Abstract
Leishmaniasis is a group of infectious diseases caused by Leishmania protozoa. The ineffectiveness, high toxicity, and/or parasite resistance of the currently available antileishmanial drugs has created an urgent need for safe and effective leishmaniasis treatment. Currently, the molecular-docking technique is used to predict the proper conformations of small-molecule ligands and the strength of the contact between a protein and a ligand, and the majority of research for the development of new drugs is centered on this type of prediction. Leishmania N-myristoyltransferase (NMT) has been shown to be a reliable therapeutic target for investigating new anti-leishmanial molecules through this kind of virtual screening. Natural products provide an incredible source of affordable chemical scaffolds that serve in the development of effective drugs. Withania somnifera leaves, roots, and fruits have been shown to contain withanolide and other phytomolecules that are efficient anti-protozoal agents against Malaria, Trypanosoma, and Leishmania spp. Through a review of previously reported compounds from W. somnifera-afforded 35 alkaloid, phenolic, and steroid compounds and 132 withanolides/derivatives, typical of the Withania genus. These compounds were subjected to molecular docking screening and molecular dynamics against L. major NMT. Calycopteretin-3-rutinoside and withanoside IX showed the highest affinity and binding stability to L. major NMT, implying that these compounds could be used as antileishmanial drugs and/or as a scaffold for the design of related parasite NMT inhibitors with markedly enhanced binding affinity.
1. Introduction
Leishmaniasis is a serious neglected tropical disease that causes several illnesses connected to immune-system dysfunction and poverty, which are sometimes associated with high fatality rates [1]. The disease is manifested in three different ways: Cutaneous leishmaniasis (CL), which is characterized by lesions at the site of infection; mucocutaneous leishmaniasis, which is characterized by invasion and destruction of the mucosa; and visceral leishmaniasis, which is the most severe form because the infection spreads to other organs like the spleen and liver [2]. Each year, around two million new cases are reported worldwide, with cutaneous leishmaniasis accounting for 75% of these infections. The primary cause of cutaneous leishmaniasis is Leishmania major, and out of the 89 countries where it is present, Afghanistan, Brazil, Iran, Peru, Saudi Arabia, and Syria account for 90% of CL cases [1,3]. Despite the long history of the disease, no vaccine is available, and all of the current medications are either ineffective, have unfavorable side effects, or are losing efficacy as resistance emerges [4].
To produce a new anti-Leishmania drug, long isolation, purification, or synthesis processes, as well as excessively expensive in vitro and in vivo biological evaluations, are required [5]. Thus, the present challenge is to identify novel molecules with potential antileishmanial activity while conserving time and money. Utilizing in silico techniques can shorten the time and lower the cost associated with developing new medications [6].
Molecular docking is an in silico technique used to identify correct conformations of small-molecule ligands and estimate the strength of the protein–ligand interaction [7]. Presently, the majority of research for the development of novel drugs focuses on this kind of analysis. Molecular docking has been employed in multiple studies to create new antiparasitic medications while considering a variety of targets to find new Leishmania treatments [8]. Discovering new leishmaniasis treatments is made more enticing by molecules implicated in parasite-specific metabolic processes [9]. The affection of this kind of target implies the death of the parasite and the control of the infection [5]. Worth mentioning is that standard chemotherapeutic antileishmanial drugs operate via related mechanisms—namely, interference in parasite metabolic processes (Table 1) [10]. Targeting specific molecular pathways is a common approach in rational drug design and discovery for developing such leishmaniasis-treating compounds. Pteridine reductase, trypanothione reductase, N-myristoyltransferase (NMT), trypanothione synthetase, inosine-uridine nucleoside hydrolase, and topoisomerases are just a few of the more than 21 potential therapeutic targets of antileishmanial drug discovery that have been reported in the literature and gathered in a book chapter [9].

Table 1.
Standard antileishmanial chemotherapeutic drugs and their suggested mode of antileishmanial effect.
N-myristoyltransferase is an example of the molecules implicated in parasite metabolic processes. It is an enzyme that catalyzes the co-translational transfer of a C14 fatty acid from myristoyl-CoA onto the N-terminal glycine residue of a significant subset of proteins [11]. N-myristoylation is believed to play a crucial role in the correct cellular localization and biological function of such modified proteins. Since 1997, it has been established that Leishmania major contains NMT [12]. Later, it was determined that Leishmania sp. needed the NMT enzyme to survive. It has been extensively studied in some organisms, including Trypanosoma brucei, T. cruzi, and Leishmania spp. [13,14,15,16,17]. In a high-throughput screening campaign run by Pfizer, four different scaffolds, aminoacylpyrrolidine (PF-03402623), piperidinylindole (PF-03393842), and thienopyrimidine (PF-003494M), together with a subsequently created fused hybrid compound, 43, were discovered to be highly effective inhibitors of the Labrus donovani NMT (Figure 1) [18]. A different experiment on a collection of 1600 pyrazolyl sulfonamide compounds led to the discovery of compound 2, a potent inhibitor of Leishmania major NMT with negligible activity against Leishmania donovani intracellular amastigotes [5]. Furthermore, several publications and patents indicated the existence of additional NMT inhibitor scaffolds, including pyrrolidines, piperidinylindoles, azetidinopyrimidines, aminomethylindazoles, benzimidazoles, thienopyrimidines, biphenyl derivatives, benzofuranes, benzothiophenes, oxadiazoles, (pyrazolomethyl)-1,3,4-oxadiazoles, and thienopyrimidine [19,20,21,22].
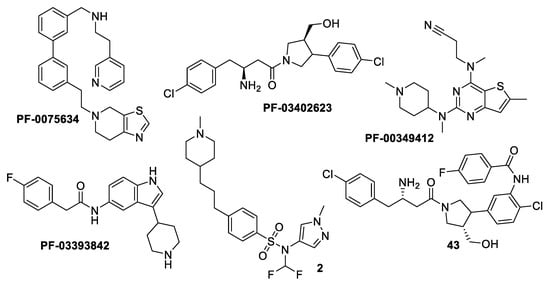
Figure 1.
Structures of examples of NMT inhibitors with antileishmanial activity [9].
N-myristoyltransferase inhibitor compounds from medicinal plants may be useful in the quest for alternatives to current treatments for CL. Withania somnifera (L.) Dunal (Solanaceae), also known as Ashwagandha or Indian ginseng, has been used as a traditional herb against a plethora of human medical conditions [23]. It is one of the most extensively used plants in the Unani and Ayurvedic systems of medicine. It has been reported as an important source of withanolides, alkaloids, steroids, flavonoids, nitrogen-containing compounds, and others [23]. Among them, withanolides are a class of highly oxygenated steroids generated from a C28 ergostane skeleton (Figure 2). They are marker compounds characteristic of Solanaceae plants, particularly those of the genus Withania [23]. They are thought to be responsible for the majority of W. somnifera bioactivity [16]. W. somnifera happens to be one of the prime examples of Rasayana, a branch of Ayurvedic science, a medicinal plant that possesses immunomodulation, anti-cancer, anti-depressant, and neuroprotective properties; promotes the body’s resistance to diseases; increases strength and intellect; and delays aging; as well as other biological properties [23]. Closer examination of pertinent in vitro and in vivo studies revealed that W. somnifera extracts molecules’ significant bioactivity against several metabolic, reproductive, cardiovascular, neurological, and psychological conditions. Additionally, it has been shown to have antibacterial and antiparasitic properties against Trypanosoma, Leishmania species, and Malaria [24]. The alleged value of W. somnifera in treating leishmaniasis, as proposed by earlier research, is highlighted in the following themes.
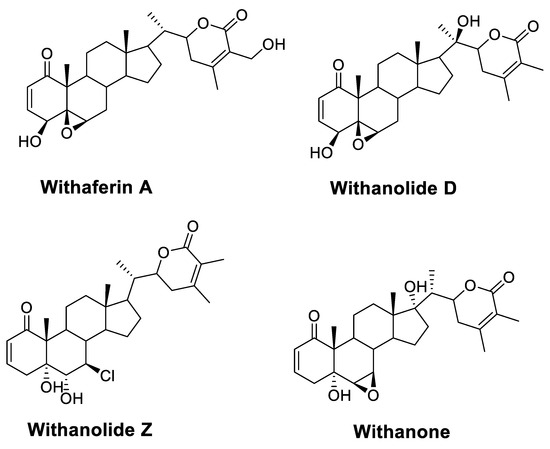
Figure 2.
Structural features of bioactive withanolides of W. somnifera represented by withaferin A, withanolides D and Z, and withanone.
1.1. Overview of the Antileishmanial Properties of Withania somnifera
Screening of W. somnifera from different geographical regions has shown that methanolic extract exhibits in vitro antileishmanial action against free-living promastigotes [25,26] and intracellular amastigotes of L. major [26]. Investigation of a solvent-soluble fraction of root and fruit hydromethanolic extracts demonstrated significant antileishmanial promastigote properties of the butanol-soluble fraction from roots and fruits, whereas in vitro growth-inhibitory assessment on axenic amastigotes revealed promising activity of the root ethyl acetate-soluble and butanol-soluble fractions [1]. Further bio-guided fractionation revealed that the withanolide-enriched fraction from W. somnifera ethanolic extract was effective [17].
Studies have shown that pure withaferin-A, a prominent withanolide in W. somnifera, possesses antileishmanial activity. It inhibits protein kinase C, which allows the apoptotic topoisomerase I-DNA complex to induce apoptosis [25,27]. Additionally, withanolide Z, a chlorinated withanolide, has been reported to exert an inhibitory effect against L. donovani topoisomerase-1 [28].
The anti-leishmanial activity of the withanolide-enriched extract and pure withanolide of W. somnifera was found to be mediated by the induction of morphological alterations from a spindle to round shape and the loss of flagella and cell integrity in promastigotes. They also induce apoptosis-like cellular death in L. donovani by inducing DNA nicks, apoptosis, and cell-cycle arrest in a dose- and time-dependent manner, actions that were mediated by increasing reactive oxygen species (ROS) production and decreasing mitochondrial potential [29]. Additionally, these withanolide-enriched fractions and pure withanolide of W. somnifera, alone or in combination with other herbal products or standard anti-Leishmanial drugs, were found to modulate hamsters’ immunological response to infection with L. donovani.
1.2. Immunomodulatory Effects of W. somnifera in Leishmaniasis Infections
One of the immunopathological consequences of active visceral leishmaniasis is the suppression of protective T-helper (Th)-1 cells and the induction of disease-promoting Th-2 cells [30]. Therefore, host immunomodulation is crucial for the treatment of visceral leishmaniasis. In animal-model research, W. somnifera chemotype NMITLI-101 R extract, withaferin A, and the chemotype NMITLI-101 R extract in combination with an ED50 dose of miltefosine were investigated for their immunoprotective and therapeutic effects against L. donovani infection. It has been observed that the efficacy of W. somnifera was linked to the compelling Th1 immune responses driven by interferon-gamma (IFN-γ) and interleukin-12 (IL-12), as well as dramatically reduced levels of Th2 cytokines (IL-4, IL-10) and transforming growth factor beta (TGF-β). Meanwhile, they significantly increased the levels of NO production, ROS creation, and Leishmania-specific IgG2 antibodies, along with profoundly delayed-type hypersensitivity (DTH) and strong T-cell responses [31,32,33,34].
In another study, when Asparagus racemosus and W. somnifera were used to treat infected mice, the parasite burden was successfully reduced and protective Th1-type immune responses were elicited, resulting in the normalization of biochemical and hematological parameters [30]. In the treatment of visceral leishmaniasis, the combination of W. somnifera extract with cisplatin led to a notable selective elevation of the Th1 type of immunity, verified immunomodulatory action, and a protective impact against the adverse effects of cisplatin on multiple bodily organs. The percentage of CD4 and CD8 T-lymphocytes, as well as the natural killer (NK) cell-associated marker NK1, increased significantly [33].
1.3. Molecular Docking Studies in the Development of Antileishmanial Drugs
According to molecular modeling and dynamic investigations of the leishmanial protein kinase C structure, withaferin A and withanone were suggested to disrupt the protein kinase C (PKC) pathway [23,35]. Molecular-docking studies of the binding mode of withaferin-A with pteridine reductase 1 (PTR1), which is involved in pteridine salvage, a crucial enzyme for parasite proliferation, revealed that withaferin-A inhibits PTR-1 enzymes through the uncompetitive mode of inhibition in the parasites.
In summary, in rational antileishmanial drug design and discovery, targeting particular biochemical pathways was found to be a typical strategy for creating leishmaniasis-treating molecules. In order to screen molecules from both natural and synthetic sources, more than 21 molecular targets were used [9]. The structure-guided creation of new lead compounds discovered in high-throughput screening efforts aimed at L. major and L. donovani NMT has led to the identification of effective inhibitors [9,36]. These newly discovered Leishmania NMT inhibitors did not possess the same cellular activity as the enzyme against Leishmania donovani axenic amastigotes, a fact that has been explained by the restricted cellular absorption related to the basic nature of the compounds [19].
W. somnifera’s wide variety of the non-basic withanolide derivative and phenolics would be an important source of new hits of Leishmania NMT inhibitors with higher cellular activity against Leishmania parasites. Thus, the goals of this study are to highlight W. somnifera anti-leishmanial properties as well as to discover new L. major NMT inhibitors using in silico molecular-docking and molecular-dynamics analysis of W. somnifera metabolites as potential anti-L. major medicine.
2. Experimental
2.1. Phytochemical Review and Data Collection
A thorough review of the previously reported compounds from W. somnifera (L.) Dunal in peer-reviewed research papers and international databases, including Science Direct, Pubchem, Google Scholar, SciFinder, etc., afforded a total of 167 different phytoconstituents, which were considered for the present study.
2.2. Molecular-Docking Simulation
PyRx software was used for the docking experiments [37]. The RCSB Protein Data Bank (https://www.rcsb.org/ accessed on 20 March 2022) was used to retrieve the three-dimensional (3D) structure of L. major N-myristoyltransferase (NMT) in complex with the thienopyrimidine inhibitor IMP-0000096 (PDB ID: 6QDF) (Figure 3) determined by X-RAY diffraction (Resolution: 1.49 Å) as a target for molecular-docking studies [21]. The 3D chemical structures of the selected molecules were retrieved from PubChem, with polar hydrogen added, partial charge corrected, and energy minimized using the Merck molecular force field (MMFF94x). Molecular-docking analysis was carried out via flexible ligand-fixed receptor-docking parameters using an active complexed ligand active site. The most stable affinity-binding interactions were selected based on the best pose scores. The docking scores and 2D and 3D interactions were recorded [38,39]. BIOVIA Discovery Studio (v21.1.0.20298) was used for 2D and 3D interaction visualization [40].
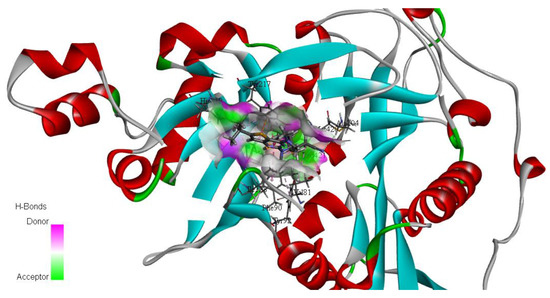
Figure 3.
3D structure of L. major NMT in complex with the thienopyrimidine inhibitor IMP-0000096 (PDB ID: 6QDF).
2.3. Molecular-Dynamics Simulation
Molecular-dynamics simulations (MDS) for the best three generated ligand–enzyme complexes were performed using the Nanoscale Molecular Dynamics (NAMD) 3.0 software, applying the CHARMM27 force field, and the MDS was continued for 100 ns following the previously described protocol [41]. The trajectory was stored every 0.1 ns and further analyzed with the VMD 1.9 software. The Molecular Mechanics Poisson–Boltzmann Surface Area (MMPBSA) embedded in the MMPBSA.py module of AMBER18 was utilized to calculate the binding free energy of the docked complex [42]. One hundred frames were processed from the trajectories in total, and the binding free energy was estimated using the following equation:
ΔGBinding = ΔGComplex − ΔGReceptor − ΔGInhibitor
Each of the aforementioned terms requires the calculation of multiple energy components, including van der Waals energy, electrostatic energy, internal energy from molecular mechanics, and a polar contribution to solvation energy.
3. Results
Earlier phytochemical research on W. somnifera led to the isolation of many phytochemical compounds. According to their chemical structures and the number of isolates, to the best of our knowledge, it could be distinguished as 10 alkaloids, 15 phenolic compounds, 10 sterols, 6 withanones, 6 chloro-containing withanolides, 5 sulfur-containing withanolides, 9 withanamide, 87 withanolides, and 19 withanosides. To discover potential hits that could be used as scaffolds for developing antileishmanial drug candidates, we evaluated these compounds against the Leishmania major NMT utilizing in silico molecular docking and molecular dynamics. The compounds are sorted according to their phytochemical groups parallel to their docking results in descending order according to pose score in Table 2.
Table 2.
List of the compounds isolated from W. somnifera along with their molecular docking results.
4. Discussion
Cutaneous leishmaniasis, caused by more than 20 species of Leishmania parasites, is a derelict tropical disease endemic in most world nations with high incidence rates. In the Kingdom of Saudi Arabia, CL is a significant public-health issue because of several risk factors, including rapid population growth and migration. The disease is endemic in many parts of the kingdom, and L. major and L. tropica are the most commonly detected species [123]. Current chemotherapeutic drugs used for leishmaniasis treatment are discouraging due to associated toxicity, a rase of drug resistance, and high cost. The development of new medications with enough safety and affordability is immediately needed to tackle this disease. W. somnifera is a perennial shrub found in open fields and deserts from the Mediterranean region to Southeast Asia [1]. A reasonable level of safety has been supported by up to 30 clinical studies; there were no obvious side effects or changes in hematological, biochemical, or vital indicators. Pre-clinical studies on chronic toxicity that lasted up to 8 months [16] further supported the safety. As stated above, both in vitro and in vivo tests were used to determine the antileishmanial activity of various organ extracts and sub-extracts of W. somnifera. However, only a small subset of metabolites was tested for antileishmanial activity. Together, these factors make it more important than ever to thoroughly research the phytoconstituents of the plant as possible sustainable and affordable antileishmanial remedies. Due to the economic issue and the hazards of handling a live Leishmania parasite in laboratory work, we adopted computational analysis, molecular docking, and molecular dynamics as a quick and cost-effective scheme to identify potential anti-L. major compounds from W. somnifera.
N-Myristoyltransferase is a prevalent essential enzyme in all Leishmania species [21]. NMT catalyzes the binding of myristate to the amino-terminal glycine residue of a subset of eukaryotic proteins involved in various cellular processes, such as vesicular protein trafficking and signal transduction. NMT has been demonstrated to be essential for viability by classical gene knockout and RNA interference, suggesting that this enzyme has the potential as a target for drug development. It has been proven that NMT is a valid therapeutic target for the treatment of fungus and parasite infections [15,124]. The discovery of the NMT crystal structure establishes a method for structural analysis of inhibitor complexes and structure-assisted drug discovery [21]. Amazingly, it has been demonstrated that the inhibitors only operate on the host enzyme despite the high degree of conservation between the active regions of the parasite and human NMTs [125]. According to the determined structure of β-sheet in the C-terminal domain and protein loops, the residues that are predicted to be the targeted inhibitor site in the NMT are Tyr-80, Val-81, Glu-82, Phe-88, Phe-90, Tyr-92, Asn1-67, Thr-203, Tyr-217, His-219, Phe-232, Tyr-326, Ile-328, Ser-330, Leu-341, Ala-343, Tyr-345, Val-374, Asn-376, Asp-396, Leu-399, Met-420, and Leu421 [126].
As a first step toward utilizing one or more of them as an effective CL medication, the wide range of previously reported compounds from W. somnifera species was chosen for virtual screening as L. major NMT inhibitors. In total, 167 compounds were studied for their binding affinity as NMT enzyme inhibitors using molecular-docking simulation (Table 2). The results revealed a wide range of compounds’ affinity to the NMT enzyme (−9.0 to −24.0 kcal/mol).
The compounds with a score of less than −18.0 kcal/mol are thought to be the most active. To evaluate the experimental stability of the docked ligand conformers, these compounds were filtered by RMSD value with a cutoff of 1.7 Å [127]. The top-scoring compounds were calycopteretin-3-rutinoside, withanamide F, withanolide A, (4β,5β,6β,17α,22R) 5,6-Epoxy-4,17,27-trihydroxy-1-oxowitha-2,24-dienolide, withanoside II, withanoside X, physagulin D (1→6)-β-D-glucopyranosyl-(1→4)-β-D-glucopyranoside, withanoside IX, sitoindoside IX, 4,16-dihydroxy-5β,6β-epoxyphysagulin D, and 24,25-dihydrowithanoside VI. These compounds showed the highest affinity for the NMT active site within the selected parameter range (Figure 4). 4,16-Dihydroxy-5β,6β-epoxyphysagulin D, calycopteretin 3-rutinoside, and withanoside IX revealed the most potent affinity to the receptor active site with −24.0, −23.3, and −22.2 kcal/mol, respectively.
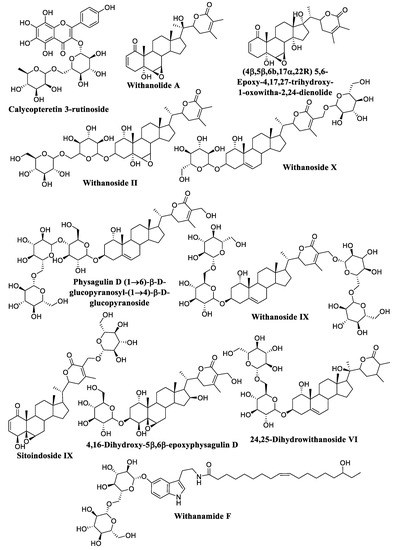
Figure 4.
The structures of top-scoring compounds.
Targeting with a pose score of −24.0 kcal/mol and an RSMD value of 1.07, 4,16-dihydroxy-5,6-epoxyphysagulin D demonstrated the highest affinity to the selected enzyme. Hydrogen bonds were formed between the compound and Met-420 and Asn-167 as H-donors, as well as interactions with Asn-383, Tyr-80, Val-378, and Asn-167 as H-acceptors (Figure 5).
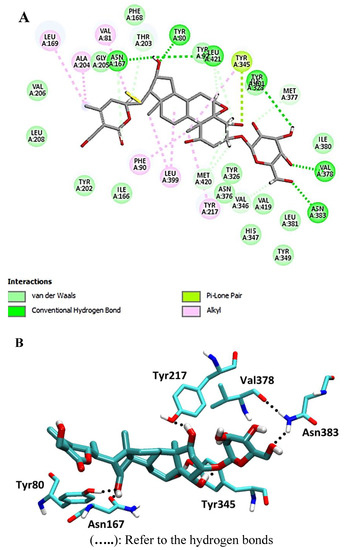
Figure 5.
2D and 3D interaction complex of 4,16-dihydroxy-5,6-epoxyphysagulin D with NMT ((A) and (B), respectively).
The flavonoid glycoside calycopteretin 3-rutinoside was found to have a potential affinity with a pose score value of −23.3 kcal/mol (RSMD = 1.24 Å) with a hydrogen-bond interaction with Met-420 and Leu-421 as H-donor in addition to Tyr-80 as H-acceptor (Figure 6).
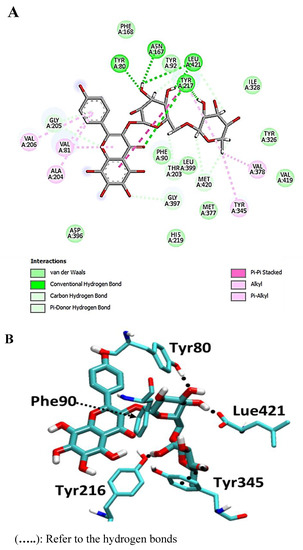
Figure 6.
2D and 3D interactions complex of calycopteretin 3-rutinoside with NMT ((A) and (B), respectively).
Furthermore, the withanolide glycoside withanoside IX formed hydrogen bonds with Glu-82, Asp-83, Val-346, Val-346, and Met-420, yielding a binding score of −22.2 kcal/mol (RSMD = 1.41; Figure 7). Other hydrophobic interactions were found in all interacted compounds, as shown in Figure 7.
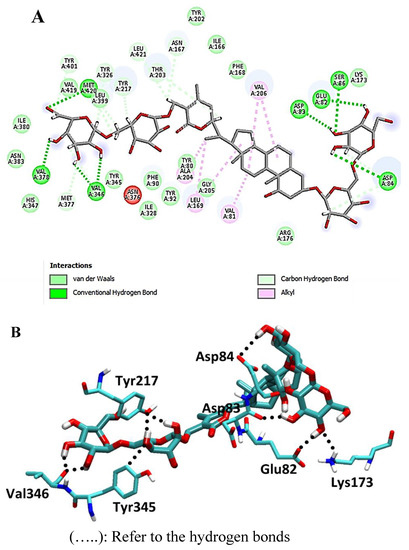
Figure 7.
2D and 3D interaction complex of withanoside IX with NMT ((A) and (B), respectively).
Compared with the pose score of the complexed thienopyrimidine inhibitor IMP-0000096 (−14.2 kcal/mol), these compounds could be promising antileishmanial scaffolds.
The introduction of promastigote into the body, the blood circulation of amastigote, phagocytosis and macrophage cell growth, lysis, and blood flow as amastigote once more are among the steps that occur in Leishmania infection [1]. However, NMT inhibitors obtained by synthesis from various research groups were unable to kill parasites inside macrophages at reasonable therapeutic doses, as shown by in vivo assay [2]. On the other hand, withanolide-enriched extracts from W. somnifera reduced the intracellular parasite load by ~50% compared to the infected control [34]. Withaferin-A produced a dose-dependent decrease in parasite number inside the macrophages at concentrations of 0.5–1.5 µM [34]. These literature results, together with our findings from docking studies, suggest that the compounds with the top docking scores could be leishmanicidal agents that operate by inhibiting L. major NMT in the free promastigote and the intercellular amastigote forms. Another exciting outcome of our research is the potential to use these molecules as a promising framework for the development of novel leishmanial NMT inhibitors and possibly NMT inhibitors of other protozoa.
Molecular-Dynamics Simulation Study
To validate the docking results, we subjected the retrieved docking poses to a 100 ns-long molecular-dynamics simulation (MDS) run. As depicted in Figure 8, calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D achieved good binding stability inside NMT’s active site, showing average RMSDs (i.e., 1.1 Å, 2.2 Å, and 3.9 Å, respectively) that were comparable with that of the co-crystallized inhibitor (i.e., 1.4 Å). The global dynamic behavior of NMT did not show significant change upon binding with either the co-crystallized inhibitor or with the three selected ligands (i.e., calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D), where the RMSFs of either the ligand-free enzyme or the enzyme bound to the three ligands and the co-crystalized inhibitor showed good alignment (Figure 8B).
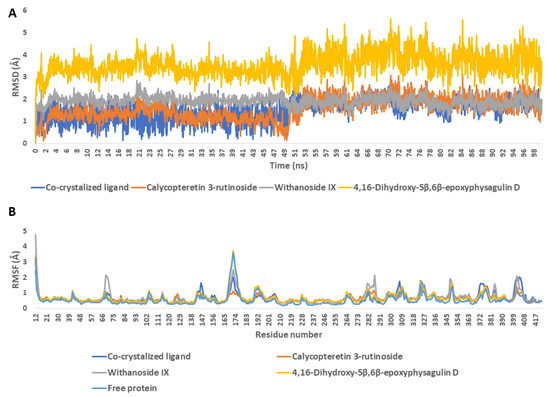
Figure 8.
(A) RMSDs of the compounds calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D along with the co-crystalized inhibitor inside the active site of NMT (PDB ID: 6QDF) throughout 100 ns MDS runs. (B) RMSFs of NMT in the absence and in the presence of calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D along with the co-crystalized inhibitor.
The frequency of H-bonds detected for each ligand along with the co-crystalized inhibitor averaged around two H-bonds (cut-off distance for H-bonds was set to 3.0 Å; Figure 9). In regard to the interaction energies (i.e., electrostatic + van der Waals energies) of each ligand, calycopteretin-3-rutinoside showed the worst average total interaction energy (−26.4 kcal/mol), whereas withanoside IX and 4,16-dihydroxy-5β,6β-epoxyphysagulin D, along with the co-crystalized inhibitor, showed convergent average total interaction energies (−55.5, −54.5, and −44.4 kcal/mol, respectively; Figure 10).
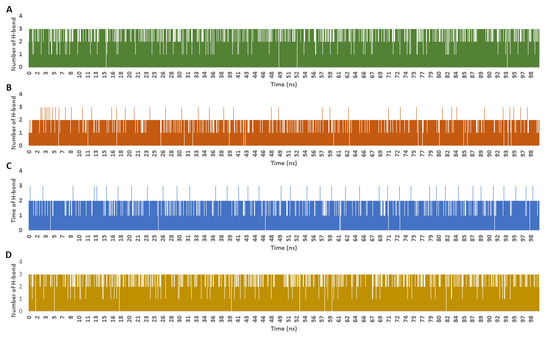
Figure 9.
Number of H-bonds detected for calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D along with the co-crystallized inhibitor inside the active site of NMT throughout 100 ns MDS runs ((A–D), respectively). The cut-off distance for H-bonds was set to 3.0 Å.
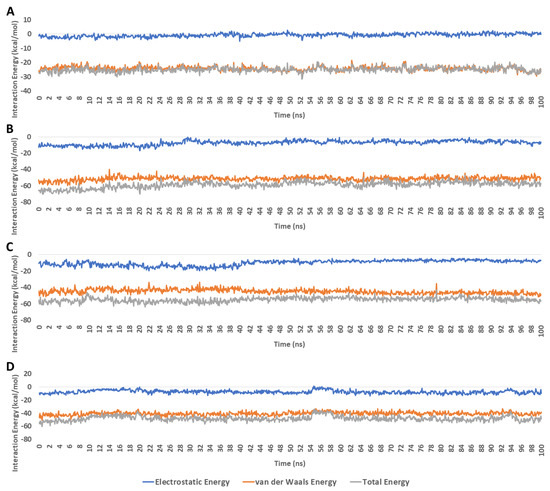
Figure 10.
Interaction energies of calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D along with the co-crystallized inhibitor inside the active site of NMT throughout 100 ns MDS runs (A–D), respectively). The total interaction energy is the sum of both electrostatic and van der Waals energies.
Accordingly, the calculated binding free energies (ΔGbinding) extracted from the MDS runs were also convergent, ranging from −7.3 kcal/mol for the co-crystallized inhibitor to −9.8 kcal/mol for 4,16-dihydroxy-5,6-epoxyphysagulin D, except for calycopteretin 3-rutinoside, which obtained the lowest negative value (Table 3).
Table 3.
Binding free energies (MM-PBSA) of top-scoring compounds along with the co-crystalized inhibitor in complex with NMT.
5. Conclusions
The urgent need to discover a new anti-L. major drug in a reasonable time and at a low cost encouraged us to consider in silico techniques to examine W. somnifera metabolites as an inhibitor of L. major NMT. Among the 167 virtually screened compounds, the phenolic glycoside calycopteretin-3-rutinoside, withanoside IX, and 4,16-dihydroxy-5β,6β-epoxyphysagulin D showed promising binding affinity towards Leishmania NMT. In light of the fact that the standard chemotherapeutic antileishmanial medications, such as antimonials, amphotericin B, miltefosine, paromomycin, and pentamidine, work by interfering with parasite metabolic processes, exploring new natural substances that are connected to parasite-specific metabolic processes, such as our discovered NMT inhibitors from W. somnifera, is extremely promising. The results of our investigation will help scientists design in vitro and preclinical animal studies employing W. somnifera metabolites, with an emphasis on the promising NMT inhibitors we have discovered.
Author Contributions
Conceptualization, M.A.A.O., M.E.A. and E.-S.A.-S.; methodology, M.M.A., A.M.S., K.A.S. and M.E.A.; software, A.M.S., K.A.S. and M.E.A.; validation, A.M.S., K.A.S. and M.E.A.; formal analysis, A.M.S., K.A.S. and M.E.A.; investigation, M.A.A.O., E.-S.A.-S., A.M.S., K.A.S. and M.E.A.; resources, M.M.A., A.M.S., K.A.S. and M.E.A.; data curation, M.M.A., A.M.S., K.A.S. and M.E.A.; writing—original draft preparation, M.M.A., M.A.A.O., E.-S.A.-S. and M.E.A.; writing—review and editing, M.A.A.O.; visualization, M.M.A., A.M.S., K.A.S. and M.E.A.; supervision, M.A.A.O.; project administration, M.A.A.O. and E.-S.A.-S.; funding acquisition, M.A.A.O. All authors have read and agreed to the published version of the manuscript.
Funding
The authors are thankful to the Deanship of Scientific Research at Najran University, Saudi Arabia, for funding this research under the National Research Priority Funding Program (NU/NRP/MRC/11/8).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data, which include the 2D and 3D structures of the compounds under investigation, are obtainable from the corresponding author upon request.
Acknowledgments
The authors are thankful to the Deanship of Scientific Research at Najran University, Saudi Arabia, for funding this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Orabi, M.A.; Zidan, S.A.; Sakagami, H.; Murakami, Y.; Ali, A.A.; Alyami, H.S.; Alshabi, A.M.; Matsunami, K. Antileishmanial and lung adenocarcinoma cell toxicity of Withania somnifera (Linn.) dunal root and fruit extracts. Nat. Prod. Res. 2022, 36, 4231–4237. [Google Scholar] [CrossRef]
- Brannigan, J.A.; Wilkinson, A.J. Drug discovery in leishmaniasis using protein lipidation as a target. Biophys. Rev. 2021, 13, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Riezk, A.; Raynes, J.G.; Yardley, V.; Murdan, S.; Croft, S.L. Activity of chitosan and its derivatives against Leishmania major and Leishmania mexicana in vitro. Antimicrob. Agents Chemother. 2020, 64, e01772-19. [Google Scholar] [CrossRef] [PubMed]
- Zidan, S.A.; Abdelhamid, R.A.; Alian, A.; Fouad, M.A.; Matsunami, K.; Orabi, M.A. Diterpenes and sterols from the Red Sea soft coral Sarcophyton trocheliophorum and their cytotoxicity and anti-leishmanial activities. J. Asian Nat. Prod. Res. 2022, 24, 794–802. [Google Scholar] [CrossRef]
- Corpas-Lopez, V.; Moniz, S.; Thomas, M.; Wall, R.J.; Torrie, L.S.; Zander-Dinse, D.; Tinti, M.; Brand, S.; Stojanovski, L.; Manthri, S. Pharmacological validation of N-myristoyltransferase as a drug target in Leishmania donovani. ACS Infect. Dis. 2018, 5, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.S.; Srinivas, K. Modern drug discovery process: An in silico approach. J. Bioinform. Seq. Anal. 2011, 2, 89–94. [Google Scholar]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef]
- Alamzeb, M.; Ali, S.; Mamoon-Ur-Rashid; Khan, B.; Ihsanullah; Adnan; Omer, M.; Ullah, A.; Ali, J.; Setzer, W.N. Antileishmanial Potential of Berberine Alkaloids from Berberis glaucocarpa Roots: Molecular Docking Suggests Relevant Leishmania Protein Targets. Nat. Prod. Commun. 2021, 16, 1934578X211031148. [Google Scholar] [CrossRef]
- Istanbullu, H.; Bayraktar, G. Toward New Antileishmanial Compounds: Molecular Targets for Leishmaniasis Treatment. In Leishmaniasis—General Aspects of a Stigmatized Disease; de Azevedo Calderon, L., Ed.; IntechOpen: London, UK, 2021; p. 256. [Google Scholar]
- Hassan, A.A.; Khalid, H.E.; Abdalla, A.H.; Mukhtar, M.M.; Osman, W.J.; Efferth, T. Antileishmanial Activities of Medicinal Herbs and Phytochemicals In Vitro and In Vivo: An Update for the Years 2015 to 2021. Molecules 2022, 27, 7579. [Google Scholar] [CrossRef]
- Wright, M.H.; Paape, D.; Storck, E.M.; Serwa, R.A.; Smith, D.F.; Tate, E.W. Global analysis of protein N-myristoylation and exploration of N-myristoyltransferase as a drug target in the neglected human pathogen Leishmania donovani. Chem. Biol. 2015, 22, 342–354. [Google Scholar] [CrossRef] [PubMed]
- McKean, P.G.; Delahay, R.; Pimenta, P.F.; Smith, D.F. Characterisation of a second protein encoded by the differentially regulated LmcDNA16 gene family of Leishmania major. Mol. Biochem. Parasitol. 1997, 85, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Towler, D.; Eubanks, S.; Towery, D.; Adams, S.; Glaser, L. Amino-terminal processing of proteins by N-myristoylation. Substrate specificity of N-myristoyl transferase. J. Biol. Chem. 1987, 262, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Frearson, J.A.; Brand, S.; McElroy, S.P.; Cleghorn, L.A.; Smid, O.; Stojanovski, L.; Price, H.P.; Guther, M.L.S.; Torrie, L.S.; Robinson, D.A. N-myristoyltransferase inhibitors as new leads to treat sleeping sickness. Nature 2010, 464, 728–732. [Google Scholar] [CrossRef]
- Price, H.P.; Menon, M.R.; Panethymitaki, C.; Goulding, D.; McKean, P.G.; Smith, D.F. Myristoyl-CoA: Protein N-myristoyltransferase, an essential enzyme and potential drug target in kinetoplastid parasites. J. Biol. Chem. 2003, 278, 7206–7214. [Google Scholar] [CrossRef] [PubMed]
- Glover, C.J.; Hartman, K.D.; Felsted, R.L. Human N-myristoyltransferase amino-terminal domain involved in targeting the enzyme to the ribosomal subcellular fraction. J. Biol. Chem. 1997, 272, 28680–28689. [Google Scholar] [CrossRef] [PubMed]
- Herrera, L.J.; Brand, S.; Santos, A.; Nohara, L.L.; Harrison, J.; Norcross, N.R.; Thompson, S.; Smith, V.; Lema, C.; Varela-Ramirez, A. Validation of N-myristoyltransferase as potential chemotherapeutic target in mammal-dwelling stages of Trypanosoma cruzi. PLoS Negl. Trop. Dis. 2016, 10, e0004540. [Google Scholar] [CrossRef]
- Hutton, J.A.; Goncalves, V.; Brannigan, J.A.; Paape, D.; Wright, M.H.; Waugh, T.M.; Roberts, S.M.; Bell, A.S.; Wilkinson, A.J.; Smith, D.F. Structure-based design of potent and selective Leishmania N-myristoyltransferase inhibitors. J. Med. Chem. 2014, 57, 8664–8670. [Google Scholar] [CrossRef]
- Åsberg, P.; Hammer, K.; Olsson, J.; Henriksson, M. Novel Compounds and Their Use in Therapy. U.S. Patent No WO2013009259, 17 January 2013. [Google Scholar]
- Rackham, M.D.; Yu, Z.; Brannigan, J.A.; Heal, W.P.; Paape, D.; Barker, K.V.; Wilkinson, A.J.; Smith, D.F.; Leatherbarrow, R.J.; Tate, E.W. Discovery of high affinity inhibitors of Leishmania donovani N-myristoyltransferase. MedChemComm 2015, 6, 1761–1766. [Google Scholar] [CrossRef]
- Bell, A.S.; Yu, Z.; Hutton, J.A.; Wright, M.H.; Brannigan, J.A.; Paape, D.; Roberts, S.M.; Sutherell, C.L.; Ritzefeld, M.; Wilkinson, A.J. Novel thienopyrimidine inhibitors of Leishmania N-myristoyltransferase with on-target activity in intracellular amastigotes. J. Med. Chem. 2020, 63, 7740–7765. [Google Scholar] [CrossRef]
- Olaleye, T.O.; Brannigan, J.A.; Roberts, S.M.; Leatherbarrow, R.J.; Wilkinson, A.J.; Tate, E.W. Peptidomimetic inhibitors of N-myristoyltransferase from human malaria and leishmaniasis parasites. Org. Biomol. Chem. 2014, 12, 8132–8137. [Google Scholar] [CrossRef]
- Paul, S.; Chakraborty, S.; Anand, U.; Dey, S.; Nandy, S.; Ghorai, M.; Saha, S.C.; Patil, M.T.; Kandimalla, R.; Proćków, J. Withania somnifera (L.) Dunal (Ashwagandha): A comprehensive review on ethnopharmacology, pharmacotherapeutics, biomedicinal and toxicological aspects. Biomed. Pharmacother. 2021, 143, 112175. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.C.; Wadhwa, R. Science of Ashwagandha: Preventive and Therapeutic Potentials; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Sharma, U.; Velpandian, T.; Sharma, P.; Singh, S. Evaluation of anti-leishmanial activity of selected Indian plants known to have antimicrobial properties. Parasitol. Res. 2009, 105, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- El-On, J.; Ozer, L.; Gopas, J.; Sneir, R.; Enav, H.; Luft, N.; Davidov, G.; Golan-Goldhirsh, A. Antileishmanial activity in Israeli plants. Ann. Trop. Med. Parasitol. 2009, 103, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Banerjee, B.; Das, B.; Ganguly, A.; Sen, T.; Pramanik, S.; Mukhopadhyay, S.; Majumder, H. Apoptosis is induced in leishmanial cells by a novel protein kinase inhibitor withaferin A and is facilitated by apoptotic topoisomerase I–DNA complex. Cell Death Differ. 2007, 14, 358–367. [Google Scholar] [CrossRef]
- Pramanick, S.; Roy, A.; Ghosh, S.; Majumder, H.K.; Mukhopadhyay, S. Withanolide Z, a new chlorinated withanolide from Withania somnifera. Planta Med. 2008, 74, 1745–1748. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Dayakar, A.; Veronica, J.; Sundar, S.; Maurya, R. An in vitro study of apoptotic like death in Leishmania donovani promastigotes by withanolides. Parasitol. Int. 2013, 62, 253–261. [Google Scholar] [CrossRef]
- Kaur, S.; Chauhan, K.; Sachdeva, H. Protection against experimental visceral leishmaniasis by immunostimulation with herbal drugs derived from Withania somnifera and Asparagus racemosus. J. Med. Microbiol. 2014, 63, 1328–1338. [Google Scholar] [CrossRef]
- Tripathi, C.D.P.; Kushawaha, P.K.; Sangwan, R.S.; Mandal, C.; Misra-Bhattacharya, S.; Dube, A. Withania somnifera chemotype NMITLI 101R significantly increases the efficacy of antileishmanial drugs by generating strong IFN-γ and IL-12 mediated immune responses in Leishmania donovani infected hamsters. Phytomedicine 2017, 24, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, C.; Gupta, R.; Kushawaha, P.; Mandal, C.; Misra Bhattacharya, S.; Dube, A. Efficacy of Withania somnifera chemotypes NMITLI–101R, 118R and Withaferin A against experimental visceral leishmaniasis. Parasite Immunol. 2014, 36, 253–265. [Google Scholar] [CrossRef]
- Sachdeva, H.; Sehgal, R.; Kaur, S. Studies on the protective and immunomodulatory efficacy of Withania somnifera along with cisplatin against experimental visceral leishmaniasis. Parasitol. Res. 2013, 112, 2269–2280. [Google Scholar] [CrossRef]
- Chandrasekaran, S.; Veronica, J.; Sundar, S.; Maurya, R. Alcoholic fractions F5 and F6 from Withania somnifera leaves show a potent Antileishmanial and Immunomodulatory activities to control experimental Visceral Leishmaniasis. Front. Med. 2017, 4, 55. [Google Scholar] [CrossRef] [PubMed]
- Grover, A.; Katiyar, S.P.; Jeyakanthan, J.; Dubey, V.K.; Sundar, D. Blocking Protein kinase C signaling pathway: Mechanistic insights into the anti-leishmanial activity of prospective herbal drugs from Withania somnifera. BMC Genom. 2012, 13, S20. [Google Scholar] [CrossRef] [PubMed]
- Phadke, S.; Pathak, D.; Somani, D. Design and In silico Studies of 2, 5-Disubstituted 1, 2, 4-Triazole and 1, 3, 4-Thiadiazole Derivatives as Pteridine Reductase 1 Inhibitors. J. Pharm. Res. Int. 2021, 33, 166–178. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Mohammed, R.; Taher, M.A.; AbouZid, S.F.; Mansour, M.A.; Almahmoud, S.A.; Huwaimel, B.; Amin, E. Characterization of Promising Cytotoxic Metabolites from Tabebuia guayacan Hemsl.: Computational Prediction and In Vitro Testing. Plants 2022, 11, 888. [Google Scholar] [CrossRef] [PubMed]
- Arif, R.; Ahmad, S.; Mustafa, G.; Mahrosh, H.S.; Ali, M.; Tahir ul Qamar, M.; Dar, H.R. Molecular Docking and Simulation Studies of Antidiabetic Agents Devised from Hypoglycemic Polypeptide-P of Momordica charantia. Biomed. Res. Int. 2021, 2021, 5561129. [Google Scholar] [CrossRef]
- Mahnashi, M.H.; Alyami, B.A.; Alqahtani, Y.S.; Jan, M.S.; Rashid, U.; Sadiq, A.; Alqarni, A.O. Phytochemical profiling of bioactive compounds, anti-inflammatory and analgesic potentials of Habenaria digitata Lindl.: Molecular docking based synergistic effect of the identified compounds. J. Ethnopharmacol. 2021, 273, 113976. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, A.; Gupta, U. Molecular Docking studies on the Anti-fungal activity of Allium sativum (Garlic) against Mucormycosis (black fungus) by BIOVIA discovery studio visualizer 21.1.0.0. Ann. Antivir. Antiretrovir. 2021, 5, 28–32. [Google Scholar]
- Allam, A.E.; Abouelela, M.E.; Assaf, H.K.; Sayed, A.M.; Nafady, A.M.; El-Shanawany, M.A.; Takano, F.; Ohta, T. Phytochemical and in silico studies for potential constituents from Centaurium spicatum as candidates against the SARS-CoV-2 main protease and RNA-dependent RNA polymerase. Nat. Prod. Res. 2021, 36, 5724–5731. [Google Scholar] [CrossRef]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Majumdar, D.N. Withania somnifera. II. Alkaloidal constituents and their chemical characterization. Indian J. Pharm. 1955, 17, 158–161. [Google Scholar]
- Tetali, S.D.; Acharya, S.; Ankari, A.B.; Nanakram, V.; Raghavendra, A.S. Metabolomics of Withania somnifera (L.) Dunal: Advances and applications. J. Ethnopharmacol. 2021, 267, 113469. [Google Scholar] [CrossRef]
- Remya, C.; Dileep, K.; Variayr, E.; Sadasivan, C. An in silico guided identification of nAChR agonists from Withania somnifera. Front. Life Sci. 2016, 9, 201–213. [Google Scholar] [CrossRef]
- Kannan, N.; Kulandaivelu, G. Novel method to isolate Withaferin A from Withania somnifera roots and its bioactivity. Allelopath. J. 2007, 20, 213. [Google Scholar]
- Khanna, K.; Schwarting, A.; Bobbitt, J. The occurrence of isopelletierine in Withania somnifera. J. Pharm. Sci. 1962, 51, 1194. [Google Scholar] [CrossRef] [PubMed]
- Sivanandhan, G.; Mariashibu, T.S.; Arun, M.; Rajesh, M.; Kasthurirengan, S.; Selvaraj, N.; Ganapathi, A. The effect of polyamines on the efficiency of multiplication and rooting of Withania somnifera (L.) Dunal and content of some withanolides in obtained plants. Acta Physiol. Plant. 2011, 33, 2279–2288. [Google Scholar] [CrossRef]
- Schröter, H.-B.; Neumann, D.; Katritzky, A.; Swinbourne, F. Withasomnine. A pyrazole alkaloid from Withania somnifera Dunl. Tetrahedron 1966, 22, 2895–2897. [Google Scholar] [CrossRef]
- Kandil, F.; El Sayed, N.; Abou-Douh, A.; Ishak, M.; Mabry, T.J. Flavonol glycosides and phenolics from Withania somnifera. Phytochemistry 1994, 37, 1215–1216. [Google Scholar] [CrossRef]
- Bolleddula, J.; Fitch, W.; Vareed, S.K.; Nair, M.G. Identification of metabolites in Withania sominfera fruits by liquid chromatography and high-resolution mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 1277–1290. [Google Scholar] [CrossRef]
- Lee, S.R.; Lee, B.S.; Yu, J.S.; Kang, H.; Yoo, M.J.; Yi, S.A.; Han, J.-W.; Kim, S.; Kim, J.K.; Kim, J.-C. Identification of anti-adipogenic withanolides from the roots of Indian ginseng (Withania somnifera). J. Ginseng Res. 2022, 46, 357–366. [Google Scholar] [CrossRef]
- Rajalakshmy, M.; Geetha, G. Isolation and identification of withasomnine, withanolides and butein from industrial herbal marc of Withania somnifera (L.) Dunal. Indian J. Nat. Prod. Resour. 2016, 7, 116–124. [Google Scholar] [CrossRef]
- Senthil, K.; Thirugnanasambantham, P.; Oh, T.J.; Kim, S.H.; Choi, H.K. Free radical scavenging activity and comparative metabolic profiling of in vitro cultured and field grown Withania somnifera roots. PLoS ONE 2015, 10, e0123360. [Google Scholar] [CrossRef]
- Poojari, P.; Kiran, K.R.; Swathy, P.S.; Muthusamy, A. Withania somnifera (L.) Dunal: An Overview of Bioactive Molecules, Medicinal Properties and Enhancement of Bioactive Molecules Through Breeding Strategies. In In Vitro Plant Breeding Towards Novel Agronomic Traits; Springer: Singapore, 2019; pp. 1–25. [Google Scholar] [CrossRef]
- Rautela, I.; Sharma, M.D.; Sharma, N.; Kishor, K.; Singh, K.; Sharma, N. Comparative GC-MS analysis of leaf and root extract of medicinal plant Withania somnifera. World J. Pharm. Res. 2018, 7, 956–972. [Google Scholar]
- Abou-Douh, A.M. New withanolides and other constituents from the fruit of Withania somnifera. Arch. Pharm. 2002, 335, 267–276. [Google Scholar] [CrossRef]
- Marslin, G.; Selvakesavan, R.K.; Franklin, G.; Sarmento, B.; Dias, A.C. Antimicrobial activity of cream incorporated with silver nanoparticles biosynthesized from Withania somnifera. Int. J. Nanomed. 2015, 10, 5955. [Google Scholar]
- Lockley, W.; Roberts, D.; Rees, H.; Goodwin, T. 24-Methylcholesta-5, 24 (25)-dien-3β-ol: A new sterol from Withania somnifera. Tetrahedron Lett. 1974, 15, 3773–3776. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S. Alterations in metabolite profile and free radical scavenging activities of Withania somnifera leaf and root extracts under supplemental ultraviolet-B radiation. Acta Physiol. Plant. 2015, 37, 260. [Google Scholar] [CrossRef]
- Dar, N.J.; Hamid, A.; Ahmad, M. Pharmacologic overview of Withania somnifera, the Indian Ginseng. Cell Mol. Life Sci. 2015, 72, 4445–4460. [Google Scholar] [CrossRef] [PubMed]
- Nile, S.H.; Liang, Y.; Wang, Z.; Zheng, J.; Sun, C.; Nile, A.; Patel, G.; Kai, G. Chemical composition, cytotoxic and pro-inflammatory enzyme inhibitory properties of Withania somnifera (L.) Dunal root extracts. S. Afr. J. Bot. 2021, 151, 46–53. [Google Scholar] [CrossRef]
- Girme, A.; Saste, G.; Pawar, S.; Balasubramaniam, A.K.; Musande, K.; Darji, B.; Satti, N.K.; Verma, M.K.; Anand, R.; Singh, R. Investigating 11 withanosides and withanolides by UHPLC–PDA and mass fragmentation studies from Ashwagandha (Withania somnifera). ACS Omega 2020, 5, 27933–27943. [Google Scholar] [CrossRef]
- Chen, L.-X.; He, H.; Qiu, F. Natural withanolides: An overview. Nat. Prod. Rep. 2011, 28, 705–740. [Google Scholar] [CrossRef]
- Dar, N.J.; Bhat, J.A.; Satti, N.K.; Sharma, P.R.; Hamid, A.; Ahmad, M. Withanone, an active constituent from Withania somnifera, affords protection against NMDA-induced excitotoxicity in neuron-like cells. Mol. Neurobiol. 2017, 54, 5061–5073. [Google Scholar] [CrossRef] [PubMed]
- Nittala, S.S.; Lavie, D. Chemistry and genetics of withanolides in Withania somnifera hybrids. Phytochemistry 1981, 20, 2741–2748. [Google Scholar] [CrossRef]
- Bessalle, R.; Lavie, D. Withanolide C, a chlorinated withanolide from Withania somnifera. Phytochemistry 1992, 31, 3648–3651. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Hussain, S.; Yousuf, S.; Dar, A. Chlorinated and diepoxy withanolides from Withania somnifera and their cytotoxic effects against human lung cancer cell line. Phytochemistry 2010, 71, 2205–2209. [Google Scholar] [CrossRef]
- Nittala, S.S.; Velde, V.V.; Frolow, F.; Lavie, D. Chlorinated withanolides from Withania somnifera and Acnistus breviflorus. Phytochemistry 1981, 20, 2547–2552. [Google Scholar] [CrossRef]
- Tong, X.; Zhang, H.; Timmermann, B.N. Chlorinated withanolides from Withania somnifera. Phytochem. Lett. 2011, 4, 411–414. [Google Scholar] [CrossRef]
- Trivedi, M.K.; Panda, P.; Sethi, K.K.; Jana, S. Metabolite profiling in Withania somnifera roots hydroalcoholic extract using LC/MS, GC/MS and NMR spectroscopy. Chem. Biodivers. 2017, 14, e1600280. [Google Scholar] [CrossRef] [PubMed]
- Misra, L.; Lal, P.; Sangwan, R.S.; Sangwan, N.S.; Uniyal, G.C.; Tuli, R. Unusually sulfated and oxygenated steroids from Withania somnifera. Phytochemistry 2005, 66, 2702–2707. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-M.; Marron, M.T.; Seddon, E.; McLaughlin, S.P.; Ray, D.T.; Whitesell, L.; Gunatilaka, A.L. 2, 3-Dihydrowithaferin A-3β-O-sulfate, a new potential prodrug of withaferin A from aeroponically grown Withania somnifera. Bioorg. Med. Chem. 2009, 17, 2210–2214. [Google Scholar] [CrossRef]
- Subbaraju, G.V.; Vanisree, M.; Rao, C.V.; Sivaramakrishna, C.; Sridhar, P.; Jayaprakasam, B.; Nair, M.G. Ashwagandhanolide, a bioactive dimeric thiowithanolide isolated from the roots of Withania somnifera. J. Nat. Prod. 2006, 69, 1790–1792. [Google Scholar] [CrossRef]
- Jayaprakasam, B.; Strasburg, G.A.; Nair, M.G. Potent lipid peroxidation inhibitors from Withania somnifera fruits. Tetrahedron 2004, 60, 3109–3121. [Google Scholar] [CrossRef]
- Ali, M.; Shuaib, M.; Ansari, S.H. Withanolides from the stem bark of Withania somnifera. Phytochemistry 1997, 44, 1163–1168. [Google Scholar] [CrossRef]
- Khan, M.I.; Maqsood, M.; Saeed, R.A.; Alam, A.; Sahar, A.; Kieliszek, M.; Miecznikowski, A.; Muzammil, H.S.; Aadil, R.M. Phytochemistry, Food Application, and Therapeutic Potential of the Medicinal Plant (Withania coagulans): A Review. Molecules 2021, 26, 6881. [Google Scholar] [CrossRef] [PubMed]
- Vitali, G.; Conte, L.; Nicoletti, M. Withanolide composition and in vitro culture of Italian Withania somnifera. Planta Med. 1996, 62, 287–288. [Google Scholar] [CrossRef] [PubMed]
- Glotter, E.; Kirson, I.; Abraham, A.; Lavie, D. Constituents of Withania somnifera Dun—XIII: The withanolides of chemotype III. Tetrahedron 1973, 29, 1353–1364. [Google Scholar] [CrossRef]
- Xu, Y.-M.; Gao, S.; Bunting, D.P.; Gunatilaka, A.L. Unusual withanolides from aeroponically grown Withania somnifera. Phytochemistry 2011, 72, 518–522. [Google Scholar] [CrossRef]
- Abraham, A.; Kirson, I.; Lavie, D.; Glotte, E. The withanolides of Withania somnifera chemotypes I and II. Phytochemistry 1975, 14, 189–194. [Google Scholar] [CrossRef]
- Gottlied, H.E.; Kirson, I. 13C NMR spectroscopy of the withanolides and other highly oxygenated C28 steroids. Org. Magn. Reson. 1981, 16, 20–25. [Google Scholar] [CrossRef]
- Jayaprakasam, B.; Nair, M.G. Cyclooxygenase-2 enzyme inhibitory withanolides from Withania somnifera leaves. Tetrahedron 2003, 59, 841–849. [Google Scholar] [CrossRef]
- Zhao, J.; Nakamura, N.; Hattori, M.; Kuboyama, T.; Tohda, C.; Komatsu, K. Withanolide derivatives from the roots of Withania somnifera and their neurite outgrowth activities. Chem. Pharm. Bull. 2002, 50, 760–765. [Google Scholar] [CrossRef]
- Joshi, P.; Misra, L.; Siddique, A.A.; Srivastava, M.; Kumar, S.; Darokar, M.P. Epoxide group relationship with cytotoxicity in withanolide derivatives from Withania somnifera. Steroids 2014, 79, 19–27. [Google Scholar] [CrossRef] [PubMed]
- El-Olemy, M.; Kadry, H. Withanolides of Withania somnifera L. Dunal growing wild in Egypt. Bull. Pharm. Sci. 1984, 7, 430–443. [Google Scholar] [CrossRef]
- Anjaneyulyu, A.; Rao, D.S. A new withanolide from the leaves of Withania somnifera. Indian J. Chem. Sec. B 1997, 36, 161–165. [Google Scholar]
- Arshad Jamal, S.; Iqbal Choudhary, M. Two new withanolides from Withania somnifera. Heterocycles 1992, 34, 689–698. [Google Scholar]
- Kundu, A.B.; Mukherjee, A.; Dey, A.K. A new withanolide from the seeds of Withania somnifera Dunal. Indian J. Chem. Sect. B 1976, 14, 434–435. [Google Scholar]
- Kumar, A.; Ali, M.; Mir, S. A new withanolide from the roots of Withania somnifera. Indian J. Chem. Sect. B 2004, 43, 2001–2003. [Google Scholar] [CrossRef]
- Neogi, P.; Kawai, M.; Butsugan, Y.; Mori, Y.; Suzuki, M. Withacoagin, a new withanolide from Withania coagulans roots. Bull. Chem. Soc. Jpn. 1988, 61, 4479–4481. [Google Scholar] [CrossRef]
- Kirson, I.; Cohen, A.; Abraham, A. Withanolides Q and R, two new 23-hydroxy-steroidal lactones. J. Chem. Soc. Perkin Trans. 1 1975, 21, 2136–2138. [Google Scholar] [CrossRef]
- Arshad Jamal, S.; Qureshi, S.; Noor Ali, S.; Choudhary, M.I. Bioactivities and structural studies of withanolides from Withania somnifera. Chem. Heterocycl. Compd. 1995, 31, 1047–1059. [Google Scholar] [CrossRef]
- Jamal, S.A.; Choudhary, M.I.; Asif, E. Two withanolides from Withania somnifera. Phytochemistry 1991, 30, 3824–3826. [Google Scholar]
- Choudhary, M.I.; Yousuf, S.; Nawaz, S.A.; Ahmed, S. Cholinesterase inhibiting withanolides from Withania somnifera. Chem. Pharm. Bull. 2004, 52, 1358–1361. [Google Scholar] [CrossRef]
- Chaurasiya, N.D.; Uniyal, G.C.; Lal, P.; Misra, L.; Sangwan, N.S.; Tuli, R.; Sangwan, R.S. Analysis of withanolides in root and leaf of Withania somnifera by HPLC with photodiode array and evaporative light scattering detection. Phytochem. Anal. 2008, 19, 148–154. [Google Scholar] [CrossRef]
- Iguchi, T.; Kuroda, M.; Ishihara, M.; Sakagami, H.; Mimaki, Y. Steroidal constituents isolated from the seeds of Withania somnifera. Nat. Prod. Res. 2021, 35, 2205–2210. [Google Scholar] [CrossRef]
- Misra, L.; Mishra, P.; Pandey, A.; Sangwan, R.S.; Sangwan, N.S.; Tuli, R. Withanolides from Withania somnifera roots. Phytochemistry 2008, 69, 1000–1004. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Abbas, S.; Jamal, S.A. Withania somnifera—A source of exotic withanolides. Heterocycles 1996, 2, 555–563. [Google Scholar]
- Kirson, I.; Glotter, E.; Abraham, A.; Lavie, D. Constituents of Withania somnifera dun—XI: The structure of three new withanolides. Tetrahedron 1970, 26, 2209–2219. [Google Scholar] [CrossRef]
- Kirson, I.; Abraham, A.; Lavie, D. Chemical Analysis of Hybrids of Withania somnifera L. (Dun.). 1. Chemotypes III (Israel) by Indian I (Delhi). Isr. J. Chem. 1977, 16, 20–24. [Google Scholar] [CrossRef]
- Velde, V.V.; Lavie, D. New withanolides of biogenetic interest from Withania somnifera. Phytochemistry 1981, 20, 1359–1364. [Google Scholar] [CrossRef]
- Kirson, I.; Glotter, E.; Lavie, D.; Abraham, A. Constitutents of Withania somnifera Dun. Part XII. The withanolides of an indian chemotype. J. Chem. Soc. C 1971, 2032–2044. [Google Scholar] [CrossRef]
- Nile, S.H.; Nile, A.; Gansukh, E.; Baskar, V.; Kai, G. Subcritical water extraction of withanosides and withanolides from ashwagandha (Withania somnifera L.) and their biological activities. Food Chem. Toxicol. 2019, 132, 110659. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, F.W.; Kirson, I.; Lavie, D.; Abraham, A. New withanolides from a cross of a South African chemotype by chemotype II (Israel) in Withania somnifera. Phytochemistry 1980, 19, 1503–1507. [Google Scholar] [CrossRef]
- Duyu, T.; Khanal, P.; Dey, Y.N.; Jha, S.K. Network pharmacology of Withania somnifera against stress associated neurodegenerative diseases. Adv. Trad. Med. 2021, 21, 565–578. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Singh, P.; Sharma, S.; Singh, T.P.; Ethayathulla, A.; Kaur, P. Identification of bioactive molecule from Withania somnifera (Ashwagandha) as SARS-CoV-2 main protease inhibitor. J. Biomol. Struct. Dyn. 2021, 39, 5668–5681. [Google Scholar] [CrossRef]
- Moujir, L.M.; Llanos, G.G.; Araujo, L.; Amesty, A.; Bazzocchi, I.L.; Jiménez, I.A. Withanolide-type steroids from Withania aristata as potential anti-leukemic agents. Molecules 2020, 25, 5744. [Google Scholar] [CrossRef]
- Khan, M.A.; Khan, H.; Ali, T. Withanolides isolated from Withania somnifera with α-glucosidase inhibition. Med. Chem. Res. 2014, 23, 2386–2390. [Google Scholar] [CrossRef]
- Bessalle, R.; Lavie, D.; Frolow, F. Withanolide Y, a withanolide from a hybrid of Withania somnifera. Phytochemistry 1987, 26, 1797–1800. [Google Scholar] [CrossRef]
- Nittala, S.S.; Frolow, F.; Lavie, D. Novel occurrence of 14β-hydroxy-group on a withanolide skeleton; X-ray crystal and molecular structure of 14β-hydroxywithanone. J. Chem. Soc. Chem. Commun. 1981, 4, 178–179. [Google Scholar] [CrossRef]
- Zhang, H.; Timmermann, B.N. Withanolide structural revisions by 13C NMR spectroscopic analysis inclusive of the γ-gauche effect. J. Nat. Prod. 2016, 79, 732–742. [Google Scholar] [CrossRef]
- Kim, S.; Yu, J.S.; Lee, J.Y.; Choi, S.U.; Lee, J.; Kim, K.H. Cytotoxic withanolides from the roots of Indian ginseng (Withania somnifera). J. Nat. Prod. 2019, 82, 765–773. [Google Scholar] [CrossRef]
- Maurya, R. Withanolides: A prospective drug for infectious and tropical diseases. In Science of Ashwagandha: Preventive and Therapeutic Potentials; Springer: Berlin/Heidelberg, Germany, 2017; pp. 105–120. [Google Scholar]
- Kuroyanagi, M.; Shibata, K.; Umehara, K. Cell differentiation inducing steroids from Withania somnifera L. (Dun.). Chem. Pharm. Bull. 1999, 47, 1646–1649. [Google Scholar] [CrossRef]
- Glotter, E.; Waitman, R.; Lavie, D. Constituents of Withania somnifera Dun. Part VIII. A new steroidal lactone, 27-deoxy-14-hydroxy-withaferin A. J. Chem. Soc. C 1966, 1765–1767. [Google Scholar] [CrossRef]
- Jayaprakasam, B.; Zhang, Y.; Seeram, N.P.; Nair, M.G. Growth inhibition of human tumor cell lines by withanolides from Withania somnifera leaves. Life Sci. 2003, 74, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Lal, P.; Misra, L.; Sangwan, R.S.; Tuli, R. New withanolides from fresh berries of Withania somnifera. Z. Naturforsch. B 2006, 61, 1143–1147. [Google Scholar] [CrossRef]
- Abbas, S.; Jamal, S.A.; Choudhary, M.I. New withanolides from Withania sp. J. Nat. Prod. 1993, 56, 1000–1006. [Google Scholar]
- Matsuda, H.; Murakami, T.; Kishi, A.; Yoshikawa, M. Structures of withanosides I, II, III, IV, V, VI, and VII, new withanolide glycosides, from the roots of Indian Withania somnifera DUNAL. and inhibitory activity for tachyphylaxis to clonidine in isolated guinea-pig ileum. Bioorg. Med. Chem. 2001, 9, 1499–1507. [Google Scholar] [CrossRef]
- Ghosal, S.; Kaur, R.; Srivastava, R. Sitoindosides IX and X, new glycowithanolides from Withania somnifera. Ind. J. Nat. Prod. 1988, 4, 12–13. [Google Scholar]
- Bhattacharya, S.K.; Goel, R.K.; Kaur, R.; Ghosal, S. Anti-stress activity of sitoindosides VII and VIII, new acylsterylglucosides from Withania somnifera. Phytother. Res. 1987, 1, 32–37. [Google Scholar] [CrossRef]
- Abuzaid, A.A.; Abdoon, A.M.; Aldahan, M.A.; Alzahrani, A.G.; Alhakeem, R.F.; Asiri, A.M.; Alzahrani, M.H.; Memish, Z.A. Cutaneous leishmaniasis in Saudi Arabia: A comprehensive overview. Vector-Borne Zoonotic Dis. 2017, 17, 673–684. [Google Scholar] [CrossRef]
- Tate, E.W.; Bell, A.S.; Rackham, M.D.; Wright, M.H. N-Myristoyltransferase as a potential drug target in malaria and leishmaniasis. Parasitology 2014, 141, 37–49. [Google Scholar] [CrossRef]
- Brannigan, J.A.; Roberts, S.M.; Bell, A.S.; Hutton, J.A.; Hodgkinson, M.R.; Tate, E.W.; Leatherbarrow, R.J.; Smith, D.F.; Wilkinson, A.J. Diverse modes of binding in structures of Leishmania major N-myristoyltransferase with selective inhibitors. IUCrJ 2014, 1, 250–260. [Google Scholar] [CrossRef]
- Brannigan, J.A.; Smith, B.A.; Yu, Z.; Brzozowski, A.M.; Hodgkinson, M.R.; Maroof, A.; Price, H.P.; Meier, F.; Leatherbarrow, R.J.; Tate, E.W. N-myristoyltransferase from Leishmania donovani: Structural and functional characterisation of a potential drug target for visceral leishmaniasis. J. Mol. Biol. 2010, 396, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Abouelela, M.E.; Assaf, H.K.; Abdelhamid, R.A.; Elkhyat, E.S.; Sayed, A.M.; Oszako, T.; Belbahri, L.; El Zowalaty, A.E.; Abdelkader, M.S.A. Identification of potential SARS-CoV-2 main protease and spike protein inhibitors from the genus aloe: An in silico study for drug development. Molecules 2021, 26, 1767. [Google Scholar] [CrossRef] [PubMed]
















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).