
Circulating microRNAs Showed Specific Responses according to Metabolic Syndrome Components and Sex of Adults from a Population-Based Study

 ,
,  , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Sampling Design
2.2. Clinical and Metabolic Measurements
2.3. Plasma MicroRNA Levels Measurement
2.3.1. RNA Extraction
2.3.2. Reverse Transcription
2.3.3. Preamplification
2.3.4. QPCR
2.4. Target Gene Prediction
2.5. Statistical Analyses
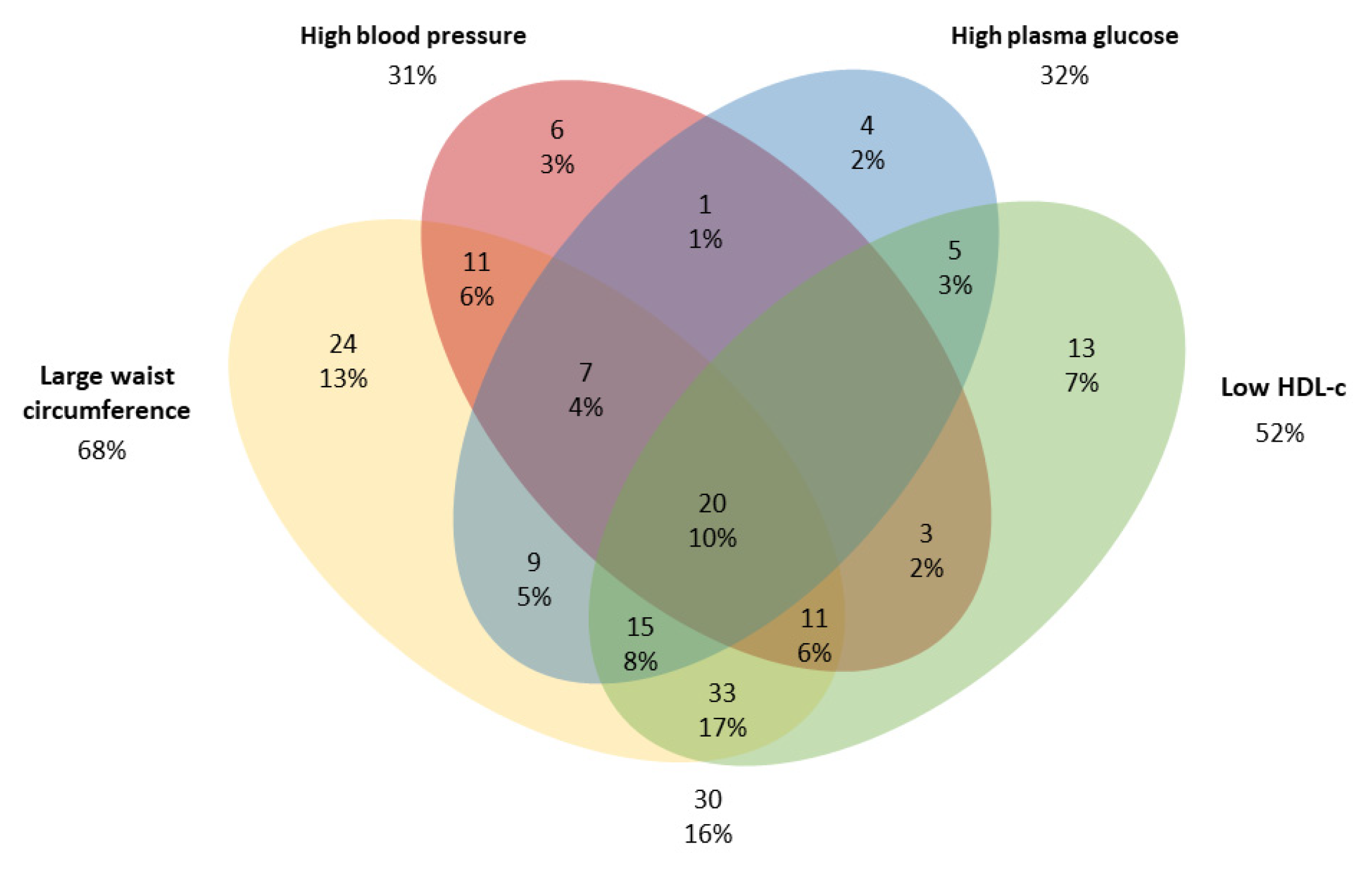
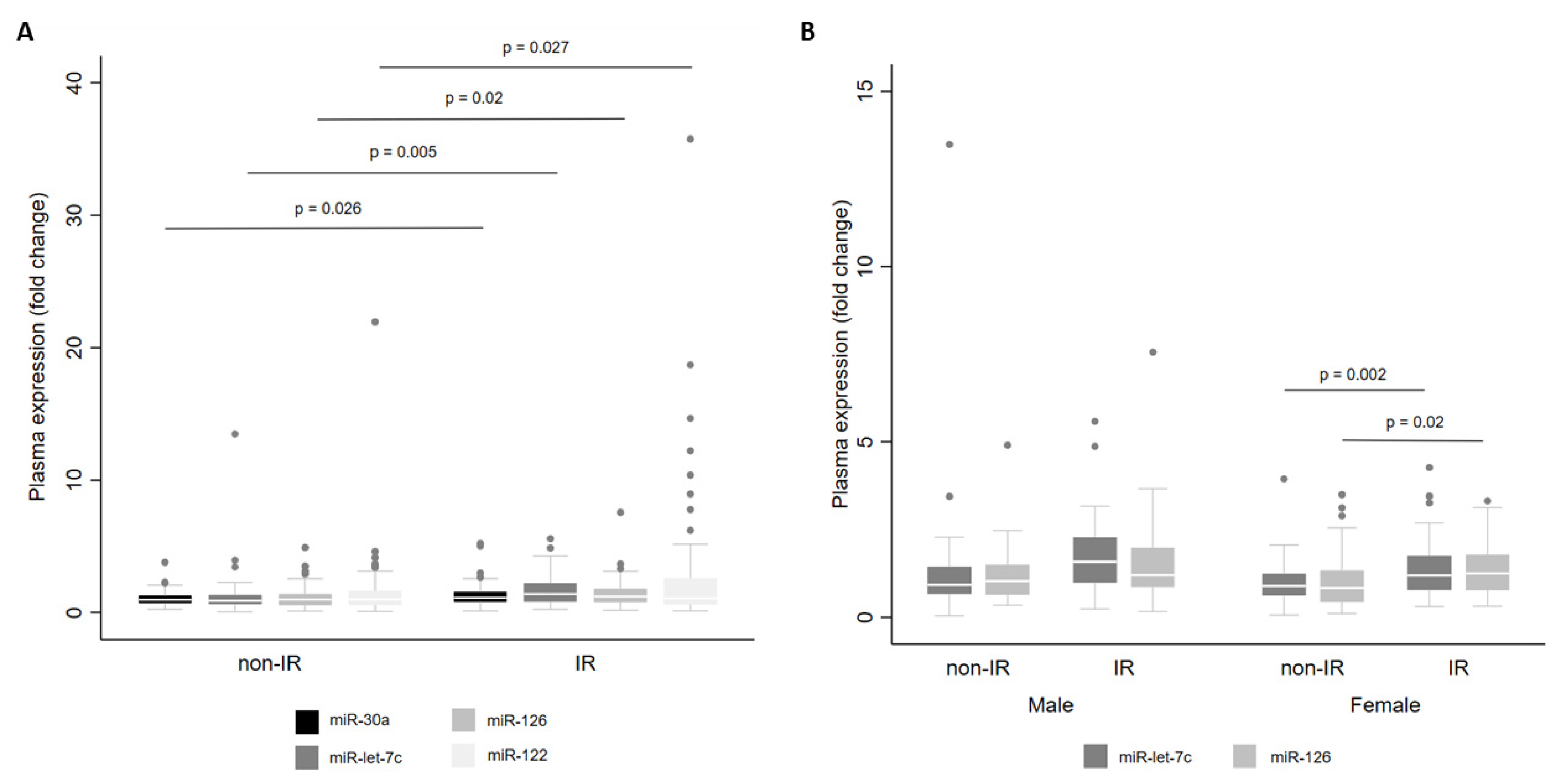
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Diabetes Federation. IDF Consensus Worldwide Definition of the Metabolic Syndrome. 2006. Available online: https://www.idf.org/e-library/consensus-statements/60-idfconsensus-worldwide-definitionof-the-metabolic-syndrome.html (accessed on 8 November 2022).
- Wang, Y.-T.; Tsai, P.-C.; Liao, Y.-C.; Hsu, C.-Y.; Juo, S.-H.H. Circulating MicroRNAs Have a Sex-Specific Association with Metabolic Syndrome. J. Biomed. Sci. 2013, 20, 72. [Google Scholar] [CrossRef] [PubMed]
- Quintanilha, B.J.; Reis, B.Z.; Duarte, G.B.S.; Cozzolino, S.M.F.; Rogero, M.M. Nutrimiromics: Role of MicroRNAs and Nutrition in Modulating Inflammation and Chronic Diseases. Nutrients 2017, 9, 1168. [Google Scholar] [CrossRef] [PubMed]
- Breininger, S.P.; Sabater, L.; Malcomson, F.C.; Afshar, S.; Mann, J.; Mathers, J.C. Obesity and Roux-En-Y Gastric Bypass Drive Changes in MiR-31 and MiR-215 Levels in the Human Rectal Mucosa. Int. J. Obes. 2022, 46, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Landrier, J.F.; Derghal, A.; Mounien, L. MicroRNAs in Obesity and Related Metabolic Disorders. Cells 2019, 8, 859. [Google Scholar] [CrossRef]
- Koh, E.H.; Chernis, N.; Saha, P.K.; Xiao, L.; Bader, D.A.; Zhu, B.; Rajapakshe, K.; Hamilton, M.P.; Liu, X.; Perera, D.; et al. MiR-30a Remodels Subcutaneous Adipose Tissue Inflammation to Improve Insulin Sensitivity in Obesity. Diabetes 2018, 67, 2541–2553. [Google Scholar] [CrossRef]
- Brandão-Lima, P.N.; de Carvalho, G.B.; Payolla, T.B.; Sarti, F.M.; Rogero, M.M. Circulating microRNA Related to Cardiometabolic Risk Factors for Metabolic Syndrome: A Systematic Review. Metabolites 2022, 12, 1044. [Google Scholar] [CrossRef]
- Qin, B.; Xiao, B.; Liang, D.; Li, Y.; Jiang, T.; Yang, H. MicroRNA Let-7c Inhibits Bcl-Xl Levels and Regulates Ox-LDL-Induced Endothelial Apoptosis. BMB Rep. 2012, 45, 464–469. [Google Scholar] [CrossRef]
- Giardina, S.; Hernandez-Alonso, P.; Díaz-Lopez, A.; Salas-Huetos, A.; Salas-Salvadó, J.; Bulló, M. Changes in Circulating MiRNAs in Healthy Overweight and Obese Subjects: Effect of Diet Composition and Weight Loss. Clin. Nutr. 2019, 38, 438–443. [Google Scholar] [CrossRef]
- Luo, Y.F.; Wan, X.X.; Zhao, L.L.; Guo, Z.; Shen, R.T.; Zeng, P.Y.; Wang, L.H.; Yuan, J.J.; Yang, W.J.; Yue, C.; et al. MicroRNA-139-5p Upregulation Is Associated with Diabetic Endothelial Cell Dysfunction by Targeting c-Jun. Aging 2021, 13, 1186–1211. [Google Scholar] [CrossRef]
- O’Neil, A.; Scovelle, A.J.; Milner, A.J.; Kavanagh, A. Gender/Sex as a Social Determinant of Cardiovascular Risk. Circulation 2018, 137, 854–864. [Google Scholar] [CrossRef]
- Fisberg, R.M.; Sales, C.H.; De Mello Fontanelli, M.; Pereira, J.L.; Alves, M.C.G.P.; Escuder, M.M.L.; César, C.L.G.; Goldbaum, M. 2015 Health Survey of São Paulo with Focus in Nutrition: Rationale, Design, and Procedures. Nutrients 2018, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.C.G.P.; Escuder, M.M.L.; Goldbaum, M.; Barros, M.B.d.A.; Fisberg, R.M.; Cesar, C.L.G. Sampling Plan in Health Surveys, City of São Paulo, Brazil, 2015. Rev. Saúde Pública 2018, 52, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis Model Assessment: Insulin Resistance and β-Cell Function from Fasting Plasma Glucose and Insulin Concentrations in Man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Geloneze, B.; Vasques, A.C.J.; Stabe, C.F.C.; Pareja, J.C.; Rosado, L.E.F.P.d.L.; Queiroz, E.C.d.; Tambascia, M.A. HOMA1-IR and HOMA2-IR Indexes in Identifying Insulin Resistance and Metabolic Syndrome: Brazilian Metabolic Syndrome Study (BRAMS). Arq. Bras. Endocrinol. Metabol. 2009, 53, 281–287. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting Effective MicroRNA Target Sites in Mammalian MRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The Biochemical Basis of MicroRNA Targeting Efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. MiRDB: An Online Database for Prediction of Functional MicroRNA Targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef]
- Streiner, D.L.; Norman, G.R. Correction for multiple testing: Is there a resolution? Chest 2011, 140, 16–18. [Google Scholar] [CrossRef]
- Lee, E.Y.; Han, K.; Kim, D.H.; Park, Y.M.; Kwon, H.S.; Yoon, K.H.; Kim, M.K.; Lee, S.H. Exposure-Weighted Scoring for Metabolic Syndrome and the Risk of Myocardial Infarction and Stroke: A Nationwide Population-Based Study. Cardiovasc. Diabetol. 2020, 19, 153. [Google Scholar] [CrossRef]
- Guembe, M.J.; Fernandez-Lazaro, C.I.; Sayon-Orea, C.; Toledo, E.; Moreno-Iribas, C.; Cosials, J.B.; Reyero, J.B.; Martínez, J.D.; Diego, P.G.; Uche, A.M.G.; et al. Risk for Cardiovascular Disease Associated with Metabolic Syndrome and Its Components: A 13-Year Prospective Study in the RIVANA Cohort. Cardiovasc. Diabetol. 2020, 19, 195. [Google Scholar] [CrossRef]
- Calderon-Dominguez, M.; Mangas, A.; Belmonte, T.; Quezada-Feijoo, M.; Ramos, M.; Toro, R. Ischemic Dilated Cardiomyopathy Pathophysiology through MicroRNA-16-5p. Rev. Española Cardiol. 2021, 74, 740–749. [Google Scholar] [CrossRef]
- Toro, R.; Pérez-Serra, A.; Mangas, A.; Campuzano, O.; Sarquella-Brugada, G.; Quezada-Feijoo, M.; Ramos, M.; Alcalá, M.; Carrera, E.; García-Padilla, C.; et al. MiR-16-5p Suppression Protects Human Cardiomyocytes against Endoplasmic Reticulum and Oxidative Stress-Induced Injury. Int. J. Mol. Sci. 2022, 23, 1036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Zhang, Y.N.; Teng, Z.Y. Downregulation of MiR-16 Protects H9c2(2-1) Cells against Hypoxia/Reoxygenation Damage by Targeting CIAPIN1 and Regulating the NF-ΚB Pathway. Mol. Med. Rep. 2019, 20, 3113–3122. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Brown, J.L.; Rosa, M.E.; Brown, L.A.; Perry, R.A.; Wiggs, M.P.; Nilsson, M.I.; Crouse, S.F.; Fluckey, J.D.; Washington, T.A.; et al. MicroRNA-16 Is Downregulated During Insulin Resistance and Controls Skeletal Muscle Protein Accretion. J. Cell. Biochem. 2016, 117, 1775–1787. [Google Scholar] [CrossRef]
- Meng, X.; Ji, Y.; Wan, Z.; Zhao, B.; Feng, C.; Zhao, J.; Li, H.; Song, Y. Inhibition of MiR-363 Protects Cardiomyocytes against Hypoxia-Induced Apoptosis through Regulation of Notch Signaling. Biomed. Pharmacother. 2017, 90, 509–516. [Google Scholar] [CrossRef]
- Peng, Y.H.; Wang, P.; He, X.Q.; Hong, M.Z.; Liu, F. Micro Ribonucleic Acid-363 Regulates the Phosphatidylinositol 3-Kinase/Threonine Protein Kinase Axis by Targeting NOTCH1 and Forkhead Box C2, Leading to Hepatic Glucose and Lipids Metabolism Disorder in Type 2 Diabetes Mellitus. J. Diabetes Investig. 2022, 13, 236–248. [Google Scholar] [CrossRef]
- Copps, K.D.; White, M.F. White Regulation of Insulin Sensitivity by Serine/Threonine Phosphorylation of Insulin Receptor Substrate Proteins IRS1 and IRS2. Diabetologia 2012, 55, 2565–2582. [Google Scholar] [CrossRef]
- Saha, P.K.; Hamilton, M.P.; Rajapakshe, K.; Putluri, V.; Felix, J.B.; Masschelin, P.; Cox, A.R.; Bajaj, M.; Putluri, N.; Coarfa, C.; et al. MiR-30a Targets Gene Networks That Promote Browning of Human and Mouse Adipocytes. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E667–E677. [Google Scholar] [CrossRef]
- Swaminathan, S.; Suzuki, K.; Seddiki, N.; Kaplan, W.; Cowley, M.J.; Hood, C.L.; Clancy, J.L.; Murray, D.D.; Méndez, C.; Gelgor, L.; et al. Differential Regulation of the Let-7 Family of MicroRNAs in CD4 + T Cells Alters IL-10 Levels. J. Immunol. 2012, 188, 6238–6246. [Google Scholar] [CrossRef]
- Piccand, J.; Strasser, P.; Hodson, D.J.; Meunier, A.; Ye, T.; Keime, C.; Birling, M.C.; Rutter, G.A.; Gradwohl, G. Rfx6 Maintains the Functional Identity of Adult Pancreatic β Cells. Cell Rep. 2014, 9, 2219–2232. [Google Scholar] [CrossRef]
- Iacomino, G.; Siani, A. Role of MicroRNAs in Obesity and Obesity-Related Diseases. Genes Nutr. 2017, 12, 23. [Google Scholar] [CrossRef] [PubMed]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/Let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Carrasco-Wong, I.; Dominguez, A.; Arnaiz, P.; Farías, M.; Barja, S.; Mardones, F.; Casanello, P. Micro-RNAs Let7e and 126 in Plasma as Markers of Metabolic Dysfunction in 10 to 12 Years Old Children. PLoS ONE 2015, 10, e0128140. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, P.; Li, F.; Yuan, G.; Wang, X.; Zhang, A.; Li, F. Plasma MiR-22-5p, MiR-132-5p, and MiR-150-3p Are Associated with Acute Myocardial Infarction. Biomed Res. Int. 2019, 2019, 5012648. [Google Scholar] [CrossRef]
- Yaman, S.O.; Orem, A.; Yucesan, F.B.; Kural, B.V.; Orem, C. Evaluation of Circulating MiR-122, MiR-30c and MiR-33a Levels and Their Association with Lipids, Lipoproteins in Postprandial Lipemia. Life Sci. 2021, 264, 118585. [Google Scholar] [CrossRef]
- Ye, D.; Zhang, T.; Lou, G.; Xu, W.; Dong, F.; Chen, G.; Liu, Y. Plasma MiR-17, MiR-20a, MiR-20b and MiR-122 as Potential Biomarkers for Diagnosis of NAFLD in Type 2 Diabetes Mellitus Patients. Life Sci. 2018, 208, 201–207. [Google Scholar] [CrossRef]
- Zhang, H.G.; Zhang, Q.J.; Li, B.W.; Li, L.H.; Song, X.H.; Xiong, C.M.; Zou, Y.B.; Liu, B.Y.; Han, J.Q.; Xiu, R.J. The Circulating Level of MiR-122 Is a Potential Risk Factor for Endothelial Dysfunction in Young Patients with Essential Hypertension. Hypertens. Res. 2020, 43, 511–517. [Google Scholar] [CrossRef]
- Veitch, S.; Njock, M.-S.; Chandy, M.; Siraj, M.A.; Chi, L.; Mak, H.; Yu, K.; Rathnakumar, K.; Perez-Romero, C.A.; Chen, Z.; et al. MiR-30 Promotes Fatty Acid Beta-Oxidation and Endothelial Cell Dysfunction and Is a Circulating Biomarker of Coronary Microvascular Dysfunction in Pre-Clinical Models of Diabetes. Cardiovasc. Diabetol. 2022, 21, 31. [Google Scholar] [CrossRef]
- Li, J.; Su, L.; Gong, Y.Y.; Ding, M.L.; Hong, S.B.; Yu, S.; Xiao, H. Downregulation of MiR-139-5p Contributes to the Antiapoptotic Effect of Liraglutide on the Diabetic Rat Pancreas and INS-1 Cells by Targeting IRS1. PLoS ONE 2017, 12, e0173576. [Google Scholar] [CrossRef]
- Canfrán-duque, A.; Lin, C.; Goedeke, L.; Suárez, Y.; Fernández-hernando, C. MicroRNAs and HDL Metabolism. Arter. Thromb. Vasc. Biol. 2016, 36, 1076–1084. [Google Scholar] [CrossRef]
- Ma, E.; Fu, Y.; Garvey, W.T. Relationship of Circulating MiRNAs with Insulin Sensitivity and Associated Metabolic Risk Factors in Humans. Metab. Syndr. Relat. Disord. 2018, 16, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Yang, J.; Liu, G.Q.; Gao, M.J.; Hou, W.F.; Zhang, L.; Gao, H.W.; Liu, Y.; Chen, G.A.; Hong, T.P. Dynamic Levels of MicroRNAs during the Differentiation of Human Embryonic Stem Cells into Insulin-Producing Cells. Gene 2013, 518, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Reis, B.Z.; Duarte, G.B.S.; Vargas-Mendez, E.; Ferreira, L.R.P.; Barbosa, F.; Cercato, C.; Rogero, M.M.; Cozzolino, S.M.F. Brazil Nut Intake Increases Circulating MiR-454-3p and MiR-584-5p in Obese Women. Nutr. Res. 2019, 67, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhang, Q.; Ma, X.; Wang, J.; Liang, T. MiRNA and MRNA Levels Analysis Reveals Potential Sex-Biased MiRNA Levels. Sci. Rep. 2017, 7, 39812. [Google Scholar] [CrossRef]
- Sharma, S.; Eghbali, M. Influence of Sex Differences on MicroRNA Gene Regulation in Disease. Biol. Sex Differ. 2014, 5, 3. [Google Scholar] [CrossRef]
- Shen, J.J.; Wang, Y.F.; Yang, W. Sex-Interacting MRNA-and MiRNA-EQTLs and Their Implications in Gene Levels Regulation and Disease. Front. Genet. 2019, 10, 313. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | All (n = 192) | Male (n = 87) | Female (n = 105) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Number of Risk Factors for MetS | Number of Risk Factors for MetS | ||||||||
| 0 | 1–2 | ≥3 | p-Value | 0 | 1–2 | ≥3 | p-Value | ||
| (n = 19) | (n = 38) | (n = 30) | (n = 10) | (n = 58) | (n = 37) | ||||
| Age (years) | 40.2 (38.5, 42.0) | 31.8 (27.8, 35.8) a | 37.9 (34.7, 41.1) b | 45.3 (42.4, 48.2) c | <0.001 | 36.8 (31.5, 42.0) a | 39.6 (37.6, 41.5) a | 47.3 (43.8, 50.8) b | <0.001 |
| Weight (kg) | 73.5 (70.7, 76.3) | 64.8 (60.3, 69.2) a | 79.7 (74.8, 84.5) b | 86.3 (81.6, 91.0) c | <0.001 | 53.4 (48.9, 57.9) a | 68.2 (63.2, 73.2) b | 75.3 (70.3, 80.2) c | <0.001 |
| Height (m) | 1.67 (1.65, 1.69) | 1.74 (1.70, 1.79) | 1.76 (1.73, 1.78) | 1.73 (1.71, 1.75) | 0.196 | 1.63 (1.55, 1.70) | 1.59 (1.57, 1.61) | 1.58 (1.55, 1.60) | 0.443 |
| BMI (kg/m2) | 26.2 (25.4, 27.1) | 21.1 (20.1, 22.2)a | 25.6 (24.2, 26.9) b | 28.7 (27.2, 30.2) c | <0.001 | 20.0 (19.2, 20.9) a | 26.6 (25.2, 28.1) b | 29.9 (28.4, 31.3) c | <0.001 |
| Waist circumference (cm) | 91.5 (89.1, 93.9) | 77.2 (73.9, 80.5)a | 92.4 (88.1, 96.7) b | 102.8 (99.8, 105.9) c | <0.001 | 74.0 (71.3, 76.8) a | 89.8 (85.5, 94.1) b | 98.4 (94.5, 102.2) c | <0.001 |
| SBP (mmHg) | 122.7 (120.1, 125.3) | 118.9 (116.0, 121.9) a | 122.8 (118.6, 127.0) a | 138.9 (131.6, 146.3) b | <0.001 | 104.1 (98.0, 110.2) a | 114.0 (111.1, 116.9) b | 132.0 (125.3, 138.7) c | <0.001 |
| DBP (mmHg) | 75.6 (73.9, 77.3) | 68.9 (66.5, 71.3) a | 74.7 (71.3, 78.0) b | 84.8 (79.8, 89.8) c | <0.001 | 65.8 (61.2, 70.5) a | 72.8 (70.8, 74.8) b | 81.3 (76.5, 86.2) c | <0.001 |
| HDL-c (mg/dL) | 45.4 (43.2, 47.7) | 55.3 (50.2, 60.4) a | 43.5 (39.0, 48.0) b | 36.5 (31.3, 41.7) c | <0.001 | 59.3 (54.4, 64.1) a | 48.7 (44.9, 52.5) b | 38.9 (35.9, 41.9) c | <0.001 |
| Triacyclglycerol (mg/dL) | 119.1 (105.9, 132.3) | 65.6 (49.9, 81.3) a | 112.3 (92.9, 131.8) b | 217.4 (175.9, 258.9) c | <0.001 | 53.1 (41.9, 64.3) a | 92.2 (80.9, 103.5) b | 145.5 (125.8, 165.1) c | <0.001 |
| Plasma glucose (mg/dL) | 101.6 (96.6, 106.5) | 86.4 (83.4, 89.5) a | 93.0 (90.6, 95.3)b | 134.2 (109.9, 158.4) c | <0.001 | 87.1 (84.0, 90.2) a | 92.3 (89.3, 95.3) b | 115.8 (103.2, 128.5) c | <0.001 |
| Plasma insulin (µUI/mL) | 13.1 (11.4, 14.8) | 5.7 (3.9, 7.4) a | 11.9 (8.3, 15.5) b | 22.2 (15.7, 28.6) c | <0.001 | 5.5 (4.4, 6.7) a | 10.7 (9.0, 12.4) b | 18.5 (14.5, 22.5) c | <0.001 |
| HOMA-IR | 3.5 (2.9, 4.1) | 1.2 (0.8, 1.6) a | 2.7 (1.9, 3.6) b | 7.0 (4.8, 9.1) c | <0.001 | 1.2 (0.9, 1.4) a | 2.4 (2.0, 2.8) b | 5.8 (4.2, 7.4) c | <0.001 |
| Male (n = 87) | Female (n = 105) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fold Change | Total | Number of Risk Factors for MetS | Number of Risk Factors for MetS | ||||||
| (n = 192) | 0 | 1–2 | ≥3 | p-Value | 0 | 1–2 | ≥3 | p-Value | |
| miR-15a | 1.2 (1.1, 1.3) | 1.2 (0.9, 1.6) | 1.5 (1.2, 1.8) | 1.2 (1.0, 1.5) | 0.232 | 0.9 (0.6, 1.2) | 1.0 (0.9, 1.2) | 1.0 (0.8, 1.1) | 0.857 |
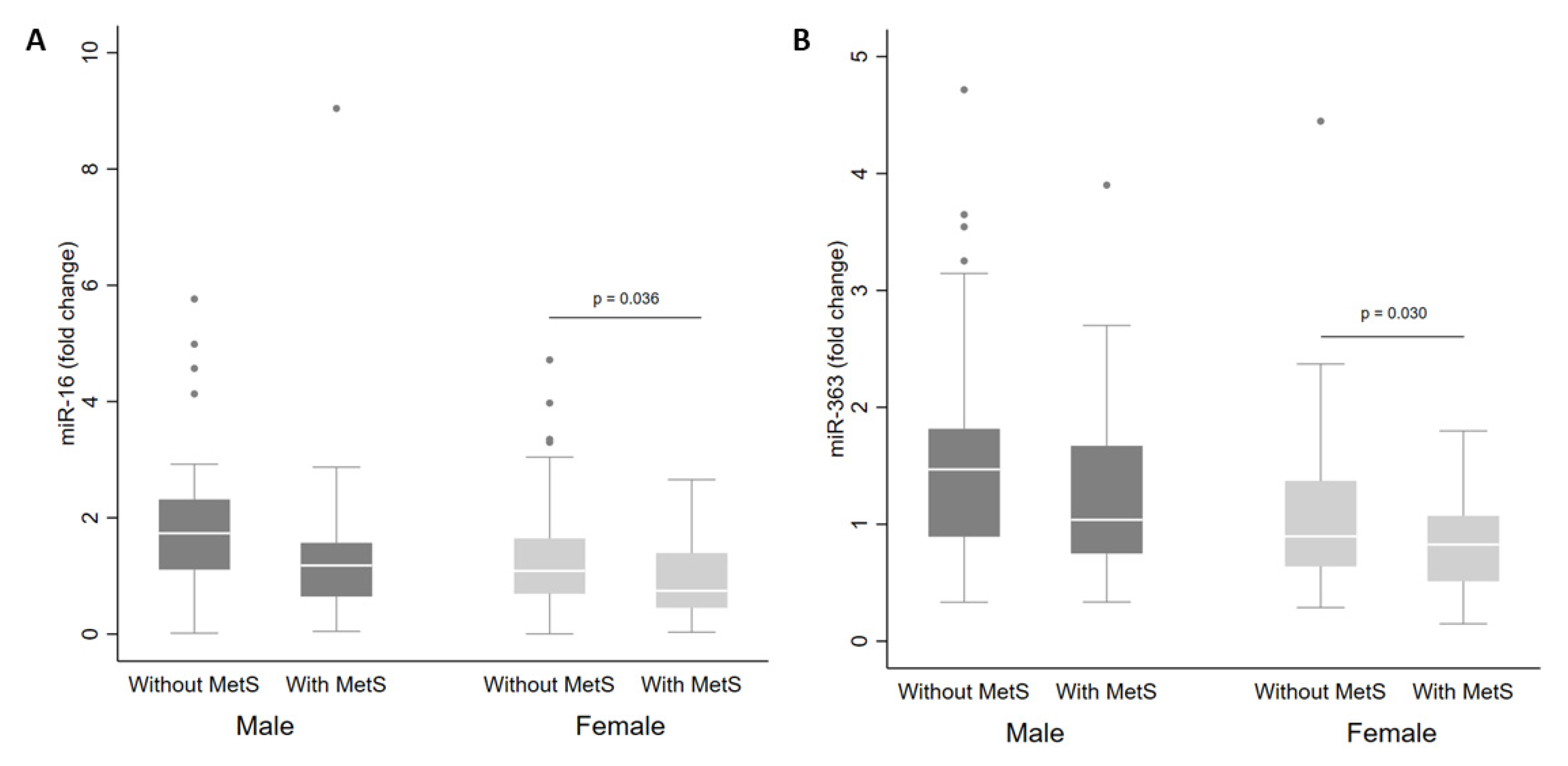
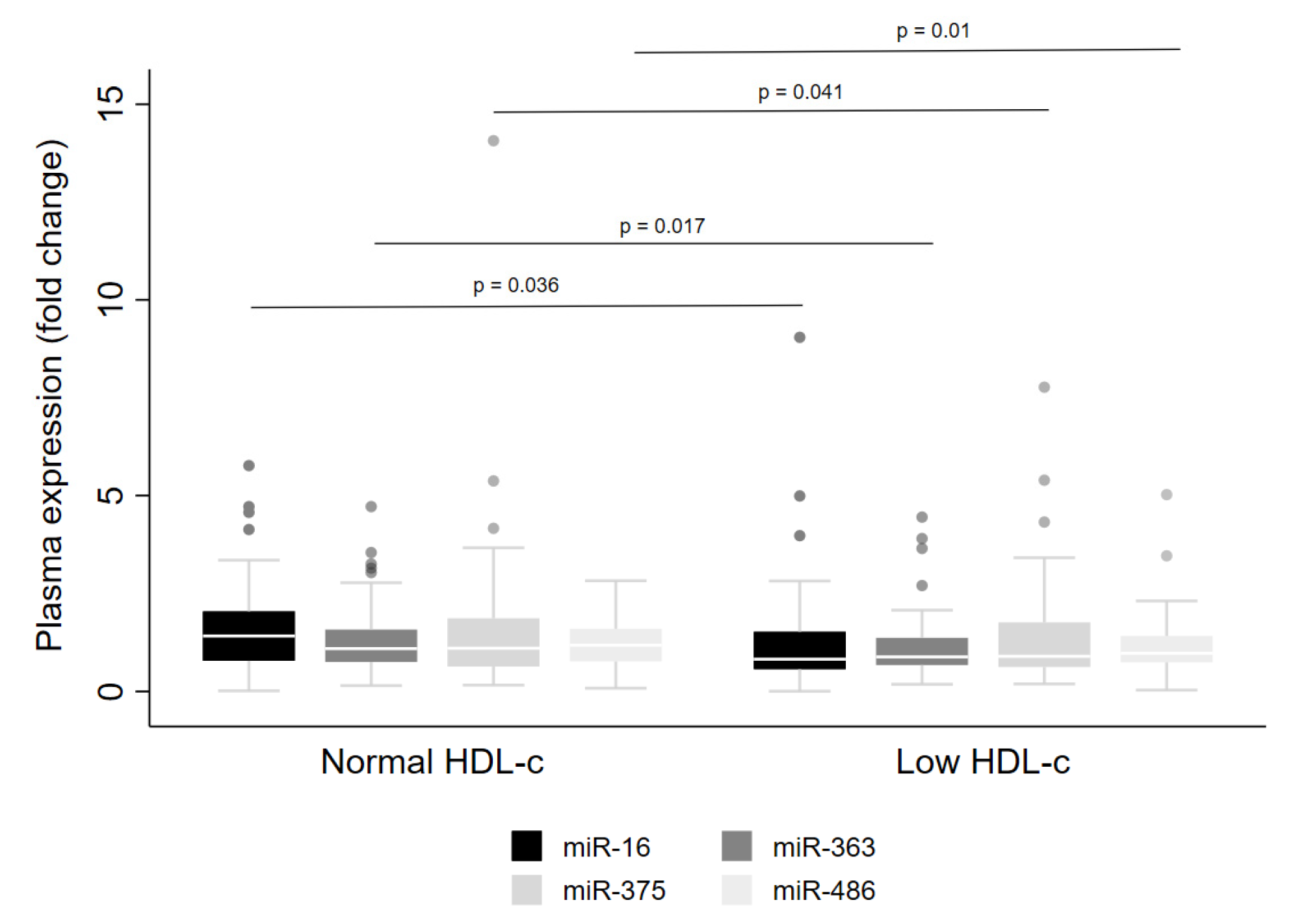
| miR-16 | 1.4 (1.2, 1.6) | 1.9 (1.3, 2.4) | 1.7 (1.3, 2.0) | 1.4 (0.7, 2.2) | 0.659 | 1.0 (0.7, 1.4) | 1.3 (1.1, 1.5) | 0.9 (0.7, 1.2) | 0.068 |
| miR-21 | 1.38 (1.19, 1.58) | 1.2 (0.6, 1.8) | 1.6 (1.1, 2.2) | 1.4 (0.9, 1.8) | 0.567 | 1.2 (0.8, 1.6) | 1.3 (1.0, 1.6) | 1.0 (0.8, 1.3) | 0.414 |
| miR-28-3p * | 1.61 (1.34, 1.88) | 1.5 (0.6, 2.3) | 1.7 (1.1, 2.4) | 1.6 (1.1, 2.2) | 0.884 | 1.1 (0.6, 1.6) | 1.6 (1.1, 2.2) | 1.3 (0.8, 1.8) | 0.397 |
| miR-30a-5p | 1.1 (1.0, 1.2) | 1.0 (0.8, 1.1)a | 1.3 (1.0, 1.5)b | 1.5 (1.1, 2.0)b | 0.029 | 1.0 (0.6, 1.5) | 0.9 (0.8, 1.0) | 1.0 (0.8, 1.2) | 0.633 |
| miR-30d | 1.3 (1.1, 1.5) | 1.16 (0.83, 1.50) | 1.5 (1.1, 2.0) | 1.6 (1.0, 2.1) | 0.528 | 0.9 (0.5, 1.2) | 1.2 (0.9, 1.5) | 1.1 (0.8, 1.4) | 0.316 |
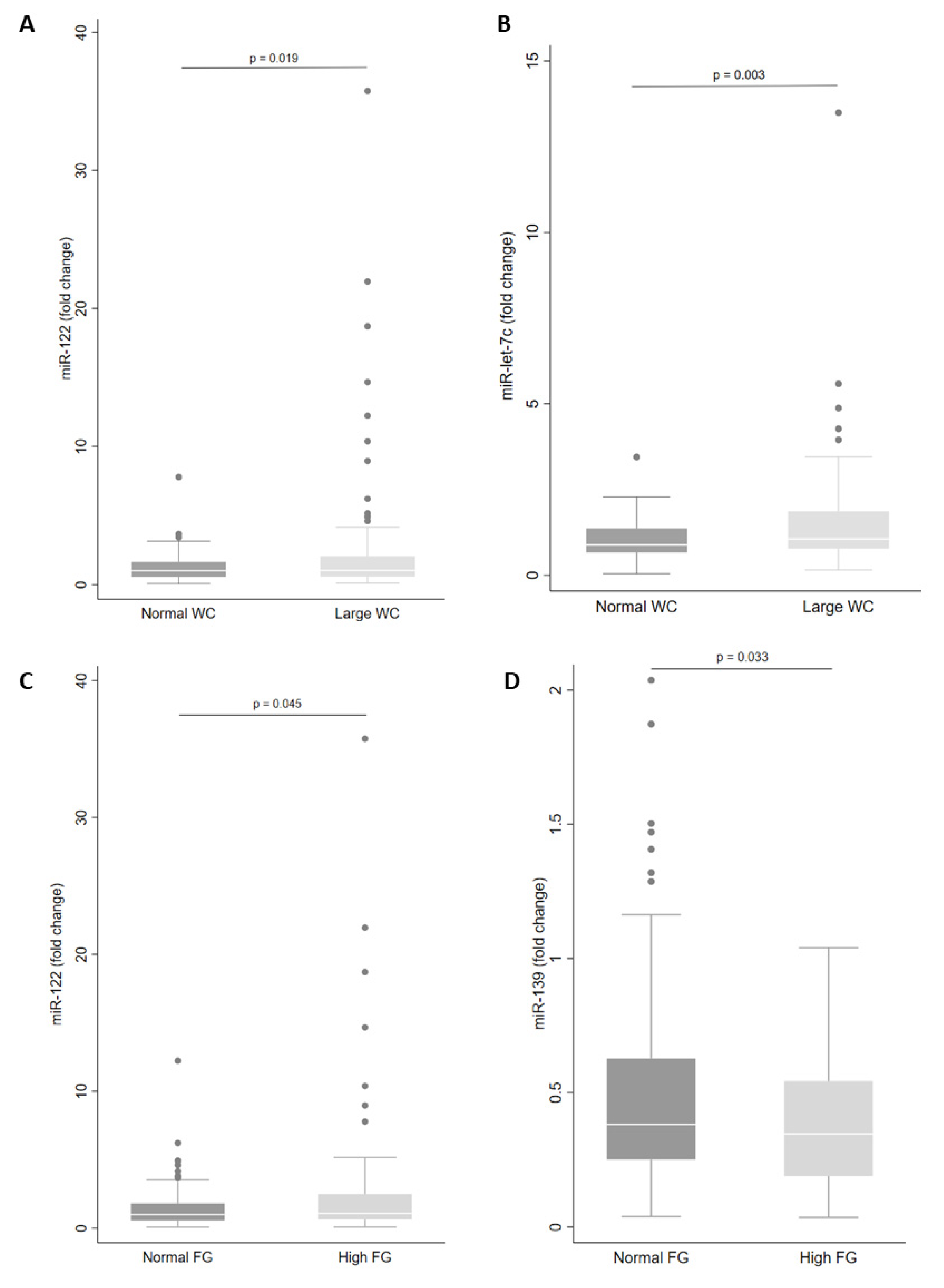
| miR-122 | 1.9 (1.3, 2.5) | 1.3 (0.8, 1.8) | 1.8 (1.2, 2.4) | 3.8 (1.5, 6.0) | 0.071 | 1.0 (0.6, 1.4) | 1.3 (0.6, 2.1) | 2.2 (0.1, 4.3) | 0.488 |
| miR-126 | 1.2 (1.1, 1.3) | 1.1 (0.6, 1.5) | 1.3 (1.0, 1.5) | 1.5 (0.9, 2.1) | 0.468 | 1.0 (0.8, 1.3) | 1.1 (0.9, 1.3) | 1.2 (0.9, 1.4) | 0.827 |
| miR-130b & | 1.3 (1.2, 1.5) | 1.6 (1.1, 2.0) | 1.4 (1.1, 1.8) | 1.3 (0.9, 1.6) | 0.566 | 1.1 (0.5, 1.7) | 1.3 (1.0, 1.6) | 1.0 (0.8, 1.3) | 0.292 |
| miR-139-3p # | 0.4 (0.4, 0.5) | 0.4 (0.3, 0.5) | 0.5 (0.3, 0.6) | 0.4 (0.3, 0.6) | 0.603 | 0.6 (0.1, 1.1) | 0.4 (0.3, 0.5) | 0.4 (0.2, 0.5) | 0.772 |
| miR-140-5p § | 1.1 (1.0, 1.3) | 1.3 (0.7, 1.8) | 1.2 (0.9, 1.5) | 1.1 (0.8, 1.5) | 0.925 | 0.6 (0.3, 0.9) | 1.1 (0.8, 1.4) | 1.1 (0.8, 1.4) | 0.05 |
| miR-146a | 1.5 (1.3, 1.8) | 1.3 (0.5, 2.0) | 1.9 (1.3, 2.5) | 1.6 (1.1, 2.2) | 0.384 | 1.0 (0.6, 1.5) | 1.5 (1.1, 1.8) | 1.3 (0.9, 1.8) | 0.334 |
| miR-150 | 1.2 (1.0, 1.4) | 1.5 (0.7, 2.3) | 1.2 (0.9, 1.5) | 1.3 (1.0, 1.7) | 0.793 | 0.8 (0.7, 0.9)a | 1.3 (0.9, 1.6)b | 1.0 (0.8, 1.2)a,b | 0.029 |
| miR-222 ** | 1.3 (1.2, 1.5) | 1.0 (0.5, 1.5) | 1.7 (1.1, 2.3) | 1.6 (0.9, 2.3) | 0.114 | 1.0 (0.7, 1.2) | 1.3 (1.0, 1.6) | 1.1 (0.8, 1.4) | 0.275 |
| miR-223 ** | 1.7 (1.4, 2.0) | 1.9 (0.8, 3.0) | 1.7 (1.46, 2.24) | 1.46 (1.03, 1.89) | 0.884 | 1.4 (0.5, 2.3) | 1.9 (1.3, 2.4) | 1.2 (0.88, 1.7) | 0.173 |
| miR-363 ** | 1.2 (1.1, 1.3) | 1.4 (1.0, 1.9) | 1.5 (1.2, 1.8) | 1.3 (1.0, 1.6) | 0.542 | 0.9 (0.6, 1.2) | 1.0 (0.8, 1.3) | 0.8 (0.6, 0.9) | 0.095 |
| miR-375 | 1.4 (1.1, 1.7) | 1.3 (1.0, 1.7) | 1.4 (1.0, 1.8) | 1.4 (0.8, 2.1) | 0.965 | 1.9 (0.9, 3.0) | 1.6 (0.9, 2.2) | 1.0 (0.7, 1.3) | 0.092 |
| miR-376a | 1.9 (1.5, 2.3) | 1.5 (0.4, 2.7) | 2.5 (1.5, 3.4) | 2.2 (0.7, 3.7) | 0.448 | 1.4 (0.4, 2.3) | 1.8 (1.3, 2.3) | 1.5 (0.9, 2.0) | 0.622 |
| miR-486-5p | 1.2 (1.1, 1.3) | 1.4 (1.0, 1.9) | 1.4 (1.2, 1.7) | 1.3 (0.9, 1.7) | 0.767 | 1.0 (0.7, 1.2) | 1.0 (0.9, 1.2) | 0.9 (0.7, 1.0) | 0.194 |
| miR-532-5p £ | 1.3 (1.1, 1.1) | 1.6 (0.8, 2.5) | 1.5 (1.1, 1.8) | 1.3 (1.0, 1.7) | 0.758 | 1.0 (0.6, 1.5) | 1.2 (0.9, 1.5) | 0.9 (0.7, 1.2) | 0.388 |
| miR-let-7c | 1.31 (1.13, 1.49) | 0.8 (0.5, 1.1)a | 1.6 (1.0, 2.1)b | 1.7 (1.3, 2.1)b | 0.003 | 1.0 (0.8, 1.2) | 1.0 (0.9, 1.2) | 1.2 (0.9, 1.5) | 0.423 |
| miRNA | Sex | Plasma Levels | Potential Target | Function | MetS Components | p-Value |
|---|---|---|---|---|---|---|
| miR-let-7c | Male | Up | CDK8 | Regulate transcription factors (SREBP, STAT1) and RNA polymerase II | Large WC | 0.015 |
| MAP family | Cell signaling (e.g., c-Jun) | |||||
| IGF2BP | Nutrient metabolism | |||||
| RFX6 | Regulate beta-cell maturation and function | |||||
| IL10 | ERK1/2, p38 and NF-κB signaling | |||||
| SOCS1/4 | Negative feedback on cytokine signaling | |||||
| CCL3 | Acute inflammatory state | |||||
| PRKAR2A | PKA activation | |||||
| miR-122 | Male | Up | PRKRA | Response to stress | Large WC | 0.015 |
| PDK4 | Regulate general metabolism | High blood pressure | 0.021 | |||
| miR-15a | Female | Up | SIRT4 AKT3 PDK4 VEGFA IKBKB CDK8 FOXO1 BCL2 | Mitochondrial functions Cell signaling Regulate general metabolism Cell proliferation and migration, apoptosis, permeabilization Activation of NF-kB Regulate transcription factors (SREBP, STAT1) and RNA polymerase II Regulate adipocytokines and insulin signaling Regulate cell death | Large WC | 0.041 |
| miR-16 | Male | Down | High triacylglycerol | 0.038 | ||
| Low HDL-c | 0.047 | |||||
| miR-222 | Female | Up | SOCS3/5 | Negative feedback of cytokine signaling | Large WC | <0.001 |
| PIK3R1 | Insulin metabolism | |||||
| CDK8 | Regulate transcription factors (SREBP, STAT1) and RNA polymerase II | |||||
| MAP3K2 | Cell signaling (NF-kB pathway) | |||||
| IGF1 | ERK signaling | |||||
| miR-146a | Female | Up | TRAF6 | Activated TLR4 signaling, Toll-Like receptor Signaling Pathways, NF-kB activation | Large WC | 0.046 |
| PRKAA2 | mTOR signaling, AMPK Signaling Pathway, Insulin signaling | |||||
| MARK1 | Energy metabolism | |||||
| miR-30d | Female | Up | SOCS1/3 | Negative feedback on cytokine signaling | Large WC | 0.047 |
| RFX6 | Regulate beta-cell maturation and function | |||||
| RUNX2 | Regulates osteogenesis and adipogenesis | |||||
| miR-30a | Male | Up | UCP3 | Protects mitochondria against lipid-induced oxidative stress | Large WC | 0.015 |
| PPARGC1A | Energy metabolism/blood pressure control, cellular cholesterol homeostasis | High blood pressure | 0.001 | |||
| SOCS3 | Negative feedback on cytokine signaling | |||||
| MAP3K | Cell signaling (e.g., c-Jun) | |||||
| IRS1/2 | Insulin signaling pathway | |||||
| PRKAR1A | PKA activation | |||||
| miR-139 | Female | Down | GSK3A | Regulates glycogen synthase, PI3K signaling pathways, transcription factors (e.g., c-Jun) | High FG | 0.016 |
| AKT1S1 | Glucose metabolism, mTOR and MAPK signaling | |||||
| PIK3R4 | Insulin and ERK/MAPK signaling | |||||
| IL10 | Inflammation | |||||
| miR-363 | Male | Down | MARK1 | Glucose metabolism | Low HDL-c | 0.026 |
| TRAF3 | Toll-like receptor (TLR3 and TLR4) cascade, TNF signaling | |||||
| MAPK8 | TLR4, TNF and IL-2 pathways | |||||
| NOTCH1 | Notch signaling pathway | |||||
| miR-486 | Male | Down | MAP3K7 | Cell signaling (e.g., c-Jun) | Reduced HDL-c | 0.012 |
| PIK3R1 | Insulin metabolism | |||||
| MARK1 | Glucose metabolism | |||||
| IGF1 | ERK signaling |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandão-Lima, P.N.; de Carvalho, G.B.; Payolla, T.B.; Sarti, F.M.; Fisberg, R.M.; Malcomson, F.C.; Mathers, J.C.; Rogero, M.M. Circulating microRNAs Showed Specific Responses according to Metabolic Syndrome Components and Sex of Adults from a Population-Based Study. Metabolites 2023, 13, 2. https://doi.org/10.3390/metabo13010002
Brandão-Lima PN, de Carvalho GB, Payolla TB, Sarti FM, Fisberg RM, Malcomson FC, Mathers JC, Rogero MM. Circulating microRNAs Showed Specific Responses according to Metabolic Syndrome Components and Sex of Adults from a Population-Based Study. Metabolites. 2023; 13(1):2. https://doi.org/10.3390/metabo13010002
Chicago/Turabian StyleBrandão-Lima, Paula N., Gabrielli B. de Carvalho, Tanyara B. Payolla, Flávia M. Sarti, Regina M. Fisberg, Fiona C. Malcomson, John C. Mathers, and Marcelo M. Rogero. 2023. "Circulating microRNAs Showed Specific Responses according to Metabolic Syndrome Components and Sex of Adults from a Population-Based Study" Metabolites 13, no. 1: 2. https://doi.org/10.3390/metabo13010002
APA StyleBrandão-Lima, P. N., de Carvalho, G. B., Payolla, T. B., Sarti, F. M., Fisberg, R. M., Malcomson, F. C., Mathers, J. C., & Rogero, M. M. (2023). Circulating microRNAs Showed Specific Responses according to Metabolic Syndrome Components and Sex of Adults from a Population-Based Study. Metabolites, 13(1), 2. https://doi.org/10.3390/metabo13010002