wall, M.E. Bioactive constituents of the roots of Licania intrapetiolaris. J. Nat. Prod. 2001, 64, 497–501. [Google Scholar] [CrossRef]
- Meng, D.; Qiang, S.; Lou, L.; Zhao, W. Cytotoxic cucurbitane-type triterpenoids from Elaeocarpus hainanensis. Planta Med. 2008, 74, 1741–1744. [Google Scholar] [CrossRef]
- Ito, A.; Chai, H.B.; Lee, D.; Kardono, L.B.S.; Riswan, S.; Farnsworth, N.R.; Cordell, G.A.; Pezzuto, J.M.; Kinghorn, A.D. Ellagic acid derivatives and cytotoxic cucurbitacins from Elaeocarpus mastersii. Phytochemistry 2002, 61, 171–174. [Google Scholar] [CrossRef]
- Wang, Y.J.; Yang, J.; Li, X.N.; Bai, H.; Luo, J.F.; He, Z.R.; Wang, Y.H. Cucurbitane-type triterpenoids from the branches and leaves of Elaeocarpus sylvestris. Phytochem. Lett. 2022, 51, 39–45. [Google Scholar] [CrossRef]
- Rodriguez, N.; Vasquez, Y.; Hussein, A.A.; Coley, P.D.; Solis, P.N.; Gupta, M.P. Cytotoxic cucurbitacin constituents from Sloanea zuliaensis. J. Nat. Prod. 2003, 66, 1515–1516. [Google Scholar] [CrossRef]
- Liu, M.T.; Lin, S.; Wang, Y.H.; He, W.Y.; Li, S.; Wang, S.J.; Yang, Y.C.; Shi, J.G. Two novel glycosidic triterpene alkaloids from the stem barks of Machilus yaoshansis. Org. Lett. 2007, 9, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Gan, M.; Liu, M.; Liu, B.; Lin, S.; Zhang, Y.; Zi, J.; Song, W.; Ye, F.; Chen, X.; Shi, J. Cucurbitane glucosides from the root of Machilus yaoshansis. J. Nat. Prod. 2011, 74, 2431–2437. [Google Scholar] [CrossRef] [PubMed]
- Bean, M.F.; Antoun, M.; Abramson, D.; Chang, C.J.; McLaughlin, J.L.; Cassady, J.M. Cucurbitacin B and Isocucurbitacin B: Cytotoxic Components of Helicteres isora. J. Nat. Prod. 1985, 48, 500. [Google Scholar] [CrossRef]
- Bhandari, P.; Kumar, N.; Singh, B.; Kaul, V.K. Cucurbitacins from Bacopa monnieri. Phytochemistry 2007, 68, 1248–1254. [Google Scholar] [CrossRef]
- Musza, L.L.; Speight, P.; McElhiney, S.; Barrow, C.J.; Gillum, A.M.; Cooper, R.; Killar, L.M. Cucurbitacins, cell adhesion inhibitors from Conobea scoparioides. J. Nat. Prod. 1994, 57, 1498–1502. [Google Scholar] [CrossRef]
- Kim, I.H.; Uchiyama, N.; Kawahara, N.; Goda, Y. Iridoid glycosides and cucurbitacin glycoside from Neopicrorhiza scrophulariiflora. Phytochemistry 2006, 67, 2691–2696. [Google Scholar] [CrossRef]
- Huang, Y.; De Bruyne, T.; Apers, S.; Ma, Y.; Claeys, M.; Vanden Berghe, D.; Pieters, L.; Vlietinck, A. Complement-inhibiting cucurbitacin glycosides from Picria fel-terrae. J. Nat. Prod. 1998, 61, 757–761. [Google Scholar] [CrossRef]
- Stuppner, H.; Wagner, H. New cucurbitacin glycosides from Picrorhiza kurrooa. Planta Med. 1989, 55, 559–563. [Google Scholar] [CrossRef]
- Stuppner, H.; Moller, E.P. Cucurbitacins with unusual side chains from Picrorhiza kurroa. Phytochemistry 1993, 33, 1139–1145. [Google Scholar] [CrossRef]
- Wang, H.; Ye, W.C.; Zhao, S.X. Cucurbitacin glycosides and the monoterpene jiofuran from Picrorhiza scrophulariiflora. Biochem. Syst. Ecol. 2004, 32, 87–89. [Google Scholar] [CrossRef]
- Arisawa, M.; Pezzuto, J.M.; Kinghorn, A.D.; Cordell, G.A.; Farnsworth, N.R. Plant Anticancer Agents XXX: Cucurbitacins from Ipomopsis aggregata (Polemoniaceae). J. Pharm. Sci. 1984, 73, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Hagiwara, K.; Iguchi, K.; Suzuki, S.; Hsu, H.Y. Isolation and structures of arvenins from Anagallis arvensis L. (Primulaceae). New cucurbitacin glucosides. Chem. Pharm. Bull. 1978, 26, 3107–3112. [Google Scholar] [CrossRef] [PubMed]
- Munoz, O.; Estevez-Braun, A.M.; Ravelo, A.G.; Gonzalez, A.G. Cucurbitacin F in seeds of Kageneckia angustifolia (Rosaceae). Z. Naturforsch. 2002, 57, 208–209. [Google Scholar] [CrossRef]
- Munoz, O.; Delporte, C.; Backhouse, N.; Erazo, S.; Negrete, R.; Maldonado, S.; Lopez-Perez, J.L.; San Feliciano, A. A new cucurbitacin glycoside from Kageneckia oblonga (Rosaceae). Z. Naturforsch. 2000, 55, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Maloney, K.N.; Fujita, M.; Eggert, U.S.; Schroeder, F.C.; Field, C.M.; Mitchison, T.J.; Clardy, J. Actin-Aggregating Cucurbitacins from Physocarpus capitatus. J. Nat. Prod. 2008, 71, 1927–1929. [Google Scholar] [CrossRef]
- Sarker, S.D.; Whiting, P.; Sik, V.; Dinan, L. Ecdysteroid antagonists (cucurbitacins) from Physocarpus opulifolius (Rosaceae). Phytochemistry 1999, 50, 1123–1128. [Google Scholar] [CrossRef]
- Konoshima, T.; Takasaki, M.; Kozuka, M.; Haruna, M.; Ito, K.; Estes, J.R.; Lee, K.H. Constituents of rosaceous plants. I. Structure of new triterpenoids from Cowania mexicana. Chem. Pharm. Bull. 1993, 41, 1612–1615. [Google Scholar] [CrossRef][Green Version]
- Kim, D.K.; Choi, S.H.; Lee, J.O.; Ryu, S.Y.; Park, D.K.; Shin, D.H.; Jung, J.H.; Pyo, S.K.; Lee, K.R.; Zee, O.P. Cytotoxic constituents of Sorbaria sorbifolia var. stellipila. Arch. Pharm. Res. 1997, 20, 85–87. [Google Scholar] [CrossRef]
- Guerrero-Analco, J.A.; Hersch-Martinez, P.; Pedraza-Chaverri, J.; Navarrete, A.; Mata, R. Antihyperglycemic effect of constituents from Hintonia standleyana in streptozotocin-induced diabetic rats. Planta Med. 2005, 71, 1099–1105. [Google Scholar] [CrossRef]
- Mata, R.; Rios, L.; Rayo Camacho, D.; Reguero, M.T.; Lorence, D. Triterpenes from Cigarrilla mexicana. Phytochemistry 1988, 27, 1887–1889. [Google Scholar] [CrossRef]
- Mata, R.; Castaneda, P.; Camacho, M.; Delgado, G. Chemical studies on Mexican plants used in traditional medicine, V. Cucurbitacin glucosides from Cigarrilla mexicana. J. Nat. Prod. 1988, 51, 836–839. [Google Scholar] [CrossRef]
- Sun, J.; Xia, F.; Wang, S.; Wang, K.Y.; Chen, J.M.; Tu, P.F. Structural elucidation of two new megastigmane glycosides from the leaves of Aquilaria sinensis. Chin. J. Nat. Med. 2015, 13, 290–294. [Google Scholar] [CrossRef]
- Fuller, R.W.; Cardellina, J.H.; Cragg, G.M.; Boyd, M.R. Cucurbitacins: Differential cytotoxicity, dereplication and first isolation from Gonystylus keithii. J. Nat. Prod. 1994, 57, 1442–1445. [Google Scholar] [CrossRef]
- Bashir, S.S.; Hussain, A.; Hussain, S.J.; Wani, O.A.; Nabi, S.Z.; Dar, N.A.; Baloch, F.S.; Mansoor, S. Plant drought stress tolerance: Understanding its physiological, biochemical and molecular mechanisms. Biotechnol. Biotechnol. Equip. 2021, 35, 1912–1925. [Google Scholar] [CrossRef]
- Aparna; Skarzyńska, A.; Pląder, W.; Pawełkowicz, M. Impact of Climate Change on Regulation of Genes Involved in Sex Determination and Fruit Production in Cucumber. Plants 2023, 12, 2651. [Google Scholar] [CrossRef]
- Shang, Y.; Ma, Y.; Zhou, Y.; Zhang, H.; Duan, L.; Chen, H.; Zeng, J.; Zhou, Q.; Wang, S.; Gu, W.; et al. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Kano, Y.; Goto, H. Relationship between the occurrence of bitter fruit in cucumber (Cucumis sativus L.) and the contents of total nitrogen, amino acid nitrogen, protein and HMG-CoA reductase activity. Sci. Hortic. 2003, 98, 1–8. [Google Scholar] [CrossRef]
- Zhao, G.; Wang, M.; Luo, C.; Li, J.; Gong, H.; Zheng, X.; Liu, X.; Luo, J.; Wu, H. Metabolome and Transcriptome Analyses of Cucurbitacin Biosynthesis in Luffa (Luffa acutangula). Front. Plant. Sci. 2022, 13, 886870. [Google Scholar] [CrossRef] [PubMed]
- Mashilo, J.; Odindo, A.O.; Shimelis, H.A.; Musenge, P.; Tesfay, S.Z.; Magwaza, L.S. Photosynthetic response of bottle gourd [Lagenaria siceraria (Molina) Standl.] to drought stress: Relationship between cucurbitacins accumulation and drought tolerance. Sci. Hortic. 2018, 231, 133–143. [Google Scholar] [CrossRef]
- Wang, W.D.; Liu, Y.; Su, Y.; Xiong, X.Z.; Shang, D.; Xu, J.J.; Liu, H.J. Antitumor and apoptotic effects of cucurbitacin a in A-549 lung carcinoma cells is mediated via G2/M cell cycle arrest and M-TOR/PI3K/Akt signalling pathway. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 75–82. [Google Scholar] [CrossRef]
- Liu, J.Y.; Liu, X.; Ma, W.; Kou, W.; Li, C.L.; Zhao, J. Anticancer activity of cucurbitacin-A in ovarian cancer cell line SKOV3 involves cell cycle arrest, apoptosis and inhibition of mTOR/PI3K/Akt signaling pathway. J. BUON. 2018, 23, 124–128. [Google Scholar] [PubMed]
- Luo, W.W.; Zhao, W.W.; Lu, J.J.; Wang, Y.T.; Chen, X.P. Cucurbitacin B suppresses metastasis mediated by reactive oxygen species (ROS) via focal adhesion kinase (FAK) in breast cancer MDA-MB-231 cells. Chin. J. Nat. Med. 2018, 16, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, G.; Bao, J.; Hao, W.; Lu, J.; Chen, X. Cucurbitacin B induced ATM-mediated DNA damage causes G2/M cell cycle arrest in a ROS-dependent manner. PLoS ONE 2014, 9, e88140. [Google Scholar] [CrossRef] [PubMed]
- Aribi, A.; Gery, S.; Lee, D.H.; Thoennissen, N.H.; Thoennissen, G.B.; Alvarez, R.; Ho, Q.; Doan, N.B.; Chan, K.T.; Toh, M.; et al. The triterpenoid cucurbitacin B augments the antiproliferative activity of chemotherapy in human breast cancer. Int. J. Cancer 2013, 132, 2730–2737. [Google Scholar] [CrossRef]
- Zhang, M.; Yin, L.; Yang, S.; Hong, J.; Chen, C.; Han, D.; Hou, Y.; Zhang, B.; Huang, L.; Zhang, A.; et al. Abstract 5728: The synergistic effect of Cucurbitacin B and radiation treatment. Cancer Res. 2012, 72, 5728. [Google Scholar] [CrossRef]
- Bakar, F. Cucurbitacin B Enhances the Anticancer Effect of Imatinib Mesylate Through Inhibition of MMP-2 Expression in MCF-7 and SW480 Tumor Cell Lines. Anticancer. Agents Med. Chem. 2016, 16, 747–754. [Google Scholar] [CrossRef]
- Kaewmeesri, P.; Pocasap, P.; Kukongviriyapan, V.; Prawan, A.; Kongpetch, S.; Senggunprai, L. Anti-metastatic Potential of Natural Triterpenoid Cucurbitacin B Against Cholangiocarcinoma Cells by Targeting Src Protein. Integr. Cancer Ther. 2022, 21, 15347354221124861. [Google Scholar] [CrossRef]
- Yar Saglam, A.S.; Alp, E.; Elmazoglu, Z.; Menevse, S. Treatment with cucurbitacin B alone and in combination with gefitinib induces cell cycle inhibition and apoptosis via EGFR and JAK/STAT pathway in human colorectal cancer cell lines. Hum. Exp. Toxicol. 2016, 35, 526–543. [Google Scholar] [CrossRef]
- Liu, X.; Duan, C.; Ji, J.; Zhang, T.; Yuan, X.; Zhang, Y.; Ma, W.; Yang, J.; Yang, L.; Jiang, Z.; et al. Cucurbitacin B induces autophagy and apoptosis by suppressing CIP2A/PP2A/mTORC1 signaling axis in human cisplatin resistant gastric cancer cells. Oncol. Rep. 2017, 38, 271–278. [Google Scholar] [CrossRef]
- Yin, D.; Wakimoto, N.; Xing, H.T.; Lu, D.; Huynh, T.; Wang, X.; Black, K.L.; Koeffler, H.P. Cucurbitacin B markedly inhibits growth and rapidly affects the cytoskeleton in glioblastoma multiforme. Int. J. Cancer 2008, 123, 1364–1375. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, J.; Zhou, J.; Huang, Z.; Hu, H.; Qiao, M.; Zhao, X.; Chen, D. Synergistic effect of cucurbitacin B in combination with curcumin via enhancing apoptosis induction and reversing multidrug resistance in human hepatoma cells. Eur. J. Pharmacol. 2015, 768, 28–40. [Google Scholar] [CrossRef]
- Touihri-Barakati, I.; Kallech-Ziri, O.; Ayadi, W.; Kovacic, H.; Hanchi, B.; Hosni, K.; Luis, J. Cucurbitacin B purifed from Ecballium elaterium (L.) A. Rich from Tunisia inhibits alpha5beta1 integrin-mediated adhesion, migration, proliferation of human glioblastoma cell line and angiogenesis. Eur. J. Pharmacol. 2017, 797, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, M.; Zhang, H.; Sun, C.; Yang, X.; Deng, Y.; Ji, W. Combined antitumor activity of cucurbitacin B and docetaxel in laryngeal cancer. Eur. J. Pharmacol. 2008, 587, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Li, C.; Cao, L.; Zhang, C.H.; Zhang, Z.H. Cucurbitacin B regulates lung cancer cell proliferation and apoptosis via inhibiting the IL-6/STAT3 pathway through the lncRNA XIST/miRlet-7c axis. Pharmaceutic. Biol. 2022, 60, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Fan, Q.; Liang, X.; Han, S.; He, J.; Wang, Q.Q.; Gao, H.; Feng, Y.; Yang, S. Cucurbitacin B inhibits TGF-β1-induced epithelial-mesenchymal transition (EMT) in NSCLC through regulating ROS and PI3K/Akt/mTOR pathways. Chin. Med. 2022, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zheng, L.; Tang, H.; Wang, W.; Lin, Y. Cucurbitacin B enhances apoptosis in geftinib resistant non-small cell lung cancer by modulating the miR-17-5p/STAT3 axis. Mol. Med. Rep. 2021, 24, 710. [Google Scholar] [CrossRef]
- Ding, X.; Chi, J.; Yang, X.; Hao, J.; Liu, C.; Zhu, C.; Wang, X.; Liu, X.; Niu, Y.; Ji, W.; et al. Cucurbitacin B synergistically enhances the apoptosis-inducing effect of arsenic trioxide by inhibiting STAT3 phosphorylation in lymphoma Ramos cells. Leuk. Lymphoma 2017, 58, 2439–2451. [Google Scholar] [CrossRef]
- Wei, J.; Chen, X.; Li, Y.; Li, R.; Bao, K.; Liao, L.; Xie, Y.; Yang, T.; Zhu, J.; Mao, F.; et al. Cucurbitacin B-induced G2/M cell cycle arrest of conjunctival melanoma cells mediated by GRP78-FOXM1-KIF20A pathway. Acta Pharmaceut. Sin. B 2022, 12, 3861–3876. [Google Scholar] [CrossRef]
- Zheng, Q.; Liu, Y.; Liu, W.; Ma, F.; Zhou, Y.; Chen, M.; Chang, J.; Wang, Y.; Yang, G.; He, G. Cucurbitacin B inhibits growth and induces apoptosis through the JAK2/STAT3 and MAPK pathways in SHSY5Y human neuroblastoma cells. Mol. Med. Rep. 2014, 10, 89–94. [Google Scholar] [CrossRef]
- Zhang, Z.R.; Gao, M.X.; Yang, K. Cucurbitacin B inhibits cell proliferation and induces apoptosis in human osteosarcoma cells via modulation of the JAK2/STAT3 and MAPK pathways. Expert. Therap. Med. 2017, 14, 805–812. [Google Scholar] [CrossRef]
- Lee, D.H.; Thoennissen, N.H.; Goff, C.; Iwanski, G.B.; Forscher, C.; Doan, N.B.; Said, J.W.; Koeffler, H.P. Synergistic effect of low-dose cucurbitacin B and low-dose methotrexate for treatment of human osteosarcoma. Cancer Lett. 2011, 306, 161–170. [Google Scholar] [CrossRef] [PubMed]
- El-Senduny, F.F.; Badria, F.A.; El-Waseef, A.M.; Chauhan, S.C.; Halaweish, F. Approach for chemosensitization of cisplatin-resistant ovarian cancer by cucurbitacin B. Tumor Biol. 2016, 37, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhao, T.; Ma, L.; Liang, M.; Guo, Y.J.; Zhao, L.M. Cucurbitacin B and SCH772984 exhibit synergistic anti-pancreatic cancer activities by suppressing EGFR, PI3K/Akt/mTOR, STAT3 and ERK signaling. Oncotarget 2017, 8, 103167. [Google Scholar] [CrossRef] [PubMed]
- Thoennissen, N.H.; Iwanski, G.B.; Doan, N.B.; Okamoto, R.; Lin, P.; Abbassi, S.; Song, J.H.; Yin, D.; Toh, M.; Xie, W.D.; et al. Cucurbitacin B Induces Apoptosis by Inhibition of the JAK/STAT Pathway and Potentiates Antiproliferative Effects of Gemcitabine on Pancreatic Cancer Cells. Cancer Res. 2009, 69, 5876–5884. [Google Scholar] [CrossRef] [PubMed]
- Iwanski, G.B.; Lee, D.H.; En-Gal, S.; Doan, N.B.; Castor, B.; Vogt, M.; Toh, M.; Bokemeyer, C.; Said, J.W.; Thoennissen, N.H.; et al. Cucurbitacin B, a novel in vivo potentiator of gemcitabine with low toxicity in the treatment of pancreatic cancer. Br. J. Pharmacol. 2010, 160, 998–1007. [Google Scholar] [CrossRef]
- Tao, B.; Wang, D.; Yang, S.; Liu, Y.; Wu, H.; Li, Z.; Chang, L.; Yang, Z.; Liu, W. Cucurbitacin B Inhibits Cell Proliferation by Regulating X-Inactive Specific Transcript Expression in Tongue Cancer. Front. Oncol. 2021, 11, 651648. [Google Scholar] [CrossRef]
- Ku, J.M.; Kim, S.R.; Hong, S.H.; Choi, H.S.; Seo, H.S.; Shin, Y.C.; Ko, S.G. Cucurbitacin D induces cell cycle arrest and apoptosis by inhibiting STAT3 and NF-κB signaling in doxorubicin-resistant human breast carcinoma (MCF7/ADR) cells. Mol. Cell Biochem. 2015, 409, 33–43. [Google Scholar] [CrossRef]
- Sikander, M.; Hafeez, B.B.; Malik, S.; Alsayari, A.; Halaweish, F.T.; Yallapu, M.M.; Chauhan, S.C.; Jaggi, M. Cucurbitacin D exhibits potent anti-cancer activity in cervical cancer. Sci. Rep. 2016, 6, 36594. [Google Scholar] [CrossRef]
- Sikander, M.; Malik, S.; Khan, S.; Kumari, S.; Chauhan, N.; Khan, P.; Halaweish, F.T.; Chauhan, B.; Yallapu, M.M.; Jaggi, M.; et al. Novel Mechanistic Insight into the Anticancer Activity of Cucurbitacin D against Pancreatic Cancer (Cuc D Attenuates Pancreatic Cancer). Cells 2019, 9, 103. [Google Scholar] [CrossRef]
- Sikander, M.; Malik, S.; Hafeez, B.B.; Mandil, H.; Halaweish, F.T.; Jaggi, M.; Chauhan, S.C. Abstract 2934: Cucurbitacin D enhances the therapeutic efficacy of docetaxel via targeting cancer stem cells and miR-145. Cancer Res. 2018, 78, 2934. [Google Scholar] [CrossRef]
- Jacquot, C.; Rousseau, B.; Carbonnelle, D.; Chinou, I.; Malleter, M.; Tomasoni, C.; Roussakis, C. Cucurbitacin-D-induced CDK1 mRNA up-regulation causes proliferation arrest of a non-small cell lung carcinoma cell line (NSCLC-N6). Anticancer. Res. 2014, 34, 4797–4806. [Google Scholar] [PubMed]
- Cheng, A.C.; Hsu, Y.C.; Tsai, C.C. The efects of cucurbitacin E on GADD45βtrigger G2/M arrest and JNK-independent pathway in brain cancer cells. J. Cell Mol. Med. 2019, 23, 3512–3519. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.Y.; Wu, Z.D.; Zhang, T.H.; Zhang, J.; Wei, Z.Y. In vitro antitumor efect of cucurbitacin E on human lung cancer cell line and its molecular mechanism. Chin. J. Nat. Med. 2020, 18, 483–490. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Chen, M.J.; Huang, T.Y. Inducement of mitosis delay by cucurbitacin E, a novel tetracyclic triterpene from climbing stem of Cucumis melo L., through GADD45gamma in human brain malignant glioma (GBM) 8401 cells. Cell Death Dis. 2014, 5, e1087. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Wang, L.L.; Xu, Q.; Liu, W.G.; Jin, H.C.; Mao, W.M.; Wang, X.; Wang, X. Growth inhibitory effect of cucurbitacin E on breast cancer cells. Int. J. Clin. Exp. Pathol. 2013, 6, 1799–1805. [Google Scholar]
- Si, W.; Lyu, J.; Liu, Z.; Wang, C.; Huang, J.; Jiang, L.; Ma, T. Cucurbitacin E inhibits cellular proliferation and enhances the chemo-response in gastric cancer by suppressing AKt activation. J. Cancer 2019, 10, 5843–5851. [Google Scholar] [CrossRef] [PubMed]
- Eyol, E.; Tanriverdi, Z.; Karakus, F.; Yilmaz, K.; Unuvar, S. Synergistic Anti-proliferative Effects of Cucurbitacin I and Irinotecan on Human Colorectal Cancer Cell Lines. Clin. Exp. Pharmacol. 2016, 6, 5. [Google Scholar] [CrossRef]
- Song, J.M.; Liu, H.X.; Li, Z.; Yang, C.; Wang, C.J. Cucurbitacin I inhibits cell migration and invasion and enhances chemosensitivity in colon cancer. Oncol. Rep. 2015, 33, 1867–1871. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, J.H.P.; Kim, J.K. Cucurbitacin-I, a natural cell-permeable triterpenoid isolated from Cucurbitaceae, exerts potent anticancer effect in colon cancer. Chem. Biol. Interact. 2014, 219, 1–8. [Google Scholar] [CrossRef]
- Zhu, X.; Huang, H.; Zhang, J.; Liu, H.; Ao, R.; Xiao, M.; Wu, Y. The anticancer effects of Cucurbitacin I inhibited cell growth of human non small cell lung cancer through PI3K/AKT/p70S6K pathway. Mol. Med. Rep. 2018, 17, 2750–2756. [Google Scholar] [CrossRef]
- Ni, Y.; Wu, S.; Wang, X.; Zhu, G.; Chen, X.; Ding, Y.; Jiang, W. Cucurbitacin I induces pro-death autophagy in A549 cells via the ERK-mTOR-STAT3 signaling pathway. J. Cell. Biochem. 2018, 119, 6104–6112. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, H.; Li, R.; Xin, J.; Wu, S.; Lan, J.; Xue, K.; Li, X.; Zuo, C.; Jiang, W.; et al. Cucurbitacin I induces cancer cell death through the endoplasmic reticulum stress pathway. J. Cell. Biochem. 2019, 120, 2391–2403. [Google Scholar] [CrossRef] [PubMed]
- Sadzuka, Y.; Fujiki, S.; Itai, S. Enhancement of doxorubicin-induced antitumor activity and reduction of adverse reactions by cucurbitacin I. Food Res. Int. 2012, 47, 64–69. [Google Scholar] [CrossRef]
- Xu, D.; Shen, H.; Tian, M.; Chen, W.; Zhang, X. Cucurbitacin I inhibits the proliferation of pancreatic cancer through the JAK2/STAT3 signalling pathway in vivo and in vitro. J. Cancer 2022, 13, 2050–2060. [Google Scholar] [CrossRef]
- Zhang, J.; Song, Y.; Liang, Y.; Zou, H.; Zuo, P.; Yan, M.; Jing, S.; Li, T.; Wang, Y.; Li, D.; et al. Cucurbitacin IIa interferes with EGFR-MAPK signaling pathway leads to proliferation inhibition in A549cells. Food. Chem. Toxicol. 2019, 132, 110654. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, T.; Ren, L.; Jing, S.; Li, Z.; Zuo, P.; Li, T.; Wang, Y.; Zhang, J.; Wei, Z. Cucurbitacin IIb induces apoptosis and cell cycle arrest through regulating EGFR/MAPK pathway. Environ. Toxicol. Pharmacol. 2021, 81, 103542. [Google Scholar] [CrossRef]
- Arjaibi, H.M.; Ahmed, M.S.; Halaweish, F.T. Mechanistic investigation of hepato-protective potential for cucurbitacins. Med. Chem. Res. 2017, 26, 1567–1573. [Google Scholar] [CrossRef]
- Zhong, H.; Huang, Y.; Deng, X.; Liu, M.; Luo, W. Cucurbitacin B supplementation reduces inflammatory responses and alveolar bone loss via regulating MPO, COX-2 and RANK/RANKL/OPG signals in a rodent model of ligature-induced periodontitis. J. King Saud. Univ.—Sci. 2020, 32, 1889–1895. [Google Scholar] [CrossRef]
- Shawkey, A.M.; Rabeh, M.A.; Abdellatif, A.O. Biofunctional molecules from Citrullus colocynthis: An HPLC/MS analysis in correlation to antimicrobial and anticancer activities. Advan. Life Sci. Tech. 2014, 17, 51–61. [Google Scholar]
- Kapoor, N.; Ghorai, S.M.; Kushwaha, P.K.; Shukla, R.; Aggarwal, C.; Bandichhor, R. Plausible mechanisms explaining the role of cucurbitacins as potential therapeutic drugs against coronavirus 2019. Inform. Med. Unlocked 2020, 21, 100484. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, Z.; Wu, Q.Q.; Jiang, X.H.; Yuan, Y.; Chang, W.; Bian, Z.Y.; Zhu, J.X.; Tang, Q.Z. Cucurbitacin B Protects Against Pressure Overload Induced Cardiac Hypertrophy. J. Cell. Biochem. 2017, 118, 3899–3910. [Google Scholar] [CrossRef] [PubMed]
- Hernández Navia, S.E.; Figueroa-Hernández, J.L.; Rodriguez-Zavala, J.S.; Rodriguez-Sosa, M.; Martinez-Vasquez, M. Anti-Diabetic Effects of Cucurbitacins from Ibervillea lindheimeri on Induced Mouse Diabetes. J. Chem. 2022, 2022, 3379557. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, B.; Sharma, U.; Parashar, G.; Parashar, N.C.; Rani, I.; Ramniwas, S.; Kaur, S.; Haque, S.; Tuli, H.S. Apoptotic and antimetastatic effect of cucurbitacins in cancer: Recent trends and advancement. Naunyn Schmiedebergs Arch. Pharmacol. 2023, 396, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Yung, M.M.H.; Ross, F.A.; Hardie, D.G.; Leung, T.H.; Zhan, J.; Ngan, H.Y.S.; Chan, D.W. Bitter melon (Momordica charantia) extract functions as a natural AMPK activator and synergistically enhances cisplatin cytotoxicity in ovarian cancer cells. Integr. Cancer Ther. 2016, 15, 376–389. [Google Scholar] [CrossRef]
- Gou, M.; Wei, X.; Men, K.; Wang, B.; Luo, F.; Zhao, X.; Wei, Y.Q.; Qian, Z.Y. PCL/PEG copolymeric nanoparticles: Potential nanoplatforms for anticancer agent delivery. Curr. Drug Targets. 2011, 12, 1131–1150. [Google Scholar] [CrossRef]
- Chountoulesi, M.; Selianitis, D.; Pispas, S.; Pippa, N. Recent Advances on PEO-PCL Block and Graft Copolymers as Nanocarriers for Drug Delivery Applications. Materials 2023, 16, 2298. [Google Scholar] [CrossRef]
- Boykin, C.; Zhang, G.; Chen, Y.H.; Zhang, R.W.; Fan, X.E.; Yang, W.M.; Lu, Q. Cucurbitacin IIa: A novel class of anti-cancer drug inducing non-reversible actin aggregation and inhibiting survivin independent of JAK2/STAT3 phosphorylation. Br. J. Cancer 2011, 104, 781–789. [Google Scholar] [CrossRef]
- Cai, Y.; Fang, X.; He, C.; Li, P.; Xiao, F.; Wang, Y.; Chen, M. Cucurbitacins: A systematic review of the phytochemistry and anticancer activity. Am. J. Chin. Med. 2015, 43, 1331–1350. [Google Scholar] [CrossRef]
- Kim, S.R.; Seo, H.S.; Choi, H.S.; Cho, S.G.; Kim, Y.K.; Hong, E.H.; Shin, Y.C.; Ko, S.G. Trichosanthes kirilowii ethanol extract and cucurbitacin D inhibit cell growth and induce apoptosis through inhibition of STAT3 activity in breast cancer cells. Evid. Based Complement. Alternat. Med. 2013, 2013, 975350. [Google Scholar] [CrossRef]
- Sun, J.; Blaskovich, M.A.; Jove, R.; Livingstone, S.K.; Coppola, D.; Sebti, S.M. Cucurbitacin Q: A selective STAT3 activation inhibitor with potent antitumor activity. Oncogene 2005, 24, 3236–3245. [Google Scholar] [CrossRef]
- Üremiş, M.M.; Üremiş, N.; Türköz, Y. Cucurbitacin E shows synergistic effect with sorafenib by inducing apoptosis in hepatocellular carcinoma cells and regulates Jak/Stat3, ERK/MAPK, PI3K/Akt/mTOR signaling pathways. Steroids 2023, 198, 109261. [Google Scholar] [CrossRef]
- Duangmano, S.; Sae-Lim, P.; Suksamrarn, A.; Patmasiriwat, P.; Domann, F.E. Cucurbitacin B causes increased radiation sensitivity of human breast cancer cells via G2/M cell cycle arrest. J. Oncol. 2012, 2012, 601682. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, K.; Ku, J.M.; Choi, Y.J.; Mok, K.; Kim, D.; Cheon, C.; Ko, S.G. Cucurbitacin D induces G2/M phase arrest and apoptosis via the ROS/p38 pathway in Capan-1 pancreatic cancer cell line. Evid. Based Complement. Alternat. Med. 2020, 2020, 6571674. [Google Scholar] [CrossRef]
- Xie, Y.L.; Tao, W.H.; Yang, T.X.; Qiao, J.G. Anticancer effect of cucurbitacin B on MKN-45 cells via inhibition of the JAK2/STAT3 signaling pathway. Exp. Ther. Med. 2016, 12, 2709–2715. [Google Scholar] [CrossRef]
- Ren, G.; Sha, T.; Guo, J.; Li, W.; Lu, J.; Chen, X. Cucurbitacin B induces DNA damage and autophagy mediated by reactive oxygen species (ROS) in MCF-7 breast cancer cells. J. Nat. Med. 2015, 69, 522–530. [Google Scholar] [CrossRef]
- Silva, V.R.; Santos, L.D.S.; Dias, R.B.; Quadros, C.A.; Bezerra, D.P. Emerging agents that target signaling pathways to eradicate colorectal cancer stem cells. Cancer Commun. 2021, 41, 1275–1313. [Google Scholar] [CrossRef]
- Silvestre, G.F.G.; de Lucena, R.P.; da Silva Alves, H. Cucurbitacins and the immune system: Update in research on anti-inflammatory, antioxidant, and immunomodulatory mechanisms. Curr. Med. Chem. 2022, 29, 3774–3789. [Google Scholar] [CrossRef]
- Rymal, K.S.; Chambliss, O.L.; Bond, M.D.; Smith, D.A. Squash Containing Toxic Cucurbitacin Compounds Occurring in California and Alabama. J. Food. Prot. 1984, 47, 270–271. [Google Scholar] [CrossRef]
- David, A.; Vallance, D.K. Bitter Principles of Cucurbitaceae. J. Pharm. Pharmacol. 1955, 7, 295–296. [Google Scholar] [CrossRef]
- Jorn, G.; Inge, S.; Hans, C.A. Cucurbitacins in Plant Food; Nordic Council of Ministers: Copenhagen, Denmark, 2006; p. 68. [Google Scholar] [CrossRef]
- Tamura, Y.; Maki, T.; Kan, K.; Nagayama, T.; Naoi, Y. Outbreaks of food poisoning through chemicals and natural toxicants in Tokyo. I. 1980–1982. Ann. Rep. Tokyo Metro. Res. Lab. Public Health 1983, 34, 171–177. [Google Scholar]
- Edery, H.; Schatzberg-Porath, G.; Gitter, S. Pharmacodynamic activity of elatericin (Cucurbitacin D). Arch. Int. Pharmacodyn. Ther. 1961, 130, 315–335. [Google Scholar] [PubMed]
- Njoroge, G.N.; Leonard, E.N. Edible and poisionous species of cucurbitaceae in the central highlands of Kenya. J. East. Afr. Nat. Hist. 1994, 83, 101–115. [Google Scholar] [CrossRef]
- Yaowalak, U.; Usaneeporn, L.; Weena, J.; Tanawan, K. Immunosuppressive effects of Cucurbitacin B on human peripheral blood lymphocytes. J. Med. Plants Res. 2010, 4, 2340–2347. [Google Scholar]
- Dai, S.; Wang, C.; Zhao, X.; Ma, C.; Fu, K.; Liu, Y.; Peng, C.; Li, Y. Cucurbitacin B: A Review of Its Pharmacology, Toxicity, and Pharmacokinetics. Pharmacol. Res. 2023, 187, 106587. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, W.; Gao, M.; Wu, C.; Yang, C.; Yang, J.; Wu, G.; Yang, B.; Kuang, H. Simultaneous Determination of Cucurbitacin B and Cucurbitacin E in Rat Plasma by UHPLC-MS/MS: A Pharmacokinetics Study after Oral Administration of Cucurbitacin Tablets. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1065–1066, 63–69. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, Q.; Wu, Q.; Chang, J.; Xue, H.; Liu, C.; Liu, X. A New Sensitive UPLC-MS/MS Method for the Determination of Cucurbitacin B in Rat Plasma: Application to an Absolute Bioavailability Study. RSC Adv. 2018, 8, 30978–30985. [Google Scholar] [CrossRef]
- Hunsakunachai, N.; Nuengchamnong, N.; Jiratchariyakul, W.; Kummalue, T.; Khemawoot, P. Pharmacokinetics of Cucurbitacin B from Trichosanthes cucumerina L. in Rats. BMC Complement. Altern. Med. 2019, 19, 157. [Google Scholar] [CrossRef]
- Tang, L.; Fu, L.; Zhu, Z.; Yang, Y.; Sun, B.; Shan, W.; Zhang, Z. Modified Mixed Nanomicelles with Collagen Peptides Enhanced Oral Absorption of Cucurbitacin B: Preparation and Evaluation. Drug Deliv. 2018, 25, 862–871. [Google Scholar] [CrossRef]
- Garg, S.; Kaul, S.C.; Wadhwa, R. Cucurbitacin B and cancer intervention: Chemistry, biology and mechanisms (Review). Int. J. Oncol. 2018, 52, 19–37. [Google Scholar] [CrossRef]
- Marostica, L.L.; de Barros, A.L.B.; Oliveira, J.; Salgado, B.S.; Cassali, G.D.; Leite, E.A.; Cardoso, V.N.; Lang, K.L.; Caro, M.S.B.; Duran, F.J.; et al. Antitumor effectiveness of a combined therapy with a new cucurbitacin B derivative and paclitaxel on a human lung cancer xenograft model. Toxicol. Appl. Pharmacol. 2017, 329, 272–281. [Google Scholar] [CrossRef]
- Liu, T.; Peng, H.; Zhang, M.; Deng, Y.; Wu, Z. Cucurbitacin B, a small molecule inhibitor of the Stat3 signaling pathway, enhances the chemosensitivity of laryngeal squamous cell carcinoma cells to cisplatin. Eur. J. Pharmacol. 2010, 641, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kang, J.G.; Kim, C.S.; Ihm, S.H.; Choi, M.G.; Yoo, H.J.; Lee, S.J. Doxorubicin has a synergistic cytotoxicity with cucurbitacin B in anaplastic thyroid carcinoma cells. Tumour Biol. 2017, 39, 1010428317692252. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Present Cucurbitacins |
|---|---|
| Acanthosicyos horridus | Cucurbitacins B and D, 23,24-dihydrocucurbitacin D (cucurbitacin R), 3-epi-isocucurbitacin D [20] |
| Alsomitra macrocarpa | Cucurbitacin E [21] |
| Bolbostemma paniculatum | Isocucurbitacin B, 23,24-dihydroisocucurbitacin B, cucurbitacin E, 23,24-dihydrocucurbitacin E [22] |
| Bryonia cretica | Cucurbitacins G, H, and J, and isocucurbitacins G and H [23]; Cucurbitacins B, D, E, and J, 23,24-dihydrocucurbitacins B and E, bryoniaosides A and B, and hexanocucurbitacin D [24] |
| Cayaponia racemosa | Cucurbitacin P, 11-deoxocucurbitacin P, deacetylpicracin [25] cucurbitacins B, D, and F, 23,24-dihydrocucurbitacins F and D (cucurbitacin R) [26] |
| Cayaponia tayuya | Cucurbitacin R and 23,24-dihydrocucurbitacin B [27] |
| Citrullus colocynthis | Cucurbitacins I, J, and T [28]; cucurbitacin B 2-O-β-D-glucopyranoside, cucurbitacin E 2-O-β-D-glucopyranoside [29]; 2-O-β-D-glucopyranosides of cucurbitacin E, I, J, K and L, and other derivatives [30]; ring-A-modified seco-cucurbitane triterpenoids [31] |
| Cucumis melo | Cucurbitacins B, D, and E [9]; cucurbitacins A, B, G, H, and R, 23,24-dihydrocucurbitacin B, 23,24-dihydroisocucurbitacin B, isocucurbitacin R, hexanorcucurbitacin D, arvenin I (cucurbitacin B 2-O-β-D-glucopyranoside), arvenin III (cucurbitacin D 2-O-β-D-glucopyranoside), 19-norlanosta-5,24-dien-11-one [32] |
| Cucumis prophetarum | Cucurbitacins B, E, I, and Q, isocucurbitacins B and D, 23,24-dihydrocucurbitacins B, E, I, and Q, 23,24-dihydroisocucurbitacin D [33] |
| Cucumis sativus | Cucurbitacin C and its derivatives [34] |
| Cucurbita andreana/ Cucurbita maxima | Cucurbitacins B, D, E, and I, 2-O-β-D-glucopyranosides of cucurbitacin B, E, and I [35] |
| Cucurbita pepo var. cylindrica | Cucurbitacin E 2-O-β-D-glucopyranoside [36] |
| Ecballium elaterium | Cucurbitacins B, D, E, and I [37]; Cucurbitacin D and 22-deoxocucurbitacin D [38] |
| Hemsleya ellipsoidea | Isocucurbitacin B, 23,24-dihydroisocucurbitacin B, cucurbitacins F, 25-O-acetylcucurbitacin F, 25-O-acetyl-23,24-dihydrocucurbitacin F [39] |
| Ibervillea sonorae | 23,24-Dihydrocucurbitacin F [40] |
| Lagenaria siceraria | Cucurbitacin I [41] |
| Luffa graveolense | Cucurbitacins D and E, isocucurbitacin B, cucurbitacin D 2-O-β-D-glucopyranoside, cucurbitacin B 20-O-β-D-glucopyranoside [42] |
| Luffa operculata | Cucurbitacins B, D, and E, isocucurbitacin B, neocucurbitacins A and B [18] |
| Momordica charantia | Momordicosides A, B, K, L, M, N, and S, karavilosides II and III, kuguaglycoside B and other derivatives [43] |
| Sechium edule var. nigrum spinosum | Cucurbitacins B, D, and I [44] |
| Trichosanthes cucumerina | Cucurbitacin B, 23,24-dihydrocucurbitacin B, bryonolic acid, bryononic acid [45] |
| Trichosanthes kirilowii | Cucurbitacins B and D, isocucurbitacins B and D, 3-epi-isocucurbitacin B, 23,24-dihydrocucurbitacins B and E, 23,24-dihydroisocucurbitacin B [46]; Cucurbitacin D, 23,24-dihydrocucurbitacin D [47] |
| Trichosanthes tricuspidata | 2-O-β-D-glucopyranosides of cucurbitacine J, K, L and 25-O-acetylcucurbitacin L 2-O-β-D-glucopyranoside, khekadaengosides A–N [48] |
| Wilbrandia ebracteata | Cucurbitacins B, D, E, G, H, J, K, P and R, 23,24-dihydrocucurbitacins B and E, 23,24-dihydroisocucurbitacin B, 22-deoxocucurbitacin D, 3-epi-isocucurbitacin G, (20R)-25-acetoxy-3,16α,20-trihydroxy-30-nor-2-(β-D-glucopyranosyloxy)-1,2,3,4,5,10-dehydrocucurbit-6-ene-11,22-dione [17] |
| Family Name | Plant Name | Present Cucurbitacins |
|---|---|---|
| Begoniaceae | Begonia heracleifolia | Cucurbitacins B and D, and their 2-O-β-D-glucopyranosides, 23,24-dihydrocucurbitacins F and D (cucurbitacin R) [49] |
| Begonia nantoensis | Cucurbitacins B, E, I, and 23,24-dihydrocucurbitacins B and E [50] | |
| Brassicaceae | Iberis amara | Cucurbitacins E and I, and their 2-O-β-D-glucopyranosides [51] |
| Iberis gibraltarica | Cucurbitacins D, E and I [52] | |
| Lepidium sativum | Cucurbitacin I [52] | |
| Cercidiphyllaceae | Cercidiphyllum japonicum | Cucurbitacin D [53] |
| Datiscaceae | Datisca glomerata | Cucurbitacins B, D, F and cucurbitacin glycosides (datisosides B, C, D, E, F, G, H) [54] |
| Columelliaceae | Desfontania spinosa | 23,24-dihydro-11-deoxocucurbitacin I, and spinosides A and B [55] |
| Chrysobalanaceae | Licania intrapetiolaris | Cucurbitacin B [56] |
| Elaeocarpaceae | Elaeocarpus hainanensis | Cucurbitacins D, F, G, I, O, 3-epi-isocucurbitacin G, and other derivatives [57] |
| Elaeocarpus mastersii | Cucurbitacins D and F [58] | |
| Elaeocarpus sylvestris | Cucurbitacin D and mogroside derivatives [59] | |
| Sloanea zuliaensis | Cucurbitacin D, 2-deoxycucurbitacin D, and 25-acetylcucurbitacin F [60] | |
| Lauraceae | Machilus yaoshansis | Machilaminosides A and B (derivatives of cucurbitacin I) [61]; 2-O-β-D-glucopyranosides of cucurbitacin I, J, and K and their derivatives [62] |
| Malvaceae | Helicteres isora | Cucurbitacin B and isocucurbitacin B [63] |
| Plantaginaceae | Bacopa monnieri | Bacobitacins A, B, C, D and cucurbitacin E [64] |
| Conobea scoparioides | Cucurbitacin E [65] | |
| Neopicrorhiza scrophulariiflora | Scrophoside A, cucurbitacin glycosides [66] | |
| Picria fel-terrae | Picfeltarraenin IA, picfeltarraenin IB, picfeltarraenin IV, and picfeltarraenin VI [67] | |
| Picrorhiza kurroa | Cucurbitacin B, 23,24-dihydrocucurbitacin B, and various derivatives and glycosides of cucurbitacin B [68,69] | |
| Picrorhiza scrophulariiflora | 2-O-β-D-glucopyranosyl-3,16,20, 25-tetrahydroxy-9-methyl-19-norlanosta-5, 23-diene-22-one, 2-O-β-D-glucopyranosyl-3,16, 20-trihydroxy-25-acetoxy-9-methyl-19-norlanosta-5, 23-diene-22-one, 2-O-β-D-glucopyranosyl-4,4,9, 14-tetramethyl-19-norpregn-5-en-20-one [70] | |
| Polemoniaceae | Ipomopsis aggregata | Cucurbitacin B, isocucurbitacin B, and 3-epi-isocucurbitacin B [71] |
| Primulaceae | Anagallis arvensis | Arvenin I (cucurbitacin B 2-O-β-D-glucopyranoside), arvenin II (23,24-dihydrocucurbitacin B 2-O-β-D-glucopyranoside), arvenin III (cucurbitacin D 2-O-β-D-glucopyranoside), arvenin IV (cucurbitacin R 2-O-β-D-glucopyranoside) [72] |
| Rosaceae | Kageneckia angustifolia | Cucurbitacin F, 2,3,16-triacetylcucurbitacin F [73] |
| Kageneckia oblonga | 3β-(β-D-glucosyloxy)-16α,23α-epoxycucurbita-5,24-dien-11-one [74] | |
| Physocarpus capitatus | Cucurbitacin F, dihydrocucurbitacin F, and hydroxyl/acetyl derivatives of cucurbitacin F [75] | |
| Physocarpus opulifolius | Cucurbitacin D, cucurbitacin F, and 3-epi-isocucurbitacin D [76] | |
| Purshia mexicana (Cowania mexicana) | Cucurbitacin F, 23,24-dihydrocucurbitacin F, 15-oxo-cucurbitacin F, 15-oxo-23,24-dihydrocucurbitacin F [77] | |
| Sorbaria sorbifolia var. stellipila | Cucurbitacins D and F [78] | |
| Rubiaceae | Hintonia standleyana | 23,24-dihydrocucurbitacin F 3-O-β-D-glucopyranoside [79] |
| Nernstia mexicana (Cigarrilla mexicana) | Cucurbitacin E, isocucurbitacin B, epi-isocucurbitacin B [80]; arvenin I (cucurbitacin B 2-O-β-D-glucopyranoside) [81] | |
| Thymelaeaceae | Aquilaria sinensis | Cucurbitacin I 2-O-β-D-glucopyranoside, bryoamaride (Cucurbitacin L 2-O-β-D-glucopyranoside) [82] |
| Gonystylus keithii | Cucurbitacins B and D [83] | |
| Tricholomataceae (Fungi) | Leucopaxillus gentianeus | Cucurbitacin B, oleyl, linoleyl and palmityl esters of cucurbitacin B, leucopaxillones A and B [12]; cucurbitacin D, 16-deoxycucurbitacin B, 18-deoxyleucopaxillone A [13] |
| Cucurbitacin Type | Cancer Type | Mechanism | References |
|---|---|---|---|
| Cucurbitacin A | lung | cell cycle arrest | [90] |
| Cucurbitacin A | ovarian | induce apoptosis, cell cycle arrest | [91] |
| Cucurbitacin B | breast | anti-metastatic | [92] |
| Cucurbitacin B | breast | cell cycle arrested | [93] |
| Cucurbitacin B | breast | induce apoptosis, inhibit cell migration | [94] |
| Cucurbitacin B | breast | induce apoptosis, cell cycle arrest | [95] |
| Cucurbitacin B | breast | inhibit cell proliferation, induce apoptosis, cell cycle arrest | [96] |
| Cucurbitacin B | cholangiocarcinoma | anti-metastatic | [97] |
| Cucurbitacin B | colon | inhibit cell growth, cell cycle arrest | [98] |
| Cucurbitacin B | colon | inhibit cell proliferation, induce apoptosis | [97] |
| Cucurbitacin B | gastric | induce apoptosis | [99] |
| Cucurbitacin B | glioblastoma | induce apoptosis | [100] |
| Cucurbitacin B | hepatoma | induce apoptosis | [101] |
| Cucurbitacin B | glioblastoma | inhibit angiogenesis | [102] |
| Cucurbitacin B | laryngeal squamous | cell cycle arrest | [103] |
| Cucurbitacin B | lung | induce apoptosis | [104] |
| Cucurbitacin B | lung | anti-metastatic | [105] |
| Cucurbitacin B | lung | induce apoptosis | [106] |
| Cucurbitacin B | lymphoma | induce apoptosis | [107] |
| Cucurbitacin B | melanoma | cell cycle arrest | [108] |
| Cucurbitacin B | neuroblastoma | induce apoptosis | [109] |
| Cucurbitacin B | osteosarcoma | induce apoptosis | [110] |
| Cucurbitacin B | osteosarcoma | induce apoptosis, cell cycle arrest | [111] |
| Cucurbitacin B | ovarian | increase the sensitivity of cisplatin | [112] |
| Cucurbitacin B | pancreatic | induce apoptosis | [113] |
| Cucurbitacin B | pancreatic | inhibit cell proliferation | [114] |
| Cucurbitacin B | pancreatic | inhibit cell proliferation | [115] |
| Cucurbitacin B | tongue squamous | inhibit cell proliferation | [116] |
| Cucurbitacin C | colon | inhibition cell growth, cell cycle arrest, induce apoptosis | [8] |
| Cucurbitacin C | lung | inhibit cell proliferation, induce apoptosis, cell cycle arrest | [8] |
| Cucurbitacin C | prostate | inhibit cell proliferation | [8] |
| Cucurbitacin D | breast | inhibit cell proliferation, induce apoptosis | [117] |
| Cucurbitacin D | cervical | inhibit cells growth and metastasis, cell cycle arrest | [118] |
| Cucurbitacin D | pancreatic | cell cycle arrest | [119] |
| Cucurbitacin D | pancreatic | induce apoptosis, cell cycle arrest | [112] |
| Cucurbitacin D | prostate | inhibit cell growth | [120] |
| Cucurbitacin D | lung | cell cycle arrest | [121] |
| Cucurbitacin E | glioblastoma | inhibit cell proliferation, induce apoptosis, cell cycle arrest | [122] |
| Cucurbitacin E | lung | induce apoptosis, cell cycle arrest | [123] |
| Cucurbitacin E | malignant glioma | antiproliferative, inhibit cell growth, cell cycle arrest | [124] |
| Cucurbitacin E | breast | inhibit cell growth | [125] |
| Cucurbitacin E | gastric | enhance the cytotoxicity of DOX in cells | [126] |
| Cucurbitacin I | colon | inhibit cell proliferation | [127] |
| Cucurbitacin I | colon | inhibit cell migration | [128] |
| Cucurbitacin I | colon | induce apoptosis, inhibit cell proliferation, cell cycle arrest | [129] |
| Cucurbitacin I | lung | induce apoptosis | [130] |
| Cucurbitacin I | lung | autophagy induction | [131] |
| Cucurbitacin I | ovarian | induce apoptosis, autophagy induction | [132] |
| Cucurbitacin I | ovarian | inhibit cell growth | [133] |
| Cucurbitacin I | pancreatic | inhibit cell growth | [134] |
| Cucurbitacin I | pancreatic | induce apoptosis, induce autophagy | [132] |
| Cucurbitacin IIa (25-O-acetyl-23,24-dihydrocucurbitacin F) | lung | induce apoptosis, cell cycle arrest | [135] |
| Cucurbitacin IIb (23,24-dihydrocucurbitacin F) | lung | induce apoptosis, cell cycle arrest | [136] |
| lung | inhibit cell growth, induce apoptosis | [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieniuk, B.; Pawełkowicz, M. Recent Advances in the Application of Cucurbitacins as Anticancer Agents. Metabolites 2023, 13, 1081. https://doi.org/10.3390/metabo13101081
Zieniuk B, Pawełkowicz M. Recent Advances in the Application of Cucurbitacins as Anticancer Agents. Metabolites. 2023; 13(10):1081. https://doi.org/10.3390/metabo13101081
Chicago/Turabian StyleZieniuk, Bartłomiej, and Magdalena Pawełkowicz. 2023. "Recent Advances in the Application of Cucurbitacins as Anticancer Agents" Metabolites 13, no. 10: 1081. https://doi.org/10.3390/metabo13101081
APA StyleZieniuk, B., & Pawełkowicz, M. (2023). Recent Advances in the Application of Cucurbitacins as Anticancer Agents. Metabolites, 13(10), 1081. https://doi.org/10.3390/metabo13101081