


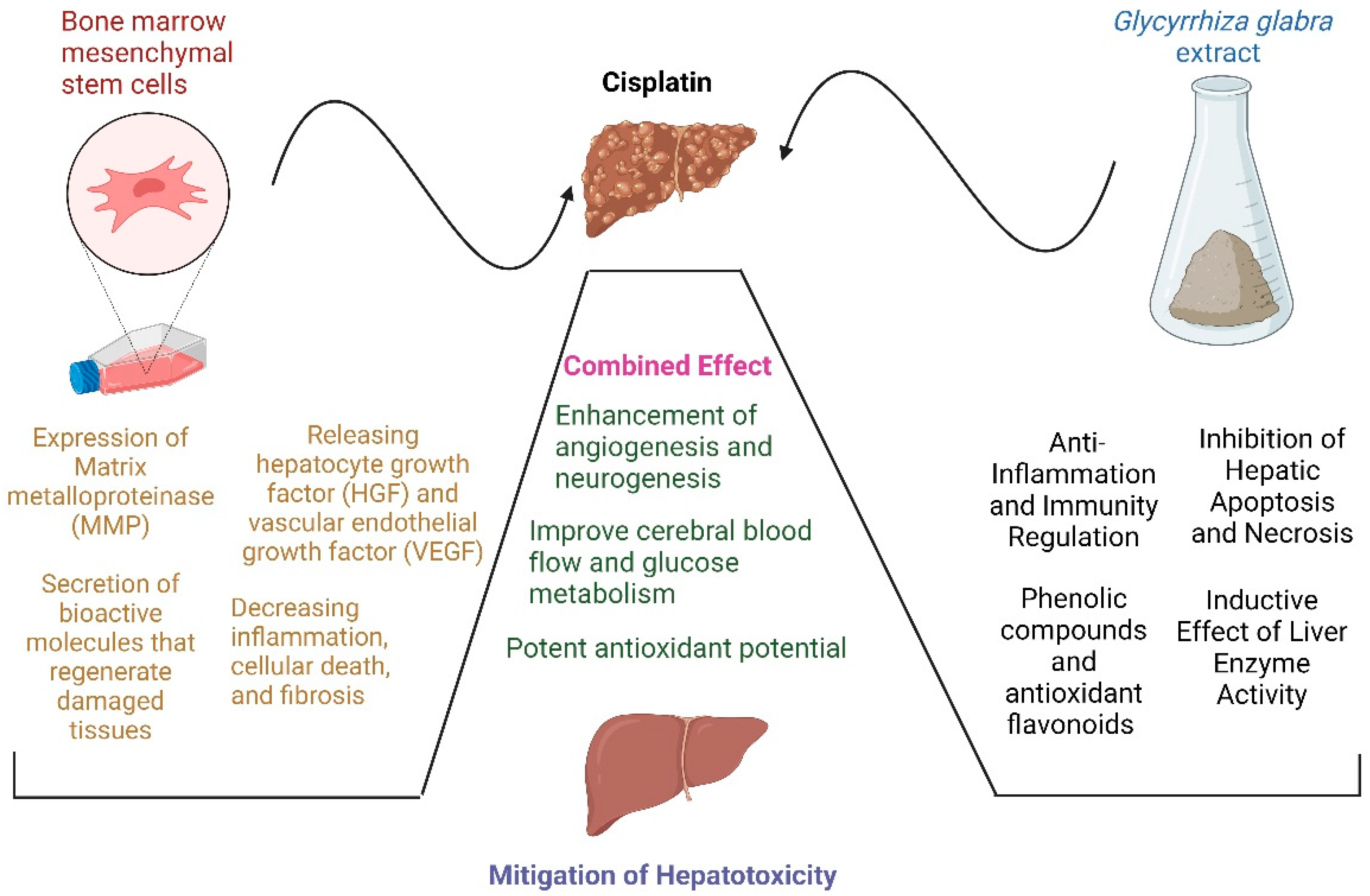
The Combined Effect of Licorice Extract and Bone Marrow Mesenchymal Stem Cells on Cisplatin-Induced Hepatocellular Damage in Rats

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation of Methanolic Extract from GGE Roots
2.2. Phytochemical Analysis and Antioxidant Activity of GGE
2.3. BM-MSCs Preparation
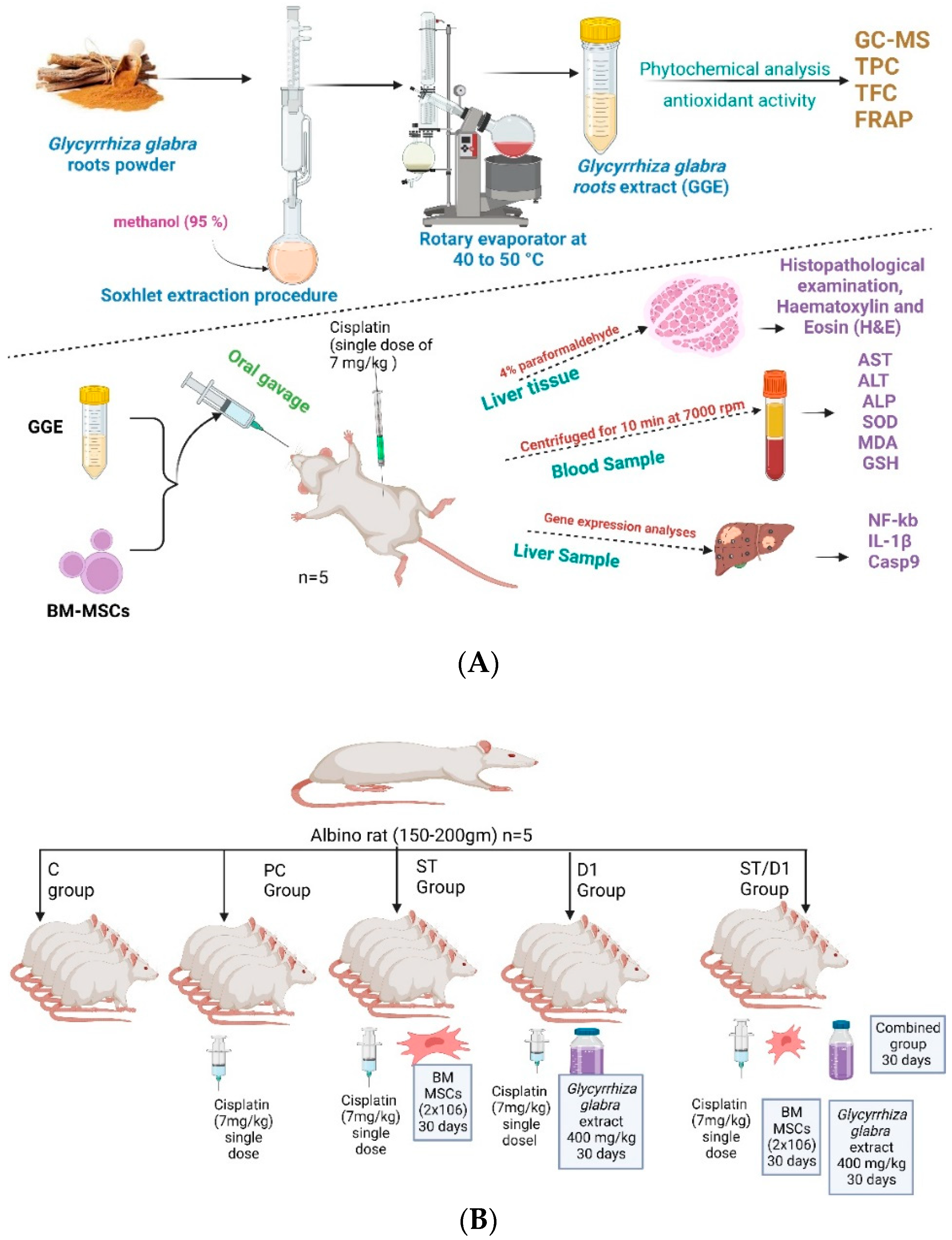
2.4. Animal Model’s Design
- The control group got saline.
- The cisplatin group (P.C. group): 7 mg/kg of cisplatin was injected intraperitoneally (I.P.) as a single dose to induce liver damage (Positive management).
- The BM-MSCs group (Mesenchymal Stem Cells from Bone Marrow—St group): 7 mg/kg of cisplatin was injected intraperitoneally (I.P.) as a single dose, and on the following day, rats began receiving 2 × 106 BM-MSCs per day in phosphate buffer solution (PBS) by intravenous (IV) injection for 1 month.
- The GGE group (licorice-based drug D1 group): 7 mg/kg of cisplatin was injected intraperitoneally (I.P.) as a single dose, and the following day, rats received 400 mg/kg/day orally of GGE for one month.
- The group consisting of BM-MSCs and GGE (Combinational treatment—St/D1 group) 7 mg/kg of cisplatin was injected intraperitoneally (I.P.) as a single dose, and the following day, rats received 2 × 106 BM-MSCs in PBS by IV injection plus 400 mg/kg/day of GGE one month [31].
2.5. Blood and Liver Tissue Collection
2.5.1. Biochemical Markers of Serum
2.5.2. Histopathological Examination, Hematoxylin, and Eosin (H&E)
2.6. Gene Expression Analyses
2.7. Statistical Analysis
3. Results
3.1. Phytochemical Analysis and the Antioxidant Power of GGE
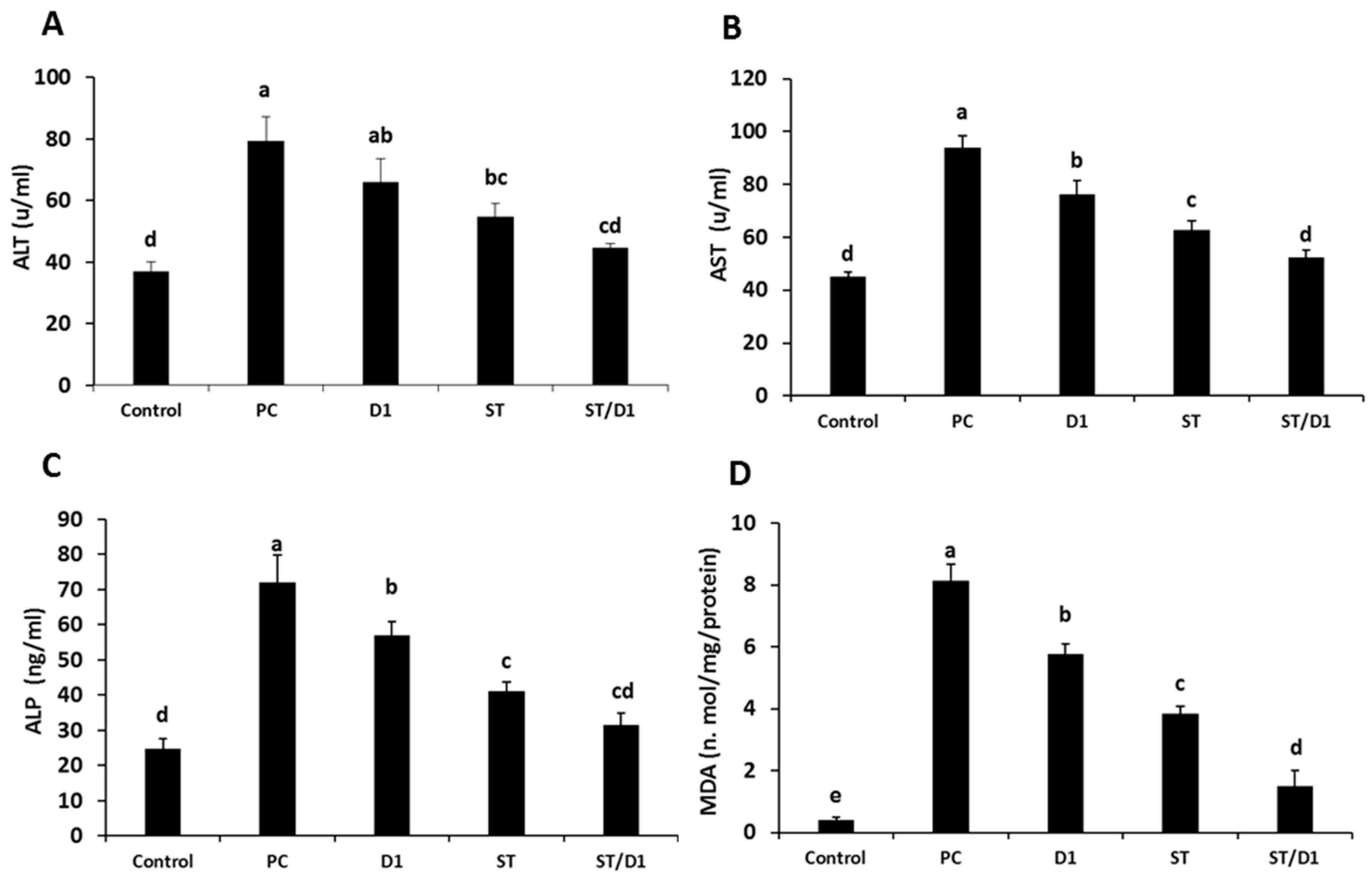
3.2. Biochemical Parameters
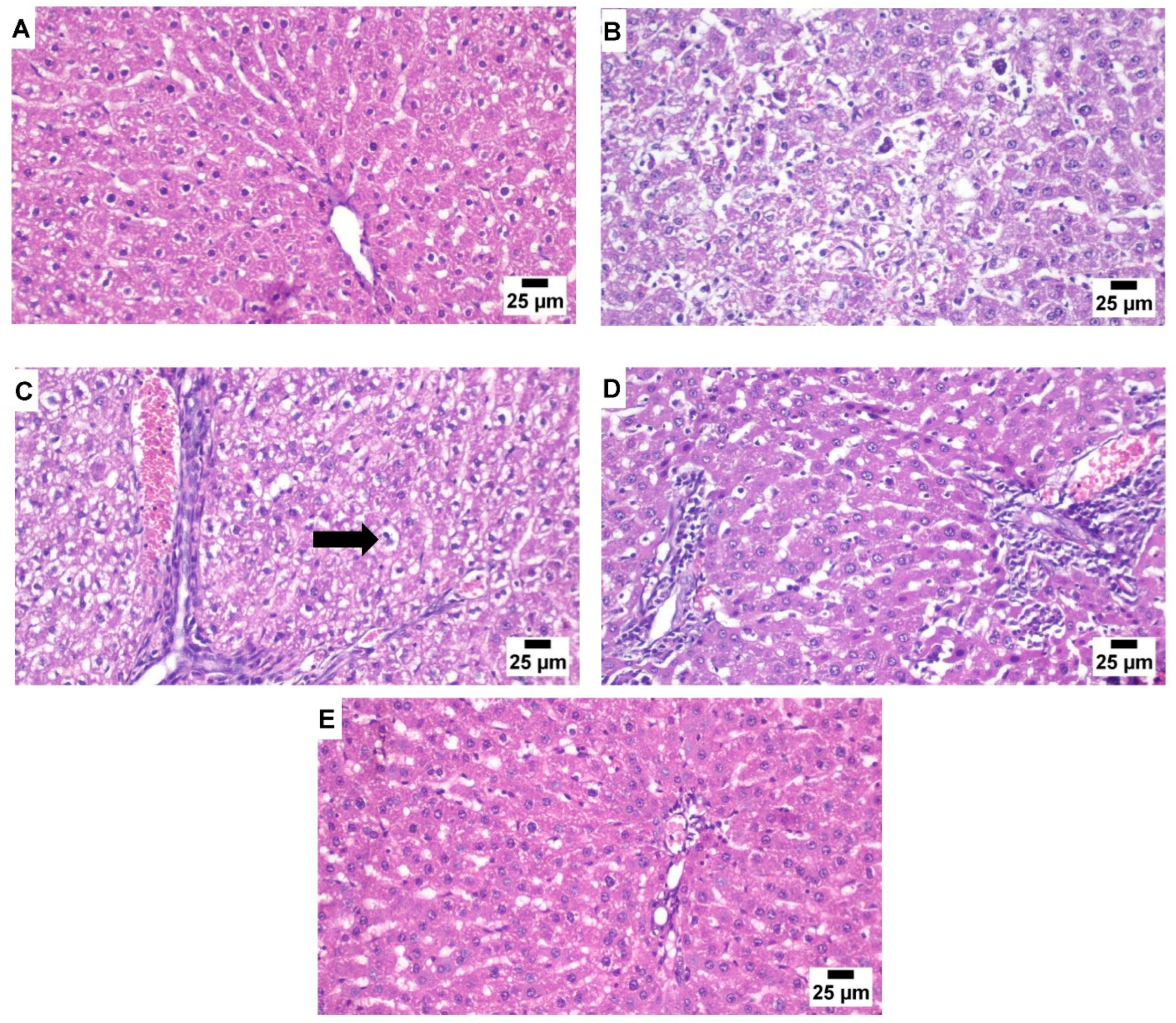
3.3. Liver Histopathology
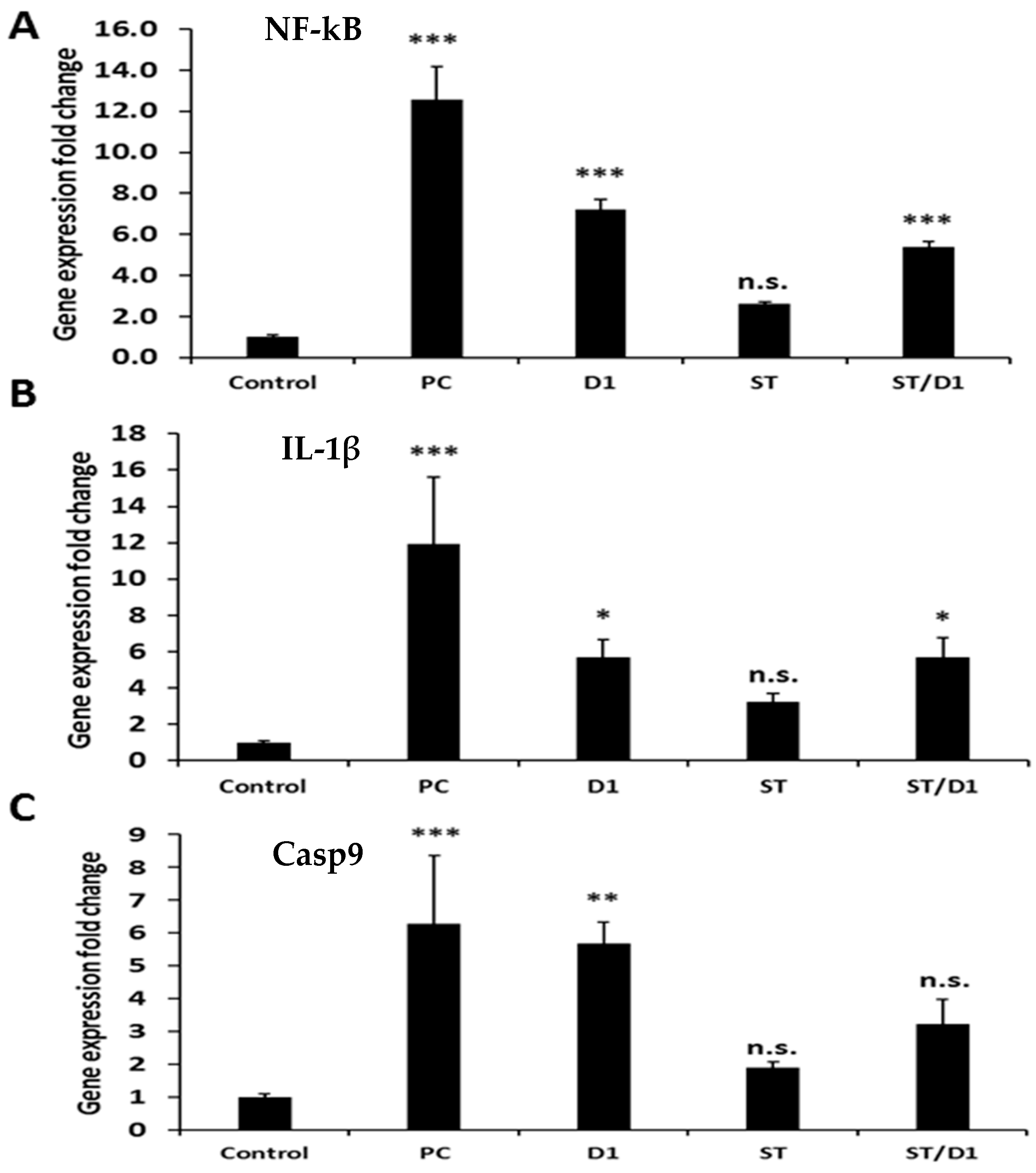
3.4. Gene Expression Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aminuddin, A.; Ng, P.Y.; Leong, C.-O.; Chua, E.W. Mitochondrial DNA Alterations May Influence the Cisplatin Responsiveness of Oral Squamous Cell Carcinoma. Sci. Rep. 2020, 10, 7885. [Google Scholar] [CrossRef] [PubMed]
- Arany, I.; Safirstein, R.L. Cisplatin Nephrotoxicity. Semin. Nephrol. 2003, 23, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Pabla, N.; Dong, Z. Cisplatin Nephrotoxicity: Mechanisms and Renoprotective Strategies. Kidney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Pinto, P.; Justian, N.; Dib, M.; Fischer, J.; Somova, M.; Burchardt, M.; Wolff, I. In Vitro Characterization of Renal Drug Transporter Activity in Kidney Cancer. Int. J. Mol. Sci. 2022, 23, 10177. [Google Scholar] [CrossRef] [PubMed]
- Hosohata, K. Role of Oxidative Stress in Drug-Induced Kidney Injury. Int. J. Mol. Sci. 2016, 17, 1826. [Google Scholar] [CrossRef]
- Quintanilha, J.C.F.; de Sousa, V.M.; Visacri, M.B.; Amaral, L.S.; Santos, R.M.M.; Zambrano, T.; Salazar, L.A.; Moriel, P. Involvement of Cytochrome P450 in Cisplatin Treatment: Implications for Toxicity. Cancer Chemother. Pharmacol. 2017, 80, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Abd Rashid, N.; Abd Halim, S.A.S.; Teoh, S.L.; Budin, S.B.; Hussan, F.; Ridzuan, N.R.A.; Jalil, N.A.A. The Role of Natural Antioxidants in Cisplatin-Induced Hepatotoxicity. Biomed. Pharmacother. 2021, 144, 112328. [Google Scholar] [CrossRef]
- Huo, H.Z.; Wang, B.; Liang, Y.K.; Bao, Y.Y.; Gu, Y. Hepatoprotective and Antioxidant Effects of Licorice Extract against CCl4-Induced Oxidative Damage in Rats. Int. J. Mol. Sci. 2011, 12, 6529–6543. [Google Scholar] [CrossRef]
- Contreras-Zentella, M.L.; Hernández-Muñoz, R. Is Liver Enzyme Release Really Associated with Cell Necrosis Induced by Oxidant Stress? Oxidative Med. Cell. Longev. 2016, 2016, 3529149. [Google Scholar] [CrossRef]
- Sadhukhan, P.; Saha, S.; Dutta, S.; Sil, P.C. Mangiferin Ameliorates Cisplatin Induced Acute Kidney Injury by Upregulating Nrf-2 via the Activation of PI3K and Exhibits Synergistic Anticancer Activity with Cisplatin. Front. Pharmacol. 2018, 9, 638. [Google Scholar] [CrossRef]
- Cüre, M.C.; Erkan, C.; Kalkan, Y.; Kirbaş, A.; Tümkaya, L.; Yilmaz, A.; Türkyilmaz, A.K.; Şehitoğlu, İ.; Süleyman, Y. Infliximab Modulates Cisplatin-Induced Hepatotoxicity in Rats. Balk. Med. J. 2016, 33, 504–511. [Google Scholar] [CrossRef]
- El-Naggar, S.A.; El-Said, K.S.; Mobasher, M.; Elbakry, M. Enhancing antitumor efficacy of cisplatin low dose by EDTA in Ehrlich ascetic carcinoma bearing mice. Braz. Arch. Biol. Technol. 2019, 62, e19180716. [Google Scholar] [CrossRef]
- Florea, A.-M.; Büsselberg, D. Cisplatin as an Anti-Tumor Drug: Cellular Mechanisms of Activity, Drug Resistance and Induced Side Effects. Cancers 2011, 3, 1351–1371. [Google Scholar] [CrossRef]
- Liao, Y.; Lu, X.; Lu, C.; Li, G.; Jin, Y.; Tang, H. Selection of Agents for Prevention of Cisplatin-Induced Hepatotoxicity. Pharmacol. Res. 2008, 57, 125–131. [Google Scholar] [CrossRef]
- Wahab, S.; Annadurai, S.; Abullais, S.S.; Das, G.; Ahmad, W.; Ahmad, M.F.; Kandasamy, G.; Vasudevan, R.; Ali, M.S.; Amir, M. Glycyrrhiza Glabra (Licorice): A Comprehensive Review on Its Phytochemistry, Biological Activities, Clinical Evidence and Toxicology. Plants 2021, 10, 2751. [Google Scholar] [CrossRef]
- Zhang, Q.; Ye, M. Chemical Analysis of the Chinese Herbal Medicine Gan-Cao (Licorice). J. Chromatogr. A 2009, 1216, 1954–1969. [Google Scholar] [CrossRef]
- Tang, B.; Qiao, H.; Meng, F.; Sun, X. Glycyrrhizin Attenuates Endotoxin- Induced Acute Liver Injury after Partial Hepatectomy in Rats. Braz. J. Med. Biol. Res. 2007, 40, 1637–1646. [Google Scholar] [CrossRef] [PubMed]
- Sohail, M.; Rakha, A.; Butt, M.S.; Asghar, M. Investigating the Antioxidant Potential of Licorice Extracts Obtained through Different Extraction Modes. J. Food Biochem. 2018, 42, e12466. [Google Scholar] [CrossRef]
- Yin, G.; Cao, L.; Xu, P.; Jeney, G.; Nakao, M.; Lu, C. Hepatoprotective and Antioxidant Effects of Glycyrrhiza Glabra Extract against Carbon Tetrachloride (CCl4)-Induced Hepatocyte Damage in Common Carp (Cyprinus Carpio). Fish Physiol. Biochem. 2011, 37, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Zahran, F.; Nabil, A.; El Karef, A.; Lotfy, A.; Mahmoud, K.; Hozayen, W.G.; Sobh, M. Effect of Antioxidants and Mesenchymal Stem Cells on Cisplatin Induced Renal Fibrosis in Rats. J. Stem Cell Res. Ther. 2016, 1, 1. [Google Scholar]
- Rosenbaum, A.J.; Grande, D.A.; Dines, J.S. The Use of Mesenchymal Stem Cells in Tissue Engineering: A Global Assessment. Organogenesis 2008, 4, 23–27. [Google Scholar] [CrossRef]
- Solter, D.; Gearhart, J. Putting Stem Cells to Work. Science 1999, 283, 1468–1470. [Google Scholar] [CrossRef] [PubMed]
- Zahran, F.; El-Ghareb, M.; Nabil, A. Bone Marrow Derived Mesenchymal Stem Cells As A Therapy for Renal Injury. IJAR 2014, 4, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Khalil, M.R.; El-Demerdash, R.S.; Elminshawy, H.H.; Mehanna, E.T.; Mesbah, N.M.; Abo-Elmatty, D.M. Therapeutic Effect of Bone Marrow Mesenchymal Stem Cells in a Rat Model of Carbon Tetrachloride Induced Liver Fibrosis. Biomed. J. 2021, 44, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Akram, R.; Saqib, M.; Khalid, B.; Arshad, A. Effects of Extraction Media and Techniques on the Antioxidant Properties and Recovery of Phenolics from Roots of Glycyrrhiza Glabra. J. Mol. Pathophysiol. 2015, 4, 138–143. [Google Scholar] [CrossRef]
- Koldaş, S.; Demirtas, I.; Ozen, T.; Demirci, M.A.; Behçet, L. Phytochemical Screening, Anticancer and Antioxidant Activities of Origanum Vulgare L. Ssp. Viride (Boiss.) Hayek, a Plant of Traditional Usage. J. Sci. Food Agric. 2015, 95, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin–Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Sahu, R.; Saxena, J. Screening of Total Phenolic and Flavonoid Content in Conventional and Non-Conventional Species of Curcuma. J. Pharmacogn. Phytochem. 2013, 21, 24–26. [Google Scholar]
- Jang, H.-J.; Lee, H.-J.; Yoon, D.-K.; Ji, D.-S.; Kim, J.-H.; Lee, C.-H. Antioxidant and Antimicrobial Activities of Fresh Garlic and Aged Garlic By-Products Extracted with Different Solvents. Food Sci. Biotechnol. 2018, 27, 219–225. [Google Scholar] [CrossRef]
- Amin, M.A.; Sabry, D.; Rashed, L.A.; Aref, W.M.; el-Ghobary, M.A.; Farhan, M.S.; Fouad, H.A.; Youssef, Y.A.-A. Short-Term Evaluation of Autologous Transplantation of Bone Marrow–Derived Mesenchymal Stem Cells in Patients with Cirrhosis: Egyptian Study. Clin. Transplant. 2013, 27, 607–612. [Google Scholar] [CrossRef]
- Hassen, M.T.; Mohamed, H.K.; Montaser, M.M.; El-Sharnouby, M.E.; Awad, N.; Ebiya, R.A. Molecular, Immunomodulatory, and Histopathological Role of Mesenchymal Stem Cells and Beetroot Extract on Cisplatin Induced Testicular Damage in Albino Rats. Animals 2021, 11, 1142. [Google Scholar] [CrossRef] [PubMed]
- Mathai, C.K.; Beutler, E. Blood Preservatives for the Glutathione Stability Test. Nature 1965, 207, 204–205. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: I. Occurrence in Higher Plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, T.; Kawada, K.; Shimada, T.; Mori, M. Lipid Peroxidation in Maternal and Cord Blood and Protective Mechanism against Activated-Oxygen Toxicity in the Blood. Am. J. Obstet. Gynecol. 1979, 135, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Haddadi, P.; Mahdavi, M.; Rahbarghazi, R. Down-Regulation of Bcl2 and Survivin, and up-Regulation of Bax Involved in Copper (II) Phenylthiosemicarbazone Complex-Induced Apoptosis in Leukemia Stem-like KG1a Cells. Process Biochem. 2020, 92, 190–196. [Google Scholar] [CrossRef]
- Taghizadeh, F.; Hosseinimehr, S.J.; Zargari, M.; Karimpour Malekshah, A.; Mirzaei, M.; Talebpour Amiri, F. Alleviation of Cisplatin-induced Hepatotoxicity by Gliclazide: Involvement of Oxidative Stress and Caspase-3 Activity. Pharm. Res. Perspect 2021, 9, e00788. [Google Scholar] [CrossRef]
- Francis, P.; Navarro, V.J. Drug Induced Hepatotoxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Mobasher, M.A.; Valverde, Á.M. Signaling pathways involved in paracetamol-induced hepatotoxicity: New insights on the role of protein tyrosine phosphatase 1B. Arch. Physiol. Biochem. 2014, 120, 51–63. [Google Scholar] [CrossRef]
- Kuna, L.; Božić, I.; Kizivat, T.; Bojanić, K.; Mršo, M.; Kralj, E.; Smolić, R.; Wu, G.Y.; Smolić, M. Models of Drug Induced Liver Injury (DILI)—Current Issues and Future Perspectives. Curr. Drug Metab. 2018, 19, 830–838. [Google Scholar] [CrossRef]
- Bilgic, Y.; Akbulut, S.; Aksungur, Z.; Erdemli, M.E.; Ozhan, O.; Parlakpinar, H.; Vardi, N.; Turkoz, Y. Protective Effect of Dexpanthenol against Cisplatin-Induced Hepatotoxicity. Exp. Ther. Med. 2018, 16, 4049–4057. [Google Scholar] [CrossRef]
- Ellabban, M.A.; Fattah, I.O.A.; Kader, G.A.; Eldeen, O.S.; Mehana, A.E.; Khodeer, D.M.; Hosny, H.; Elbasiouny, M.S.; Masadeh, S. The Effects of Sildenafil and/or Nitroglycerin on Random-Pattern Skin Flaps After Nicotine Application in Rats. Sci. Rep. 2020, 10, 3212. [Google Scholar] [CrossRef]
- Almaeen, A.H.; Alduraywish, A.A.; Mobasher, M.A.; Almadhi, O.I.M.; Nafeh, H.M.; El-Metwally, T.H. Oxidative stress, immunological and cellular hypoxia biomarkers in hepatitis C treatment-naïve and cirrhotic patients. Arch Med Sci. 2020, 17, 368–375. [Google Scholar] [CrossRef]
- El-Said, K.S.; Atta, A.; Mobasher, M.A.; Germoush, M.O.; Mohamed, T.M.; Salem, M.M. Quercetin mitigates rheumatoid arthritis by inhibiting adenosine deaminase in rats. Mol. Med. 2022, 28, 24. [Google Scholar] [CrossRef] [PubMed]
- Real, M.; Barnhill, M.S.; Higley, C.; Rosenberg, J.; Lewis, J.H. Drug-Induced Liver Injury: Highlights of the Recent Literature. Drug Saf. 2019, 42, 365–387. [Google Scholar] [CrossRef] [PubMed]
- Avrutsky, M.I.; Troy, C.M. Caspase-9: A Multimodal Therapeutic Target with Diverse Cellular Expression in Human Disease. Front. Pharmacol. 2021, 12, 1728. [Google Scholar] [CrossRef]
- Kuida, K. Caspase-9. Int. J. Biochem. Cell Biol. 2000, 32, 121–124. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Beshbishy, A.M.; El-Mleeh, A.; Abdel-Daim, M.M.; Devkota, H.P. Traditional Uses, Bioactive Chemical Constituents, and Pharmacological and Toxicological Activities of Glycyrrhiza Glabra L.(Fabaceae). Biomolecules 2020, 10, 352. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-C.; Lee, Y.-H.; Kim, S.H.; Kim, K.-J.; Kim, K.-M.; Oh, S.; Jung, Y.-S. Hepatoprotective Effect of Licorice, the Root of Glycyrrhiza Uralensis Fischer, in Alcohol-Induced Fatty Liver Disease. BMC Complement Altern. Med. 2016, 16, 19. [Google Scholar] [CrossRef]
- Aithal, A.P.; Bairy, L.K.; Seetharam, R.N.; Kumar, N. Hepatoprotective Effect of Bone Marrow-Derived Mesenchymal Stromal Cells in CCl4-Induced Liver Cirrhosis. 3 Biotech. 2021, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Kaur, H.; Dhindsa, A.S. Glycyrrhiza glabra: A phytopharmacological review. Int. J. Pharm. Sci. Res. 2013, 4, 2470. [Google Scholar]
- Vijayalakshmi, U.; Shourie, A. Evaluation of Different Methods for Extraction of Antioxidant Phenolic Compounds from Glycyrrhiza Glabra Roots. World J. Pharm. Res. 2015, 4, 1524–1537. [Google Scholar]
- Damle, M. Glycyrrhiza Glabra (Liquorice)—A Potent Medicinal Herb. Int. J. Herb. Med. 2014, 2, 132–136. [Google Scholar]
- El-Tahawy, N.F.; Ali, A.H.; Saied, S.R.; Abdel-Wahab, Z. Effect of Glycyrrhizin on Lipopolysaccharide/D-Galactosamine-Induced Acute Hepatitis in Albino Rats: A Histological and Immunohistochemical Study. Egypt. J. Histol. 2011, 34, 518–527. [Google Scholar] [CrossRef]
- Mohamed, H.K.; Mobasher, M.A.; Ebiya, R.A.; Hassen, M.T.; Hagag, H.M.; El-Sayed, R.; Abdel-Ghany, S.; Said, M.M.; Awad, N.S. Anti-Inflammatory, Anti-Apoptotic, and Antioxidant Roles of Honey, Royal Jelly, and Propolis in Suppressing Nephrotoxicity Induced by Doxorubicin in Male Albino Rats. Antioxidants 2022, 11, 1029. [Google Scholar] [CrossRef]
- Li, J.; Cao, H.; Liu, P.; Cheng, G.; Sun, M. Glycyrrhizic Acid in the Treatment of Liver Diseases: Literature Review. Biomed. Res. Int. 2014, 2014, 872139. [Google Scholar] [CrossRef]
- Honda, H.; Nagai, Y.; Matsunaga, T.; Saitoh, S.-I.; Akashi-Takamura, S.; Hayashi, H.; Fujii, I.; Miyake, K.; Muraguchi, A.; Takatsu, K. Glycyrrhizin and Isoliquiritigenin Suppress the LPS Sensor Toll-like Receptor 4/MD-2 Complex Signaling in a Different Manner. J. Leukoc. Biol. 2012, 91, 967–976. [Google Scholar] [CrossRef]
- Schröfelbauer, B.; Raffetseder, J.; Hauner, M.; Wolkerstorfer, A.; Ernst, W.; Szolar, O.H.J. Glycyrrhizin, the Main Active Compound in Liquorice, Attenuates pro-Inflammatory Responses by Interfering with Membrane-Dependent Receptor Signalling. Biochem. J. 2009, 421, 473–482. [Google Scholar] [CrossRef]
- Tu, J.-H.; He, Y.-J.; Chen, Y.; Fan, L.; Zhang, W.; Tan, Z.-R.; Huang, Y.-F.; Guo, D.; Hu, D.-L.; Wang, D.; et al. Effect of Glycyrrhizin on the Activity of CYP3A Enzyme in Humans. Eur. J. Clin. Pharmacol. 2010, 66, 805–810. [Google Scholar] [CrossRef]
- Feng, J.; Yao, W.; Zhang, Y.; Xiang, A.P.; Yuan, D.; Hei, Z. Intravenous Anesthetics Enhance the Ability of Human Bone Marrow-Derived Mesenchymal Stem Cells to Alleviate Hepatic Ischemia-Reperfusion Injury in a Receptor-Dependent Manner. Cell Physiol. Biochem. 2018, 47, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Eom, Y.W.; Shim, K.Y.; Baik, S.K. Mesenchymal Stem Cell Therapy for Liver Fibrosis. Korean J. Intern. Med. 2015, 30, 580–589. [Google Scholar] [CrossRef]
- Gazdic, M.; Arsenijevic, A.; Markovic, B.S.; Volarevic, A.; Dimova, I.; Djonov, V.; Arsenijevic, N.; Stojkovic, M.; Volarevic, V. Mesenchymal Stem Cell-Dependent Modulation of Liver Diseases. Int. J. Biol. Sci. 2017, 13, 1109–1117. [Google Scholar] [CrossRef]
- Sakaida, I.; Terai, S.; Yamamoto, N.; Aoyama, K.; Ishikawa, T.; Nishina, H.; Okita, K. Transplantation of Bone Marrow Cells Reduces CCl4-Induced Liver Fibrosis in Mice. Hepatology 2004, 40, 1304–1311. [Google Scholar] [CrossRef]
- Efimenko, A.; Starostina, E.; Kalinina, N.; Stolzing, A. Angiogenic Properties of Aged Adipose Derived Mesenchymal Stem Cells after Hypoxic Conditioning. J. Transl. Med. 2011, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Gui, Y.; Yeganeh, M.; Ramanathan, S.; Leblanc, C.; Pomerleau, V.; Ferbeyre, G.; Saucier, C.; Ilangumaran, S. SOCS1 Controls Liver Regeneration by Regulating HGF Signaling in Hepatocytes. J. Hepatol. 2011, 55, 1300–1308. [Google Scholar] [CrossRef]
- Galland, S.; Stamenkovic, I. Mesenchymal Stromal Cells in Cancer: A Review of Their Immunomodulatory Functions and Dual Effects on Tumor Progression. J. Pathol. 2020, 250, 555–572. [Google Scholar] [CrossRef]
- Miceli, V.; Bulati, M.; Iannolo, G.; Zito, G.; Gallo, A.; Conaldi, P.G. Therapeutic Properties of Mesenchymal Stromal/Stem Cells: The Need of Cell Priming for Cell-Free Therapies in Regenerative Medicine. Int. J. Mol. Sci. 2021, 22, 763. [Google Scholar] [CrossRef] [PubMed]
- Aithal, A.P.; Bairy, L.K.; Seetharam, R.N. Safety Assessment of Human Bone Marrow-Derived Mesenchymal Stromal Cells Transplantation in Wistar Rats. J. Clin. Diagn. Res. 2017, 11, FF01–FF03. [Google Scholar] [CrossRef]
- Cheng, C.-Y.; Ho, T.-Y.; Lee, E.-J.; Su, S.-Y.; Tang, N.-Y.; Hsieh, C.-L. Ferulic Acid Reduces Cerebral Infarct through Its Antioxidative and Anti-Inflammatory Effects Following Transient Focal Cerebral Ischemia in Rats. Am. J. Chin. Med. 2008, 36, 1105–1119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, Z.; Lai, X.; Tu, W. Differentiation of Human Bone Marrow Stromal Cells into Neural-like Cells Induced by Sodium Ferulate in Vitro. Cell Mol. Immunol. 2005, 2, 225–229. [Google Scholar] [PubMed]
- Chen, J.; Zhang, Z.G.; Li, Y.; Wang, L.; Xu, Y.X.; Gautam, S.C.; Lu, M.; Zhu, Z.; Chopp, M. Intravenous Administration of Human Bone Marrow Stromal Cells Induces Angiogenesis in the Ischemic Boundary Zone after Stroke in Rats. Circ. Res. 2003, 92, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Zhang, R.; Katakowski, M.; Gautam, S.C.; Xu, Y.; Lu, M.; Zhang, Z.; Chopp, M. Combination Therapy of Stroke in Rats with a Nitric Oxide Donor and Human Bone Marrow Stromal Cells Enhances Angiogenesis and Neurogenesis. Brain Res. 2004, 1005, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Gabryel, B.; Trzeciak, H.I. Role of Astrocytes in Pathogenesis of Ischemic Brain Injury. Neurotox. Res. 2001, 3, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Kowiański, P.; Lietzau, G.; Steliga, A.; Waśkow, M.; Moryś, J. The Astrocytic Contribution to Neurovascular Coupling--Still More Questions than Answers? Neurosci. Res. 2013, 75, 171–183. [Google Scholar] [CrossRef]
- Girouard, H.; Iadecola, C. Neurovascular Coupling in the Normal Brain and in Hypertension, Stroke, and Alzheimer Disease. J. Appl. Physiol. 2006, 100, 328–335. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Sequence (5′–3′) |
|---|---|---|
| NF-kB | Forward | GCAGCACTACTTCTTGACCACC |
| Reverse | TCTGCTCCTGAGCATTGACGTC | |
| IL-1β | Forward | CCACAGACCTTCCAGGAGAATG |
| Reverse | GTGCAGTTCAGTGATCGTACAGG | |
| Casp9 | Forward | AGCCAGATGCTGTCCCATAC |
| Reverse | CAGGAGACAAAACCTGGGAA | |
| B-actin | Forward | GTGACATCCACACCCAGAGG |
| Reverse | ACAGGATGTCAAAACTGCCC |
| Caption | GSH (m. mol/mg/Protein) | SOD (u/mg/Protein) |
|---|---|---|
| Control | 4.49 ± 0.43 a | 4.15 ± 0.50 a |
| Cisplatin | 3.55 ± 0.51 a,b | 0.47 ± 0.10 d |
| GGE + Cisplatin | 0.87 ± 0.19 d | 1.29 ± 0.31 c,d |
| BM-MSCs + Cisplatin | 2.51 ± 0.48 b,c | 2.15 ± 0.23 c |
| BM-MSCs + GGE + Cisplatin | 1.36 ± 0.44 c,d | 3.16 ± 0.23 b |
| p-Value | <0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mobasher, M.A.; Ahmed, E.I.; Hakami, N.Y.; Germoush, M.O.; Awad, N.S.; Khodeer, D.M. The Combined Effect of Licorice Extract and Bone Marrow Mesenchymal Stem Cells on Cisplatin-Induced Hepatocellular Damage in Rats. Metabolites 2023, 13, 94. https://doi.org/10.3390/metabo13010094
Mobasher MA, Ahmed EI, Hakami NY, Germoush MO, Awad NS, Khodeer DM. The Combined Effect of Licorice Extract and Bone Marrow Mesenchymal Stem Cells on Cisplatin-Induced Hepatocellular Damage in Rats. Metabolites. 2023; 13(1):94. https://doi.org/10.3390/metabo13010094
Chicago/Turabian StyleMobasher, Maysa A., Eman Ibrahim Ahmed, Nora Y. Hakami, Mousa O. Germoush, Nabil S Awad, and Dina M. Khodeer. 2023. "The Combined Effect of Licorice Extract and Bone Marrow Mesenchymal Stem Cells on Cisplatin-Induced Hepatocellular Damage in Rats" Metabolites 13, no. 1: 94. https://doi.org/10.3390/metabo13010094
APA StyleMobasher, M. A., Ahmed, E. I., Hakami, N. Y., Germoush, M. O., Awad, N. S., & Khodeer, D. M. (2023). The Combined Effect of Licorice Extract and Bone Marrow Mesenchymal Stem Cells on Cisplatin-Induced Hepatocellular Damage in Rats. Metabolites, 13(1), 94. https://doi.org/10.3390/metabo13010094