
Influence of Altered Thyroid Hormone Mechanisms in the Progression of Metabolic Dysfunction Associated with Fatty Liver Disease (MAFLD): A Systematic Review

 ,
,  and
and
Abstract
:
1. Introduction
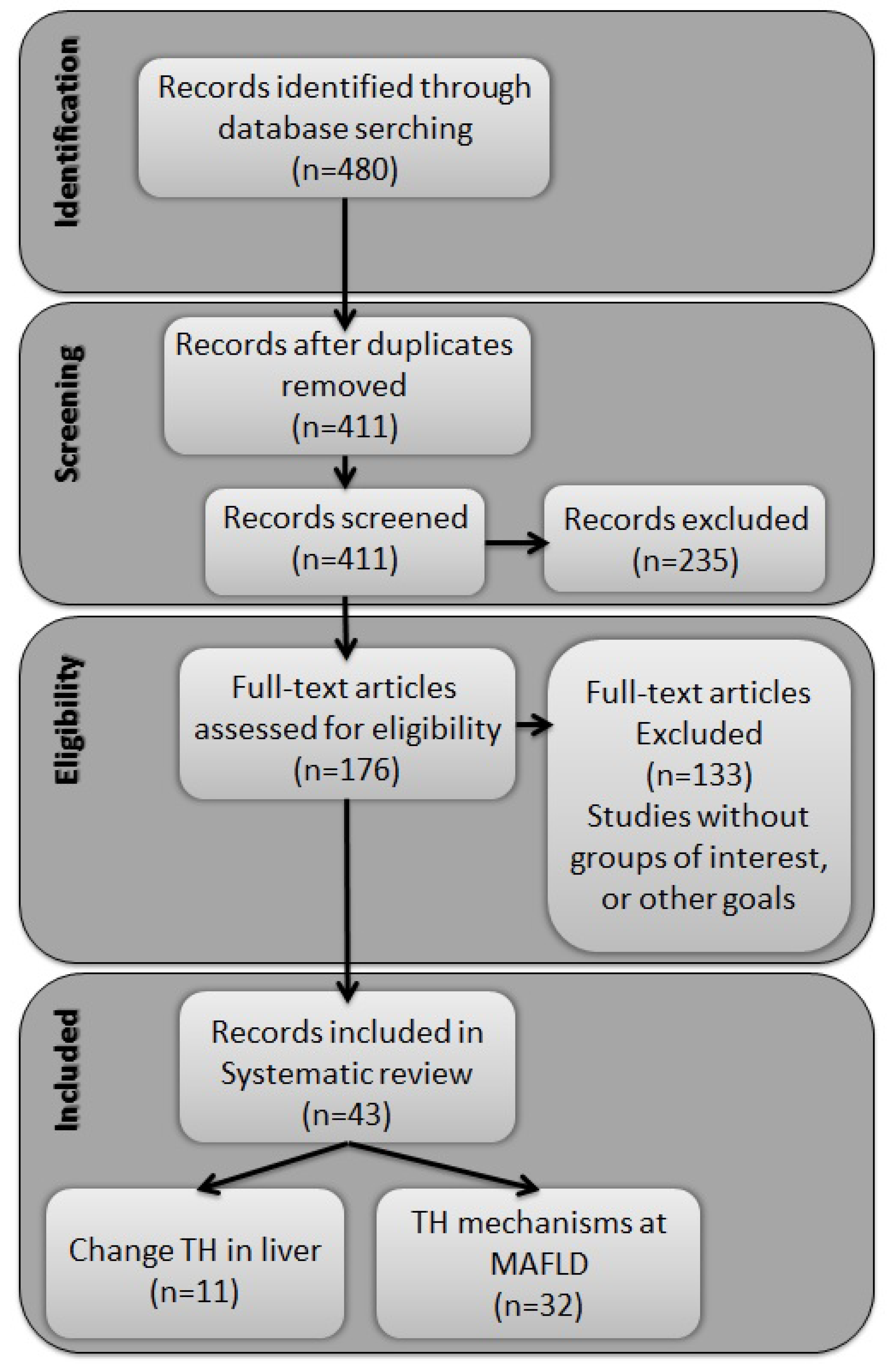
2. Results
2.1. Identified Records
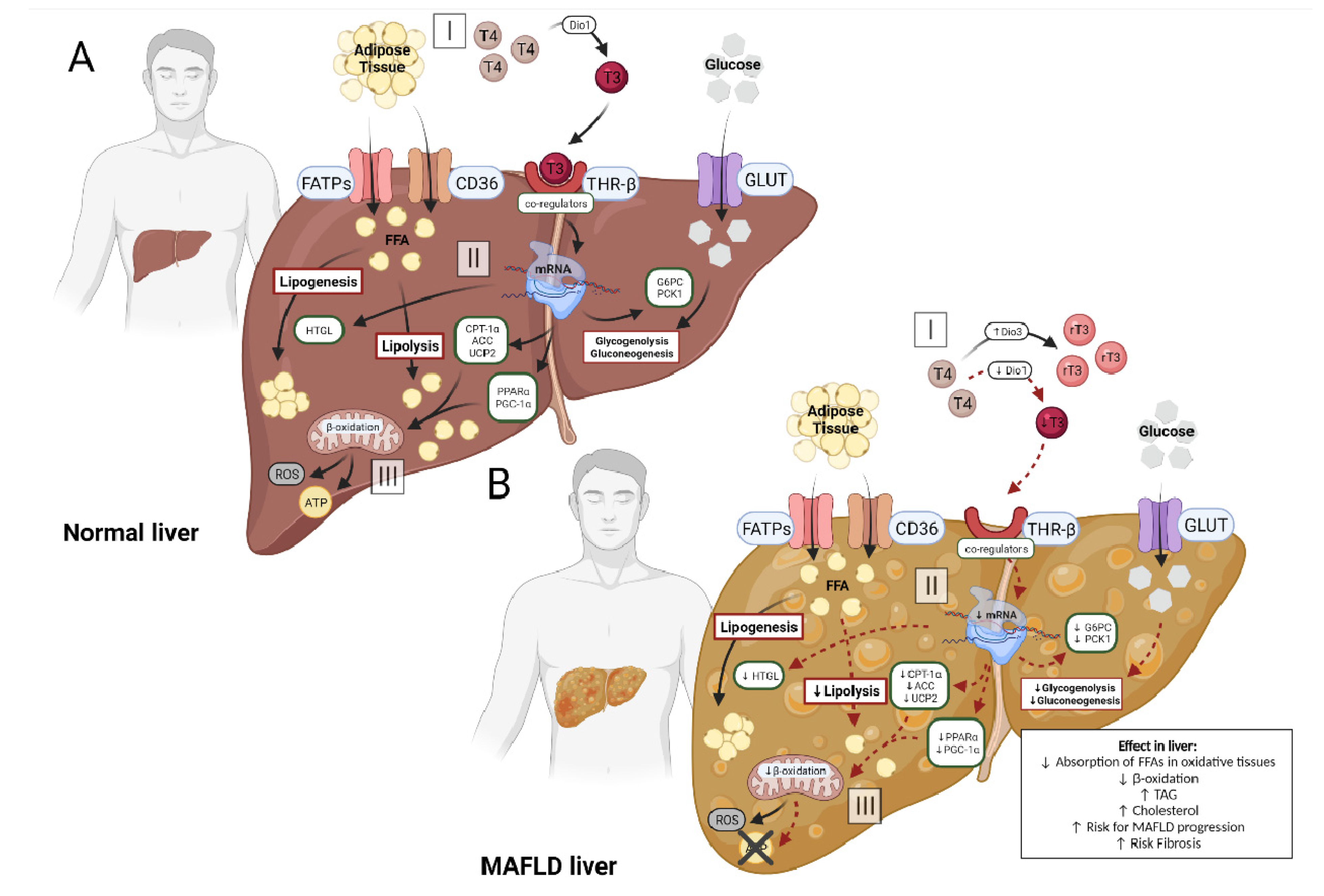
2.2. Altered Circulating Levels of Thyroid Hormone and the Liver

{kind=link}
{kind=link}
{kind=link}
| Manuscript | Sample | Study Design | MAFLD Assessment Technique | Groups | Serum Levels of THs | Effect on Liver |
|---|---|---|---|---|---|---|
| Klieverik et al. [14] (2009) | Rats | — | — | G1: Euthyroid G2: Hypothyroidism G3: Thyrotoxic | G2: ↓ THs G3: ↑ THs | Hypothyroidism ↓ Absorption of FFAs in oxidative tissues ↑ FFAs WAT absorption Thyrotoxic ↑ Absorption of oxidative tissue FFAs |
| Liangpunsakul et al. [18] (2003) | Human | Retrospective study | Biopsies and imaging | G1: Control G2: NASH | G2: ↓ T3 | Increased risk to NASH development |
| Chung et al. [19] (2012) | Human | Cross sectional | Imaging | G1: Euthyroidism with NAFLD G2: Subclinical hypothyroidism with NAFLD | G2: ↓ T3 | ↑ NAFLD prevalence |
| Bano et al. [15] (2016) | Human | Prospective cohort | Imaging | G1: NAFLD Euthyroidism G2: NAFLD Hypothyroidism G3: NAFLD Hyperthyroidism | G2: ↓ THs | ↑ Fibrosis ↑ Risk for NAFLD progression |
| Kim et al. [20] (2018) | Human | Cross sectional | Biopsies | G1: NAFLD strict-normal thyroid function G2: NAFLD low thyroid function | G2: ↓ THs | ↑ Fibrosis ↑ Risk of progression to NASH |
| Manka et al. [21] (2019) | Human | Retrospective study | Imaging | G1: NAFLD grade 1 G2: NAFLD grade 2 G3: NAFLD grade 3 G4: NAFLD grade 4 | G1: ↓ T3 G2: ↓↓ T3 G3: ↓↓ T3 G4: ↓↓↓ T3 | ↑ Risk of Fibrosis |
| Kim et al. [22] (2020) | Human | Retrospective study | Imaging | G1: NAFLD strict-normal thyroid function G2: NAFLD low thyroid function | G2: ↓ THs | ↑ Fibrosis ↑ Risk for all-cause and cardiovascular mortality |
| D’Ambrosio et al. [23] (2021) | Human | Retrospective Single-Center study | Biopsies | G1: NASH G2: Fibrosis G3: Steatosis | G1: ↓ T3 G2: ↓↓ T3 G3: ↓↓↓ T3 | ↑ Risk of NAFLD progression |
| Du et al. [16] (2021) | Human | Retrospective study | Imaging | G1: DM2 with NAFLD without Fibrosis G2: DM2 + NAFLD + Fibrosis | G2: ↓ T3 | ↑ Fibrosis |
| Chaves et al. [17] (2021) | Human | Cross sectional | Imaging | G1: R243Q mutation of the THR-β gene G2: Their wild-type first-degree relatives | G1: ↓ THR-β | ↑ Risk for NAFLD progression |
| Wang et al. [24] (2021) | Human | Cross sectional | Imaging | G1: Hypothyroidism G2: Hypothyroidism + NAFLD | G2: ↓ T3 | ↑ Liver FFAs |
2.3. Metabolic Action of Thyroid Hormone in Liver with MAFLD
| Manuscript | Sample | Study Design | NAFLD Assessment Technique | Groups | Treatment | Dose | THs | TH Target | Effect on Liver |
|---|---|---|---|---|---|---|---|---|---|
| Mustafa et al. [25] (2009) | Human | Cross sectional | Imaging | G1: Control G2: NASH | — | — | G2: ↓ T3 | ↑ serum MDA ↑ serum NO ↓ serum GSH ↓ serum GPx | ↑ Risk NASH progression |
| Krause et al. [26] (2018) | Human | Cohort Study | Biopsies | G1: NAFLD | — | — | G1: ↓ T3 | ↓ Dio1 mRNA ↓ THR-β mRNA | ↑ Hyperlipidemia ↑ Risk NASH progression |
| Harrison et al. [27] (2019) | Human | Randomized Controlled Trial | Biopsies | G1: NASH + Placebo G2: NASH + MGL-3196 | MGL-3196 | 80 mg | G2: ↑ THR-β | THR-β | ↓ Hepatic fat Restoration of Mitochondrial function |
| Harrison et al. [28] (2021) | Human | Extension Study | Biopsies | G1: NASH | MGL-3196 | 80 or 100 mg | G1: ↑ T3 and ↓ rT3 | THR-β | ↓ Risk NASH progression |
| Manuscript | Sample | Groups | Treatment | Dose | THs | TH Target | Effect on Liver |
|---|---|---|---|---|---|---|---|
| Nozaki et al. [31] (1992) | Cells | HepG2 cells | T3 | 0.1/μg/mL 0.2/μg/mL 0.3/μg/mL | ↑ T3 | ↑ HTGL mRNA ↑ Hepatic lipid hydrolysis | ↑ Lipogenesis |
| Zhang et al. [32] (2004) | Cells | HepG2 cells with Luciferase Vectors-CPT-1 | T3 | 100 nM | ↑ T3 | ↑ PGC-1α mRNA ↑ CPT-1α mRNA | ↑ Fatty acid β-oxidation |
| Grasselli et al. [29] (2011) | Cells | G1: Control G2: FFAs G3: FFAs + T2 G4: FFAs + T3 | T2 T3 | 10−7 to 10−5 M 10−7 to 10−5 M | G3: ↑ T2 G4: ↑ T3 | ↓ PPAR-δ and -γ mRNA ↓ SOD ↓ CAT | ↓ Excess fat ↓ TAG |
| Grasselli et al. [30] (2014) | Cells | G1: Control cells G2: Hepatoma cell + oleate/palmitate | T2 T3 | 10−6 M to 10−5 M | G2: ↑ T3 | ↑ UCP2 mRNA ↑ CPT-1 mRNA ↑ UCP2 protein ↑ CPT1 protein ↑ ROS ↓ CAT ↓ GSH | ↓ Extracellular TAG ↑ Fatty-acid oxidation ↓ NAFLD progression |
| Ness et al. [46] (1990) | Rats | G1: Normal G2: Hypophysectomized | T3 | 10 μg/100 g | G2: ↑ T3 | ↑ CYP7A1 mRNA | ↑ Cholesterol metabolism |
| Huang et al. [44] (1998) | Rats | G1: Hypothyroidism G2: Hyperthyroidism G3: Euthyroidism | — | — | G1: ↓ THs G2: ↑ THs | Hyperthyroid ↑ Acetyl-CoA carboxylase mRNA Hypothyroid ↓ Acetyl-CoA carboxylase mRNA | Hyperthyroid ↑ Hepatic lipogenesis Hypothyroidism ↓ Hepatic lipogenesis |
| Feng et al. [43] (2000) | Mice | G1: Control G2: Hyperthyroid | — | — | G2: ↑ T3 + THR | ↑ G6PC ↑ PCK1 | ↑ Glycogenolysis ↑ Gluconeogenesis |
| Jackson-Hayes et al. [53] (2003) | Mice | Transgenics (CPT-1α-luciferase) with/without the 1st intron of the CPT-1α gene | — | — | ↑ T3 + THR | ↑ CPT-1α gene | ↑ Fatty acid β-oxidation |
| Noguchi-Yachide et al. [49] (2007) | Mice | G1: Euthyroidism G2: Thyrotoxic G3: Hypothyroidism | — | — | G2: ↑ T3 + THR | ↑ LXR-α mRNA ↑ CYP7A1 mRNA | Lipid homeostasis |
| Liu et al. [52] (2007) | Mice | G1: WT control G2: Mutation in THR | — | — | G2: ↓ T3 + THR | ↓ PPARα protein | ↓ Fatty acid β-oxidation |
| Lopez et al. [45] (2007) | Rats | G1: Normal G2: Hypophysectomized G3: Thyroidectomy | T3 | 10 μg/100 g | G3: ↑ T3 | ↑ LDL receptor mRNA | ↑ FFA absorption |
| Cable et al. [55] (2009) | Rats | G1: NASH + Vehicle G2: NASH + T3 G3: NASH + MB07811 | MB07811 T3 | 1–50 mg/kg/day 650 μg/kg/day | G2: ↑ T3 G3: ↑ T3 + THR | ↑ CPT-1 mRNA ↑ PGC-1α mRNA | ↑ Mitochondrial β-oxidation |
| Mollica et al. [38] (2009) | Rats | G1: Control G2: HFD G3: HFD + T2 | T2 | 25 μg/100 g | G3: ↑ T2 | ↑ PPAR-α ↑ CPT-1 | ↑ mitochondrial respiration ↓ degree of steatosis |
| Adams et al. [50] (2010) | Mice | G1: C57BL/6 control G2: C57BL/6 with T3 | PBS T3 | 500 μg/kg | G2: ↑ T3 | ↑ FGF21 mRNA | ↑ lipolysis ↑ hepatic fatty acid oxidation |
| Sousa et al. [48] (2011) | Rats | G1: Euthyroidism G2: Hypothyroidism | Soybean oil Fish oil | 0.5 mL | G2: ↓ T3 | ↑ PPARα protein ↓ D1 mRNA | ↓ Serum triglycerides ↓ Hepatic TAG levels |
| Grasselli et al. [34] (2012) | Rats | G1: DP G2: HFD G3: HFD + T2 G4: DP + T2 | T2 | 25 μg/100 g | G3: ↑ T2 G4: ↑ T2 | ↑ PPARγ mRNA ↑ acyl-CoA oxidase mRNA | ↓ Inflammation ↓ Adipose triglyceride lipase ↑ FFA oxidation |
| Santana-Farré et al. [47] (2012) | Rats | G1: Neonatal Hypothyroidism G2: Age-matched euthyroid G3: Euthyroid weight paired | — | — | G1: ↓ T3 | ↑ PPARα mRNA ↓ LXR mRNA ↓ CD36 mRNA ↓ genes uptake Ags | ↓ Absorption of FFAs in the liver |
| Cavallo et al. [42] (2013) | Rats | G1: Euthyroid G2: Hypothyroid G3: Hypothyroid + T2 | T2 | 150 µg/100 g | G3: ↑ T2 | ↑ CPT-1 protein ↑ OXPHOS | ↑ Fatty acid β-oxidation ↓ Adiposity ↓ Dyslipidemia |
| Alonso-Merino et al. [51] (2016) | Cells Rats | G1: Euthyroid G2: Hyperthyroid | T4 T3 | T4 7 ng/g T3 35 ng/g | G2: ↑ T3 + THR | ↓ TGF-β mRNA | ↓ Fibrosis progression |
| Iannucci et al. [35] (2017) | Rats | G1: Control G2: HFD G3: HFD + T2 G4: HFD + T3 | T2 T3 | 25 µg/100 g 2.5 µg/100 g | G3: ↑ T3 + THR G4: ↑ T3 + THR | ↑ CPT-1α protein ↑ UCP2 protein ↑ p-ERK protein ↑ p-Akt protein | ↑ Lipolysis ↑ Autophagy ↑ Fatty acid β-oxidation |
| Senese et al. [39] (2017) | Rats | G1: Control G2: HFD G3: HFD + T2 G4: HFD + T3 | T2 T3 | 25 μg/100 g−1 2.5 μg/100 g−1 | G3: ↑ T2 G4: ↑ T3 | ↑ Dio1 mRNA ↑ THRβ mRNA | ↓ TAG ↓ Lipogenesis ↑ Fatty acid oxidation |
| Bruinstroop et al. [56] (2018) | Rats | G1: Control G2: MCD diet | — | — | G2: ↓ T3 | ↓ T3 hepatic ↓ Dio1 mRNA | ↑ NAFLD progression |
| Xia et al. [40] (2019) | Mice | G1: C57BL/6 control G2: C57BL/6 HFD G3: C57BL/6 HFD + Myr | Myricetin | 100 mg/kg−1 | G3: ↑ T4 and ↑ T3 | ↑ Dio1 mRNA ↑ Dio1 protein ↑ Dio1 activity ↑ THRβ mRNA ↑ THRβ protein | ↓ Hepatic steatosis ↑ Lipid metabolism |
| Luong et al. [37] (2020) | Rats | G1: Control G2: HFD G3: HFD + MGL-3169 (5.0 mg/kg) G4: HFD + MGL-3169 (1.5 mg/kg) G5: HFD + MGL-3169 (0.5 mg/kg) G6: HFD + T3 (0.5 mg/kg) | MGL-3196 T3 | 0.5–5.0 mg/kg | G3: ↑ T3 and ↑ THR G4: ↑ T3 and ↑ THR G5: ↑ T3 and ↑ THR G6: ↑ T3 and ↑ THR | ↑ Dio1 mRNA ↑ Me1 mRNA | ↓ Serum lipid profile ↑ FFA oxidation |
| Bruinstroop et al. [54] (2021) | Mice | G1: Control NCD G2: Control WDF G3: Dio1 LKD WDF | — | — | G3: ↓ T3 | ↓ Dio1 mRNA ↓ Dio1 activity | ↑ TAG ↑ Cholesterol ↑ Risk for NAFLD progression |
| Caddeo et al. [33] (2021) | Mice | G1: C57BL/6 G2: C57BL/6 + HFD G3: C57BL/6 + HFD + MGL-3196 G4: C57BL/6 + HFD + TG68 | MGL-3196 TG68 | 3 mg⋅kg−1 2.8 mg⋅kg−1 | G3: ↑ T3 G4: ↑ T3 | ↑ Dio1 mRNA ↑ THRsp mRNA | ↓ liver weight ↓ Serum TAG ↓ Plasma ALT ↓ Plasma AST |
| Kannt et al. [36] (2021) | Mice | G1: C57BL/6J + DP G2: C57BL/6J + HFD G3: C57BL/6J + HFD + Resmetirom | Resmetirom | 3 mg·kg−1 | G3: ↑ THR | ↑ Dio1 mRNA ↑ CYP7A1 mRNA ↑ Me1 mRNA | ↓ Serum lipid profile ↓ Liver weight ↓ NAFLD Score |
| Ge et al. [41] (2022) | Mice | G1: C57BL/6 control G2: C57BL/6 LOP G3: C57BL/6 HOP G4: C57BL/6 LOP + Dityr G5: C57BL/6 Dityr | — | — | G2: ↓ T3 G3: ↓ T3 G4: ↓ T3 G5: ↓ T3 | ↓ Dio1 mRNA ↓ THRβ mRNA ↓ CPT-1α mRNA ↓ PPARα mRNA ↓ PGC-1α mRNA ↓ CYP7A1 mRNA ↑ MDA ↑ ROS ↓ CAT ↓ GSH/GSSG | ↑ Risk NAFLD ↑ Inflammation ↑ Oxidative stress ↓ Hepatic energy metabolism ↑ Hepatic lipid synthesis ↓ Hepatic lipid catabolism ↓ Fatty-acid oxidation |
3. Discussion
3.1. THs Dependent Mechanisms in Hepatic Metabolism
3.2. Thyroid Hormone Metabolism Alterations and MAFLD
4. Methods
4.1. Protocol and Registration
4.2. Study Objectives
4.3. Eligibility Criteria
4.4. Search Strategy and Study Selection
4.5. Data Collection and Extraction
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Peeters, R.P.; Visser, T.J. Metabolism of Thyroid Hormone. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Bianco, A.C.; Salvatore, D.; Gereben, B.; Berry, M.J.; Larsen, P.R. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocr. Rev. 2002, 23, 38–89. [Google Scholar] [CrossRef] [PubMed]
- Ritter, M.J.; Amano, I.; Hollenberg, A.N. Thyroid Hormone Signaling and the Liver. Hepatology 2020, 72, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Duntas, L.H. Thyroid Disease and Lipids. Thyroid 2002, 12, 287–293. [Google Scholar] [CrossRef]
- Raftopoulos, Y.; Gagné, D.J.; Papasavas, P.; Hayetian, F.; Maurer, J.; Bononi, P.; Caushaj, P.F. Improvement of Hypothyroidism after Laparoscopic Roux-en-Y Gastric Bypass for Morbid Obesity. Obes. Surg. 2004, 14, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.A.; Singh, B.K.; Yen, P.M. Thyroid hormone regulation of hepatic lipid and carbohydrate metabolism. Trends Endocrinol. Metab. 2014, 25, 538–545. [Google Scholar] [CrossRef]
- Sinha, R.A.; Bruinstroop, E.; Singh, B.K.; Yen, P.M. Nonalcoholic Fatty Liver Disease and Hypercholesterolemia: Roles of Thyroid Hormones, Metabolites, and Agonists. Thyroid 2019, 29, 1173–1191. [Google Scholar] [CrossRef]
- Than, N.N.; Newsome, P.N. A concise review of non-alcoholic fatty liver disease. Atherosclerosis 2015, 239, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Arab, J.P.; Arrese, M.; Trauner, M. Recent Insights into the Pathogenesis of Nonalcoholic Fatty Liver Disease. Annu. Rev. Pathol. 2018, 13, 321–350. [Google Scholar] [CrossRef]
- Masarone, M.; Federico, A.; Abenavoli, L.M.; Loguercio, C.; Persico, M. Non Alcoholic Fatty Liver: Epidemiology and Natural History. Rev. Recent Clin. Trials 2015, 9, 126–133. [Google Scholar] [CrossRef]
- Eshraghian, A.; Jahromi, A.H. Non-alcoholic fatty liver disease and thyroid dysfunction: A systematic review. World J. Gastroenterol. 2014, 20, 8102–8109. [Google Scholar] [CrossRef]
- Guo, Z.; Li, M.; Han, B.; Qi, X. Association of non-alcoholic fatty liver disease with thyroid function: A systematic review and meta-analysis. Dig. Liver Dis. 2018, 50, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Li, B.; Zou, Y. The relationship between non-alcoholic fatty liver disease and hypothyroidism: A systematic review and meta-analysis. Medicine 2021, 100, e25738. [Google Scholar] [CrossRef] [PubMed]
- Klieverik, L.P.; Coomans, C.P.; Endert, E.; Sauerwein, H.P.; Havekes, L.M.; Voshol, P.J.; Rensen, P.C.N.; Romijn, J.A.; Kalsbeek, A.; Fliers, E. Thyroid Hormone Effects on Whole-Body Energy Homeostasis and Tissue-Specific Fatty Acid Uptake in vivo. Endocrinology 2009, 150, 5639–5648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, A.; Chaker, L.; Plompen, E.P.C.; Hofman, A.; Dehghan, A.; Franco, O.; Janssen, H.L.A.; Murad, S.D.; Peeters, R.P. Thyroid Function and the Risk of Nonalcoholic Fatty Liver Disease: The Rotterdam Study. J. Clin. Endocrinol. Metab. 2016, 101, 3204–3211. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Chai, S.; Zhao, X.; Sun, J.; Zhang, X.; Huo, L. Association between thyroid hormone levels and advanced liver fibrosis in patients with type 2 diabetes mellitus and non-alcoholic fatty liver disease. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 2399–2406. [Google Scholar] [CrossRef]
- Chaves, C.; Bruinstroop, E.; Refetoff, S.; Yen, P.M.; Anselmo, J.D. Increased Hepatic Fat Content in Patients with Resistance to Thyroid Hormone Beta. Thyroid 2021, 31, 1127–1134. [Google Scholar] [CrossRef]
- Liangpunsakul, S.; Chalasani, N. Is hypothyroidism a risk factor for non-alcoholic steatohepatitis? J. Clin. Gastroenterol. 2003, 37, 340–343. [Google Scholar] [CrossRef]
- Chung, G.E.; Kim, D.; Kim, W.; Yim, J.Y.; Park, M.J.; Kim, Y.J.; Yoon, J.H.; Lee, H.S. Non-alcoholic fatty liver disease across the spectrum of hypothyroidism. J. Hepatol. 2012, 57, 150–156. [Google Scholar] [CrossRef]
- Kim, D.; Kim, W.; Joo, S.K.; Bae, J.M.; Kim, J.H.; Ahmed, A. Subclinical Hypothyroidism and Low-Normal Thyroid Function Are Associated With Nonalcoholic Steatohepatitis and Fibrosis. Clin. Gastroenterol. Hepatol. 2018, 16, 123–131. [Google Scholar] [CrossRef]
- Manka, P.; Bechmann, L.; Best, J.; Sydor, S.; Claridge, L.C.; Coombes, J.D.; Canbay, A.; Moeller, L.; Gerken, G.; Wedemeyer, H.; et al. Low Free Triiodothyronine Is Associated with Advanced Fibrosis in Patients at High Risk for Nonalcoholic Steatohepatitis. Am. J. Dig. Dis. 2019, 64, 2351–2358. [Google Scholar] [CrossRef]
- Kim, D.; Vazquez-Montesino, L.M.; Escober, J.A.; Fernandes, C.T.; Cholankeril, G.; Loomba, R.; Harrison, S.A.; Younossi, Z.M.; Ahmed, A. Low Thyroid Function in Nonalcoholic Fatty Liver Disease Is an Independent Predictor of All-Cause and Cardiovascular Mortality. Am. J. Gastroenterol. 2020, 115, 1496–1504. [Google Scholar] [CrossRef]
- D’Ambrosio, R.; Campi, I.; Maggioni, M.; Perbellini, R.; Giammona, E.; Stucchi, R.; Borghi, M.; Degasperi, E.; De Silvestri, A.; Persani, L.; et al. The relationship between liver histology and thyroid function tests in patients with non-alcoholic fatty liver disease (NAFLD). PLoS ONE 2021, 16, e0249614. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wang, B.; Yang, Y.; Xu, J.; Hong, M.; Xia, M.; Li, X.; Gao, X. Thyroid function and non-alcoholic fatty liver disease in hyperthyroidism patients. BMC Endocr. Disord. 2021, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.H.A.; Ali, E.M.; Mohamed, T.; Abdou, H.I. Oxidative stress and thyroid hormones in patients with liver diseases. Eur. J. Intern. Med. 2009, 20, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Krause, C.; Grohs, M.; El Gammal, A.T.; Wolter, S.; Lehnert, H.; Mann, O.; Mittag, J.; Kirchner, H. Reduced expression of thyroid hormone receptor β in human nonalcoholic steatohepatitis. Endocr. Connect. 2018, 7, 1448–1456. [Google Scholar] [CrossRef] [Green Version]
- A Harrison, S.; Bashir, M.R.; Guy, C.D.; Zhou, R.; A Moylan, C.; Frias, J.P.; Alkhouri, N.; Bansal, M.B.; Baum, S.; A Neuschwander-Tetri, B.; et al. Resmetirom (MGL-3196) for the treatment of non-alcoholic steatohepatitis: A multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2019, 394, 2012–2024. [Google Scholar] [CrossRef]
- Harrison, S.A.; Bashir, M.; Moussa, S.E.; McCarty, K.; Frias, J.P.; Taub, R.; Alkhouri, N. Effects of Resmetirom on Noninvasive Endpoints in a 36-Week Phase 2 Active Treatment Extension Study in Patients With NASH. Hepatol. Commun. 2021, 5, 573–588. [Google Scholar] [CrossRef]
- Grasselli, E.; Voci, A.; Canesi, L.; De Matteis, R.; Goglia, F.; Cioffi, F.; Fugassa, E.; Gallo, G.; Vergani, L. Direct effects of iodothyronines on excess fat storage in rat hepatocytes. J. Hepatol. 2011, 54, 1230–1236. [Google Scholar] [CrossRef]
- Grasselli, E.; Voci, A.; Canesi, L.; Salis, A.; Damonte, G.; Compalati, A.D.; Goglia, F.; Gallo, G.; Vergani, L. 3,5-Diiodo-L-Thyronine Modifies the Lipid Droplet Composition in a Model of Hepatosteatosis. Cell. Physiol. Biochem. 2014, 33, 344–356. [Google Scholar] [CrossRef]
- Nozaki, S.; Shimomura, I.; Funahashi, T.; Menju, M.; Kubo, M.; Matsuzawa, Y. Stimulation of the activity and mRNA level of hepatic triacylglycerol lipase by triiodothyronine in HepG2 cells. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1992, 1127, 298–302. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, K.; Song, S.; Elam, M.B.; Cook, G.A.; Park, E.A. Peroxisomal proliferator-activated receptor-gamma coactivator-1 alpha (PGC-1 alpha) enhances the thyroid hormone induction of carnitine palmitoyltransferase I (CPT-I alpha). J. Biol. Chem. 2004, 279, 53963–53971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caddeo, A.; Kowalik, M.A.; Serra, M.; Runfola, M.; Bacci, A.; Rapposelli, S.; Columbano, A.; Perra, A. TG68, a Novel Thyroid Hormone Receptor-β Agonist for the Treatment of NAFLD. Int. J. Mol. Sci. 2021, 22, 13105. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, E.; Voci, A.; Demori, I.; Canesi, L.; De Matteis, R.; Goglia, F.; Lanni, A.; Gallo, G.; Vergani, L. 3,5-Diiodo-l-thyronine modulates the expression of genes of lipid metabolism in a rat model of fatty liver. J. Endocrinol. 2011, 212, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannucci, L.F.; Cioffi, F.; Senese, R.; Goglia, F.; Lanni, A.; Yen, P.M.; Sinha, R.A. Metabolomic analysis shows differential hepatic effects of T(2) and T(3) in rats after short-term feeding with high fat diet. Sci. Rep. 2017, 7, 2023. [Google Scholar] [CrossRef] [Green Version]
- Kannt, A.; Wohlfart, P.; Madsen, A.N.; Veidal, S.S.; Feigh, M.; Schmoll, D. Activation of thyroid hormone receptor-β improved disease activity and metabolism independent of body weight in a mouse model of non-alcoholic steatohepatitis and fibrosis. J. Cereb. Blood Flow Metab. 2021, 178, 2412–2423. [Google Scholar] [CrossRef]
- Luong, X.G.; Stevens, S.K.; Jekle, A.; Lin, T.-I.; Gupta, K.; Misner, D.; Chanda, S.; Mukherjee, S.; Williams, C.; Stoycheva, A.; et al. Regulation of gene transcription by thyroid hormone receptor β agonists in clinical development for the treatment of non-alcoholic steatohepatitis (NASH). PLoS ONE 2020, 15, e0240338. [Google Scholar] [CrossRef]
- Mollica, M.P.; Lionetti, L.; Moreno, M.; Lombardi, A.; De Lange, P.; Antonelli, A.; Lanni, A.; Cavaliere, G.; Barletta, A.; Goglia, F. 3,5-diiodo-l-thyronine, by modulating mitochondrial functions, reverses hepatic fat accumulation in rats fed a high-fat diet. J. Hepatol. 2009, 51, 363–370. [Google Scholar] [CrossRef]
- Senese, R.; Cioffi, F.; de Lange, P.; Leanza, C.; Iannucci, L.F.; Silvestri, E.; Moreno, M.; Lombardi, A.; Goglia, F.; Lanni, A. Both 3,5-Diiodo-L-Thyronine and 3,5,3′-Triiodo-L-Thyronine Prevent Short-term Hepatic Lipid Accumulation via Distinct Mechanisms in Rats Being Fed a High-Fat Diet. Front. Physiol. 2017, 8, 706. [Google Scholar] [CrossRef]
- Xia, S.F.; Qiu, Y.Y.; Chen, L.M.; Jiang, Y.Y.; Huang, W.; Xie, Z.X.; Tang, X.; Sun, J. Myricetin alleviated hepatic steatosis by acting on microRNA-146b/thyroid hormone receptor b pathway in high-fat diet fed C57BL/6J mice. Food Funct. 2019, 10, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Yang, Y.; Jiang, Y.; Feng, C.; Li, B.; Sun, J.; Tang, X.; Shi, Y.; Le, G. Oxidized Pork Induces Hepatic Steatosis by Impairing Thyroid Hormone Function in Mice. Mol. Nutr. Food Res. 2021, 66, e2100602. [Google Scholar] [CrossRef]
- Cavallo, A.; Priore, P.; Gnoni, G.V.; Papa, S.; Zanotti, F.; Gnoni, A. 3,5-Diiodo-L-Thyronine Administration To Hypothyroid Rats Rapidly Enhances Fatty Acid Oxidation Rate and Bioenergetic Parameters in Liver Cells. PLoS ONE 2013, 8, e52328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Jiang, Y.; Meltzer, P.; Yen, P.M. Thyroid hormone regulation of hepatic genes in vivo detected by complementary DNA microarray. Mol. Endocrinol. 2000, 14, 947–955. [Google Scholar] [CrossRef]
- Huang, C.; Freake, H.C. Thyroid hormone regulates the acetyl-CoA carboxylase PI promoter. Biochem. Biophys. Res. Commun. 1998, 249, 704–708. [Google Scholar] [CrossRef] [PubMed]
- Lopez, D.; Socarrás, J.F.A.; Bedi, M.; Ness, G.C. Activation of the hepatic LDL receptor promoter by thyroid hormone. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2007, 1771, 1216–1225. [Google Scholar] [CrossRef]
- Ness, G.C.; Pendleton, L.C.; Li, Y.C.; Chiang, J.Y. Effect of thyroid hormone on hepatic cholesterol 7α hydroxylase, LDL receptor, HMG-CoA reductase, farnesyl pyrophosphate synthetase and apolipoprotein A-I mRNA levels in hypophysectomized rats. Biochem. Biophys. Res. Commun. 1990, 172, 1150–1156. [Google Scholar] [CrossRef]
- Santana-Farré, R.; Mirecki-Garrido, M.; Bocos, C.; Henríquez-Hernández, L.A.; Kahlon, N.; Herrera, E.; Norstedt, G.; Parini, P.; Flores-Morales, A.; Fernández-Pérez, L. Influence of Neonatal Hypothyroidism on Hepatic Gene Expression and Lipid Metabolism in Adulthood. PLoS ONE 2012, 7, e37386. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.L.; Cordeiro, A.; Oliveira, L.S.; de Paula, G.S.M.; Faustino, L.C.; Ortiga-Carvalho, T.M.; Oliveira, K.J.; Pazos-Moura, C.C. Thyroid hormone contributes to the hypolipidemic effect of polyunsaturated fatty acids from fish oil: In vivo evidence for cross talking mechanisms. J. Endocrinol. 2011, 211, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Noguchi-Yachide, T.; Aoyama, A.; Makishima, M.; Miyachi, H.; Hashimoto, Y. Liver X receptor antagonists with a phthalimide skeleton derived from thalidomide-related glucosidase inhibitors. Bioorganic Med. Chem. Lett. 2007, 17, 3957–3961. [Google Scholar] [CrossRef]
- Adams, A.C.; Astapova, I.; Fisher, F.M.; Badman, M.K.; Kurgansky, K.E.; Flier, J.S.; Hollenberg, A.N.; Maratos-Flier, E. Thyroid Hormone Regulates Hepatic Expression of Fibroblast Growth Factor 21 in a PPARα-dependent Manner. J. Biol. Chem. 2010, 285, 14078–14082. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Merino, E.; Martin Orozco, R.; Ruiz-Llorente, L.; Martinez-Iglesias, O.A.; Velasco-Martin, J.P.; Montero-Pedrazuela, A.; Fanjul-Rodriguez, L.; Contreras-Jurado, C.; Regadera, J.; Aranda, A. Thyroid hormones inhibit TGF-beta signaling and attenuate fibrotic responses. Proc. Natl. Acad. Sci. USA 2016, 113, 3451–3460. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Heymann, R.S.; Moatamed, F.; Schultz, J.J.; Sobel, D.; Brent, G.A. A mutant thyroid hormone receptor alpha antagonizes peroxisome proliferator-activated receptor alpha signaling in vivo and impairs fatty acid oxidation. Endocrinology 2007, 148, 1206–1217. [Google Scholar]
- Jackson-Hayes, L.; Song, S.; Lavrentyev, E.N.; Jansen, M.S.; Hillgartner, F.B.; Tian, L.; Wood, P.A.; Cook, G.A.; Park, E.A. A Thyroid Hormone Response Unit Formed between the Promoter and First Intron of the Carnitine Palmitoyltransferase-Iα Gene Mediates the Liver-specific Induction by Thyroid Hormone. J. Biol. Chem. 2003, 278, 7964–7972. [Google Scholar] [PubMed] [Green Version]
- Bruinstroop, E.; Zhou, J.; Tripathi, M.; Yau, W.W.; Boelen, A.; Singh, B.K.; Yen, P.M. Early induction of hepatic deiodinase type 1 inhibits hepatosteatosis during NAFLD progression. Mol. Metab. 2021, 53, 101266. [Google Scholar] [PubMed]
- Cable, E.E.; Finn, P.D.; Stebbins, J.W.; Hou, J.; Ito, B.R.; van Poelje, P.D.; Linemeyer, D.L.; Erion, M.D. Reduction of hepatic steatosis in rats and mice after treatment with a liver-targeted thyroid hormone receptor agonist. Hepatology 2009, 49, 407–417. [Google Scholar] [PubMed]
- Bruinstroop, E.; Dalan, R.; Cao, Y.; Bee, Y.M.; Chandran, K.; Cho, L.W.; Soh, S.B.; Teo, E.K.; Toh, S.-A.; Leow, M.K.S.; et al. Low-Dose Levothyroxine Reduces Intrahepatic Lipid Content in Patients With Type 2 Diabetes Mellitus and NAFLD. J. Clin. Endocrinol. Metab. 2018, 103, 2698–2706. [Google Scholar]
- Mendoza, A.; Hollenberg, A.N. New insights into thyroid hormone action. Pharmacol. Ther. 2017, 173, 135–145. [Google Scholar]
- Ishizuka, T.; Lazar, M.A. The Nuclear Receptor Corepressor Deacetylase Activating Domain Is Essential for Repression by Thyroid Hormone Receptor. Mol. Endocrinol. 2005, 19, 1443–1451. [Google Scholar]
- Shimizu, H.; Astapova, I.; Ye, F.; Bilban, M.; Cohen, R.N.; Hollenberg, A.N. NCoR1 and SMRT Play Unique Roles in Thyroid Hormone Action in Vivo. Mol. Cell. Biol. 2015, 35, 555–565. [Google Scholar]
- Coburn, C.T.; Hajri, T.; Ibrahimi, A.; Abumrad, N.A. Role of CD36 in Membrane Transport and Utilization of Long-Chain Fatty Acids by Different Tissues. J. Mol. Neurosci. 2001, 16, 117–122. [Google Scholar]
- Brenta, G. Why can insulin resistance be a natural consequence of thyroid dysfunction? J. Thyroid Res. 2011, 2011, 152850. [Google Scholar]
- Liu, Y.-Y.; Brent, G.A. Thyroid hormone crosstalk with nuclear receptor signaling in metabolic regulation. Trends Endocrinol. Metab. 2010, 21, 166–173. [Google Scholar] [PubMed] [Green Version]
- Yang, L.; Li, P.; Fu, S.; Calay, E.S.; Hotamisligil, G.S. Defective hepatic autophagy in obesity promotes ER stress and causes insulin resistance. Cell Metab. 2010, 11, 467–478. [Google Scholar] [PubMed] [Green Version]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [PubMed] [Green Version]
- Wierzbicki, M.; Chabowski, A.; Zendzian-Piotrowska, M.; Górski, J. Differential effects of in vivo PPAR alpha and gamma activation on fatty acid transport proteins expression and lipid content in rat liver. J. Physiol. Pharmacol. 2009, 60, 99–106. [Google Scholar]
- Crunkhorn, S.; Patti, M.-E. Links between Thyroid Hormone Action, Oxidative Metabolism, and Diabetes Risk? Thyroid 2008, 18, 227–237. [Google Scholar]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. BMJ 2009, 339, b2535. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marschner, R.A.; Arenhardt, F.; Ribeiro, R.T.; Wajner, S.M. Influence of Altered Thyroid Hormone Mechanisms in the Progression of Metabolic Dysfunction Associated with Fatty Liver Disease (MAFLD): A Systematic Review. Metabolites 2022, 12, 675. https://doi.org/10.3390/metabo12080675
Marschner RA, Arenhardt F, Ribeiro RT, Wajner SM. Influence of Altered Thyroid Hormone Mechanisms in the Progression of Metabolic Dysfunction Associated with Fatty Liver Disease (MAFLD): A Systematic Review. Metabolites. 2022; 12(8):675. https://doi.org/10.3390/metabo12080675
Chicago/Turabian StyleMarschner, Rafael Aguiar, Fernanda Arenhardt, Rafael Teixeira Ribeiro, and Simone Magagnin Wajner. 2022. "Influence of Altered Thyroid Hormone Mechanisms in the Progression of Metabolic Dysfunction Associated with Fatty Liver Disease (MAFLD): A Systematic Review" Metabolites 12, no. 8: 675. https://doi.org/10.3390/metabo12080675
APA StyleMarschner, R. A., Arenhardt, F., Ribeiro, R. T., & Wajner, S. M. (2022). Influence of Altered Thyroid Hormone Mechanisms in the Progression of Metabolic Dysfunction Associated with Fatty Liver Disease (MAFLD): A Systematic Review. Metabolites, 12(8), 675. https://doi.org/10.3390/metabo12080675