
An Update on Sphingolipidomics: Is Something Still Missing? Some Considerations on the Analysis of Complex Sphingolipids and Free-Sphingoid Bases in Plasma and Red Blood Cells

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
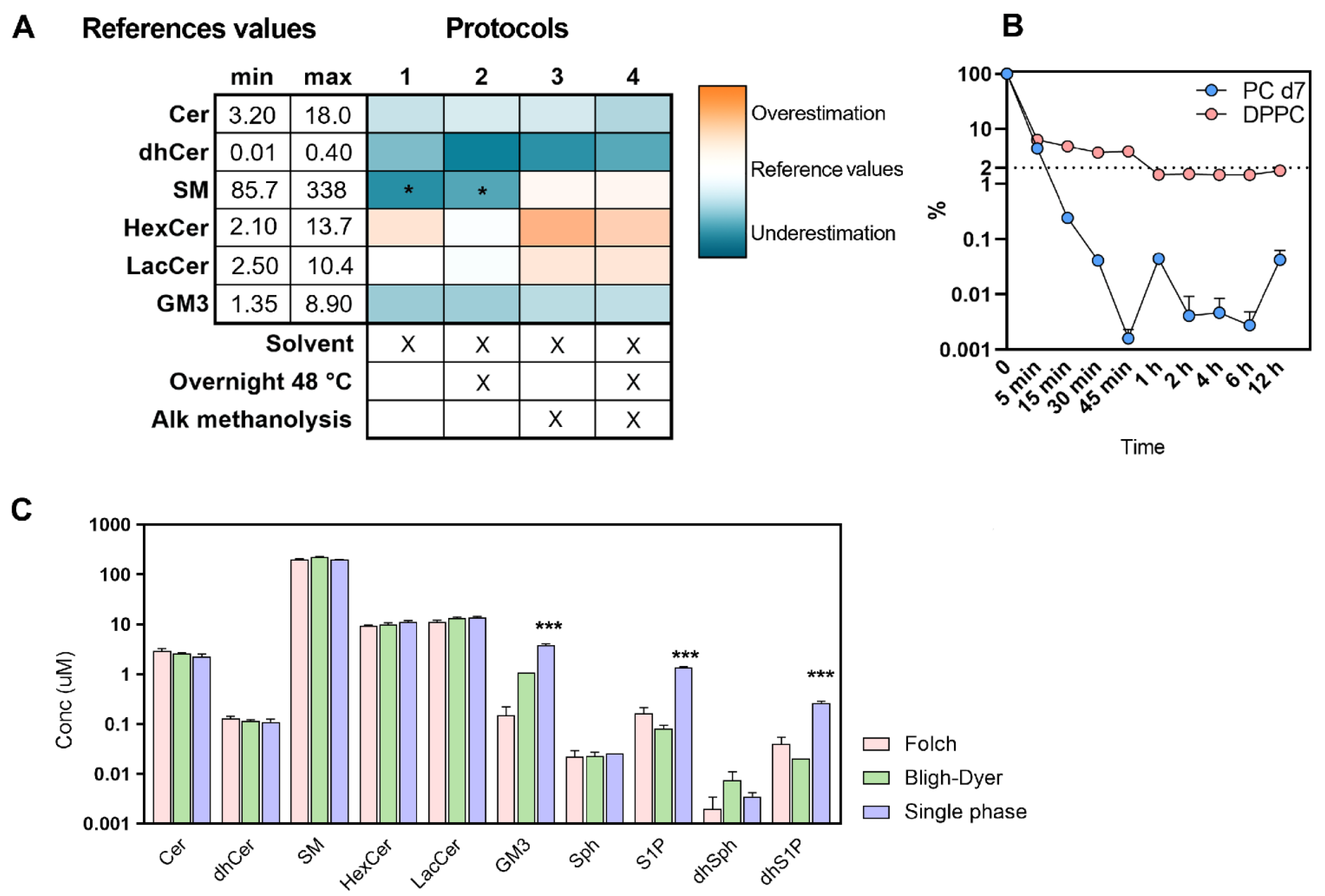
2.1. Set Up of the Extraction Procedure
2.2. Choosing the Best Analytical Condition
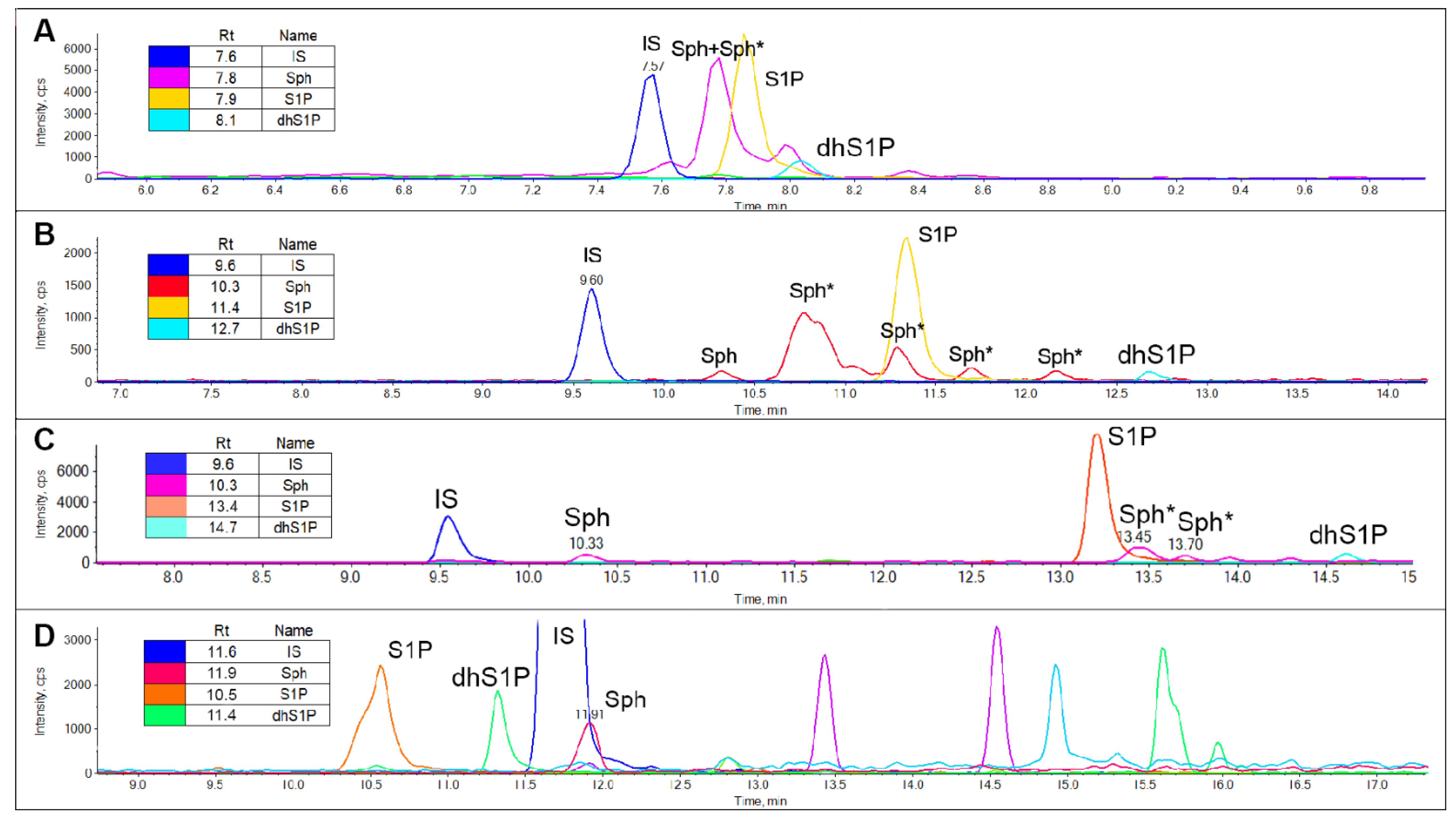
2.2.1. Sphingoid Bases Derivatization
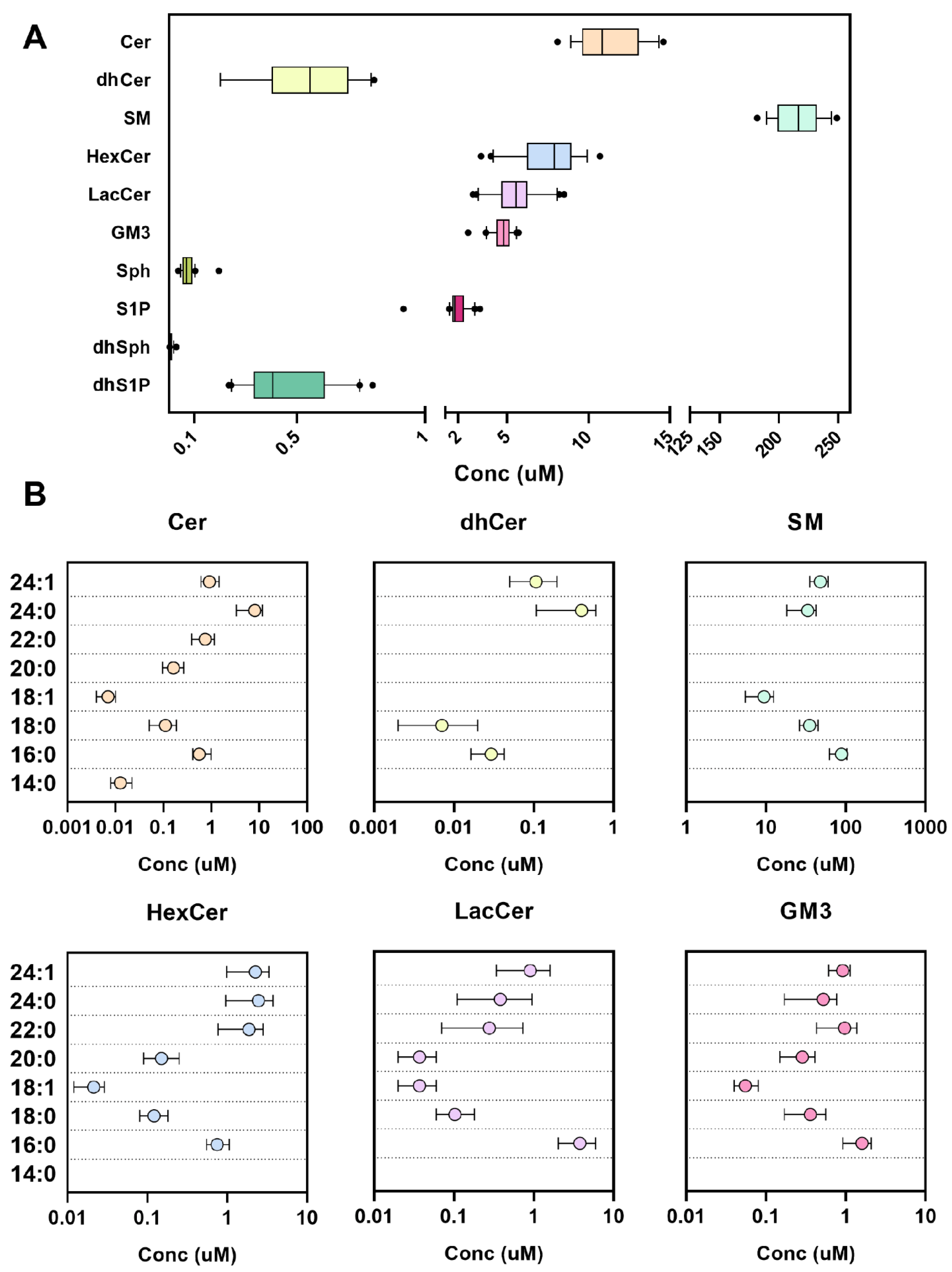
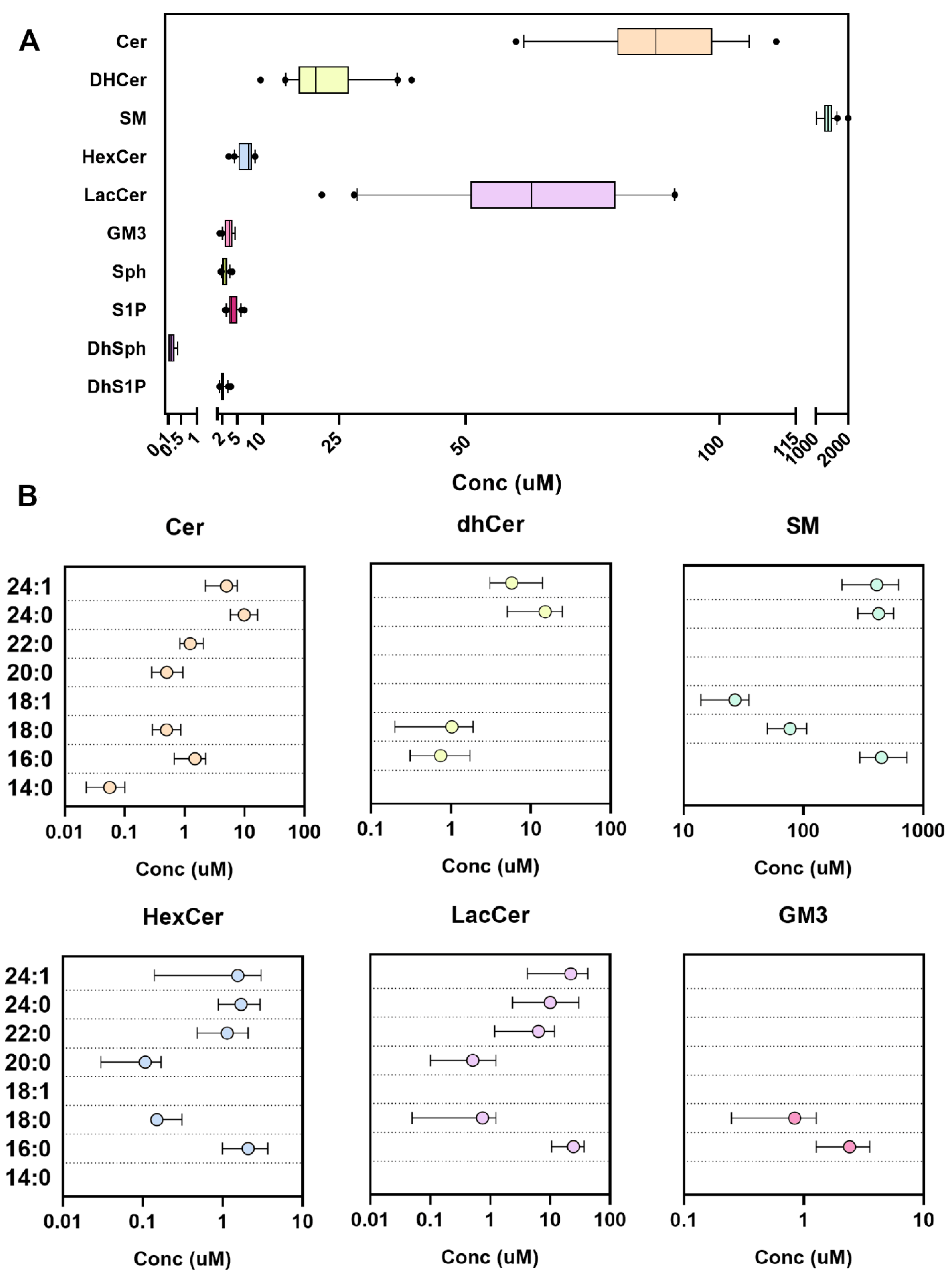
2.3. Performance in Human Plasma and Red Blood Cells
3. Materials and Methods
3.1. Biological Samples from Healthy Volunteers
3.2. Chemicals and Reagents
3.3. LC-MS/MS
3.3.1. Sphingolipids and Glycosphingolipids
3.3.2. Free Sphingoid Bases
3.3.3. Sphingoid Bases as Phenylthiourea Derivatives
3.4. Extraction Procedures
3.5. Derivatization of Free Sphingoid Bases
3.6. Condition for Alkaline Methanolysis
3.7. Comparison between Traditional Biphasic and Monophasic Extractions
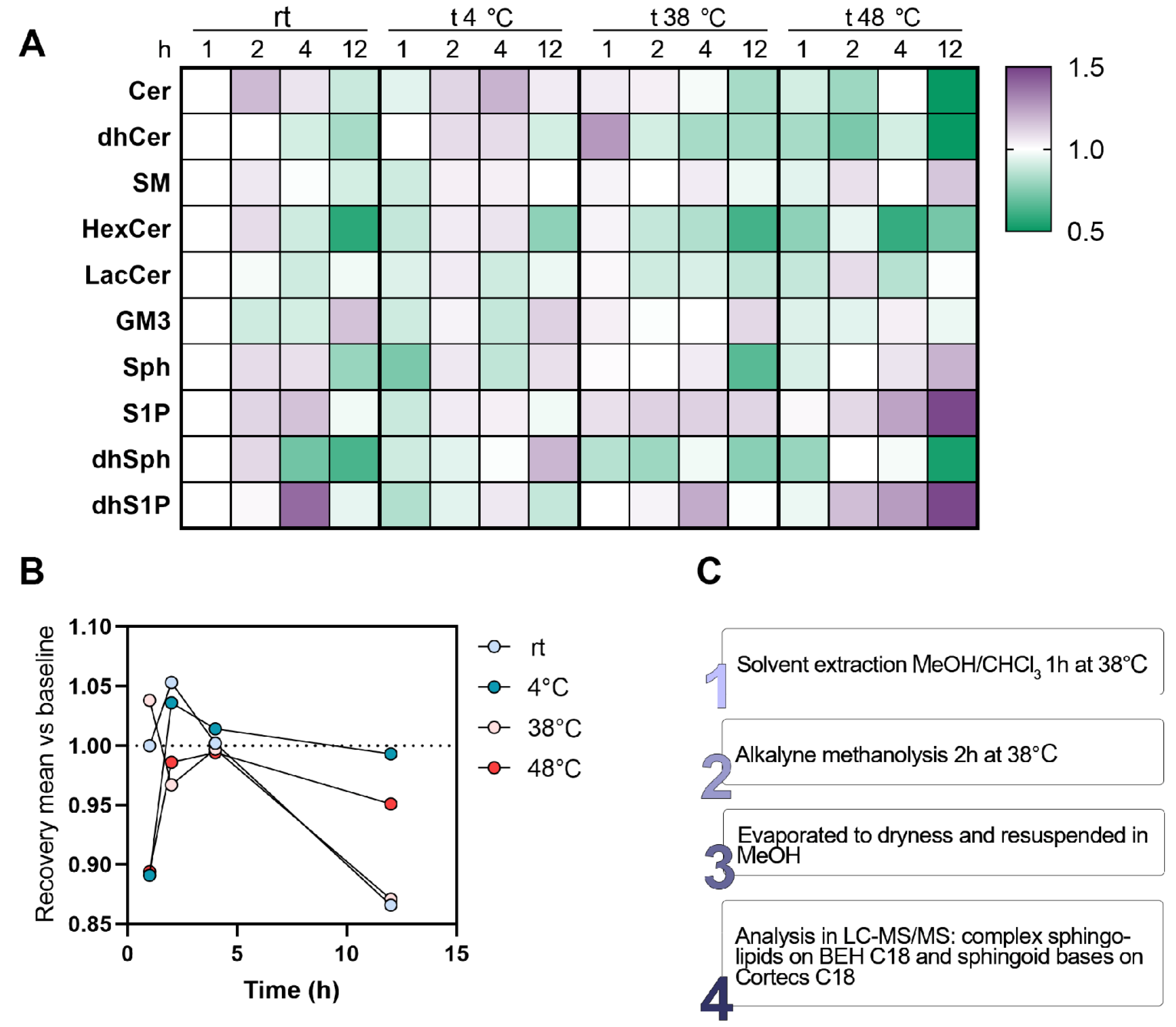
3.8. Time and Temperature for Isolating Sphingolipids from a Biological Matrix
3.9. Operating Protocol for Plasma Samples
3.10. Red Blood Cells Protocol
3.11. Methods Performances
3.12. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Merrill, A.H.; Schmelz, E.M.; Dillehay, D.L.; Spiegel, S.; Shayman, J.A.; Schroeder, J.J.; Riley, R.T.; Voss, K.A.; Wang, E. Sphingolipids—The Enigmatic Lipid Class: Biochemistry, Physiology, and Pathophysiology. Toxicol. Appl. Pharmacol. 1997, 142, 208–225. [Google Scholar] [CrossRef]
- Futerman, A.H. Sphingolipids. In Biochemistry of Lipids, Lipoproteins and Membranes: Sixth Edition; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 297–326. ISBN 978-0-44463-438-2. [Google Scholar]
- Dowhan, W.; Bogdanov, M.; Mileykovskaya, E. Functional Roles of Lipids in Membranes. In Biochemistry of Lipids, Lipoproteins and Membranes: Sixth Edition; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 1–40. ISBN 978-0-44463-438-2. [Google Scholar]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and Their Metabolism in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Mullen, T.D.; Jenkins, R.W.; Clarke, C.J.; Bielawski, J.; Hannun, Y.A.; Obeid, L.M. Ceramide Synthase-Dependent Ceramide Generation and Programmed Cell Death: Involvement of Salvage Pathway in Regulating Postmitochondrial Events. J. Biol. Chem. 2011, 286, 15929–15942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Spassieva, S.D.; Bieberich, E. Ceramide and S1P Signaling in Embryonic Stem Cell Differentiation. Methods Mol. Biol. 2018, 1697, 153–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohno, M.; Momoi, M.; Oo, M.L.; Paik, J.-H.; Lee, Y.-M.; Venkataraman, K.; Ai, Y.; Ristimaki, A.P.; Fyrst, H.; Sano, H.; et al. Intracellular Role for Sphingosine Kinase 1 in Intestinal Adenoma Cell Proliferation. Mol. Cell. Biol. 2006, 26, 7211–7223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patwardhan, G.A.; Beverly, L.J.; Siskind, L.J. Sphingolipids and Mitochondrial Apoptosis. J. Bioenerg. Biomembr. 2016, 48, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Canela, N.; Herrero, P.; Mariné, S.; Nadal, P.; Ras, M.R.; Rodríguez, M.Á.; Arola, L. Analytical Methods in Sphingolipidomics: Quantitative and Profiling Approaches in Food Analysis. J. Chromatogr. A 2016, 1428, 16–38. [Google Scholar] [CrossRef]
- Haynes, C.A.; Allegood, J.C.; Park, H.; Sullards, M.C. Sphingolipidomics: Methods for the Comprehensive Analysis of Sphingolipids. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2009, 877, 2696–2708. [Google Scholar] [CrossRef] [Green Version]
- Morano, C.; Roda, G.; Paroni, R.; Dei Cas, M. Tip-Tip Filtration Ameliorates Single-Phase Extraction Methods for Plasma Large-Scale Lipidomics Analysis. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2022, 1189, 123099. [Google Scholar] [CrossRef]
- Merrill, A.H.; Sullards, M.C.; Allegood, J.C.; Kelly, S.; Wang, E. Sphingolipidomics: High-Throughput, Structure-Specific, and Quantitative Analysis of Sphingolipids by Liquid Chromatography Tandem Mass Spectrometry. Methods 2005, 36, 207–224. [Google Scholar] [CrossRef]
- Dalmau, N.; Jaumot, J.; Tauler, R.; Bedia, C. Epithelial-to-Mesenchymal Transition Involves Triacylglycerol Accumulation in DU145 Prostate Cancer Cells. Mol. Biosyst. 2015, 11, 3397–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houjou, T.; Yamatani, K.; Nakanishi, H.; Imagawa, M.; Shimizu, T.; Taguchi, R. Rapid and Selective Identification of Molecular Species in Phosphatidylcholine and Sphingomyelin by Conditional Neutral Loss Scanning and MS3. Rapid Commun. Mass Spectrom. 2004, 18, 3123–3130. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.W.K.; Braidy, N.; Pickford, R.; Sachdev, P.S.; Poljak, A. Comparison of Single Phase and Biphasic Extraction Protocols for Lipidomic Studies Using Human Plasma. Front. Neurol. 2019, 10, 879. [Google Scholar] [CrossRef]
- Dei Cas, M.; Zulueta, A.; Mingione, A.; Caretti, A.; Ghidoni, R.; Signorelli, P.; Paroni, R. An Innovative Lipidomic Workflow to Investigate the Lipid Profile in a Cystic Fibrosis Cell Line. Cells 2020, 9, 1197. [Google Scholar] [CrossRef]
- Hammad, S.M.; Harden, O.C.; Wilson, D.A.; Twal, W.O.; Nietert, P.J.; Oates, J.C. Plasma Sphingolipid Profile Associated with Subclinical Atherosclerosis and Clinical Disease Markers of Systemic Lupus Erythematosus: Potential Predictive Value. Front. Immunol. 2021, 12, 694318. [Google Scholar] [CrossRef]
- Bui, H.H.; Leohr, J.K.; Kuo, M.S. Analysis of Sphingolipids in Extracted Human Plasma Using Liquid Chromatography Electrospray Ionization Tandem Mass Spectrometry. Anal. Biochem. 2012, 423, 187–194. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Han, Y.; Wang, T.; Wang, Y.; Gong, J.; Gao, K.; Chen, W.; Li, W.; Zhang, H.; et al. A Simple and Rapid Method for Extraction and Measurement of Circulating Sphingolipids Using LC-MS/MS: A Targeted Lipidomic Analysis. Anal. Bioanal. Chem. 2022, 414, 2041–2054. [Google Scholar] [CrossRef]
- Scherer, M.; Böttcher, A.; Schmitz, G.; Liebisch, G. Sphingolipid Profiling of Human Plasma and FPLC-Separated Lipoprotein Fractions by Hydrophilic Interaction Chromatography Tandem Mass Spectrometry. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 68–75. [Google Scholar] [CrossRef]
- Chan, R.B.; Perotte, A.J.; Zhou, B.; Liong, C.; Shorr, E.J.; Marder, K.S.; Kang, U.J.; Waters, C.H.; Levy, O.A.; Xu, Y.; et al. Elevated GM3 Plasma Concentration in Idiopathic Parkinson’s Disease: A Lipidomic Analysis. PLoS ONE 2017, 12, e0172348. [Google Scholar] [CrossRef] [PubMed]
- Hammad, S.M.; Pierce, J.S.; Soodavar, F.; Smith, K.J.; Al Gadban, M.M.; Rembiesa, B.; Klein, R.L.; Hannun, Y.A.; Bielawski, J.; Bielawska, A. Blood Sphingolipidomics in Healthy Humans: Impact of Sample Collection Methodology. J. Lipid Res. 2010, 51, 3074–3087. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Liu, D.; Xin, B.; Cechner, K.; Zhou, X.; Wang, H.; Zhou, A. Quantification of Monosialogangliosides in Human Plasma through Chemical Derivatization for Signal Enhancement in LC-ESI-MS. Anal. Chim. Acta 2016, 929, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Schmidt, R.; Geisslinger, G. LC-MS/MS-Analysis of Sphingosine-1-Phosphate and Related Compounds in Plasma Samples. Prostagland. Other Lipid Mediat. 2006, 81, 162–170. [Google Scholar] [CrossRef]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics Reveals a Remarkable Diversity of Lipids in Human Plasma1. J. Lipid Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [Green Version]
- Knapp, M.; Lisowska, A.; Zabielski, P.; Musiał, W.; Baranowski, M. Sustained Decrease in Plasma Sphingosine-1-Phosphate Concentration and Its Accumulation in Blood Cells in Acute Myocardial Infarction. Prostagland. Other Lipid Mediat. 2013, 106, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Chipeaux, C.; de Person, M.; Burguet, N.; Billette de Villemeur, T.; Rose, C.; Belmatoug, N.; Héron, S.; Le Van Kim, C.; Franco, M.; Moussa, F. Optimization of Ultra-High Pressure Liquid Chromatography—Tandem Mass Spectrometry Determination in Plasma and Red Blood Cells of Four Sphingolipids and Their Evaluation as Biomarker Candidates of Gaucher’s Disease. J. Chromatogr. A 2017, 1525, 116–125. [Google Scholar] [CrossRef]
- Dupuis, L.; Chipeaux, C.; Bourdelier, E.; Martino, S.; Reihani, N.; Belmatoug, N.; Billette de Villemeur, T.; Hivert, B.; Moussa, F.; Le Van Kim, C.; et al. Effects of Sphingolipids Overload on Red Blood Cell Properties in Gaucher Disease. J. Cell. Mol. Med. 2020, 24, 9726–9736. [Google Scholar] [CrossRef]
- Awojoodu, A.O.; Keegan, P.M.; Lane, A.R.; Zhang, Y.; Lynch, K.R.; Platt, M.O.; Botchwey, E.A. Acid Sphingomyelinase Is Activated in Sickle Cell Erythrocytes and Contributes to Inflammatory Microparticle Generation in SCD. Blood 2014, 124, 1941–1950. [Google Scholar] [CrossRef] [Green Version]
- Świderska, M.; Knapp, P.; Bodnar, L.; Błachnio-Zabielska, A.; Reszeć, J.; Chabowski, A. Blood Bioactive Sphingolipids in Patients with Advanced Serous Epithelial Ovarian Cancer-Mass Spectrometry Analysis Obstetrics and Gynecology. Arch. Med. Sci. 2021, 17, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Burrello, J.; Biemmi, V.; Dei Cas, M.; Amongero, M.; Bolis, S.; Lazzarini, E.; Bollini, S.; Vassalli, G.; Paroni, R.; Barile, L. Sphingolipid Composition of Circulating Extracellular Vesicles after Myocardial Ischemia. Sci. Rep. 2020, 10, 16182. [Google Scholar] [CrossRef] [PubMed]
- Santa, T. Isothiocyanates as Derivatization Reagents for Amines in Liquid Chromatography/Electrospray Ionization-Tandem Mass Spectrometry. Biomed. Chromatogr. 2010, 24, 915–918. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | CV% Intra-Day (n = 5) | CV% Inter-Days (n = 10) |
|---|---|---|
| Cer | 1.6 | 7.9 |
| dhCer | 3.8 | 7.6 |
| SM | 3.8 | 7.9 |
| HexCer | 7.2 | 6.0 |
| LacCer | 7.8 | 12.8 |
| GM3 | 7.6 | 14.0 |
| Sph | 4.2 (4.8) | 4.8 (13.1) |
| S1P | 5.0 (2.1) | 3.8 (12.4) |
| dhSph | 10.1 (11.1) | 11.2 (14.8) |
| dhS1P | 8.1 (2.4) | 6.8 (11.9) |
| Class | CV% Intra-Day (n = 5) | CV% Inter-Days (n = 10) |
|---|---|---|
| Cer | 7.4 | 10.5 |
| dhCer | 7.2 | 15.0 |
| SM | 8.4 | 9.3 |
| HexCer | 13.4 | 10.5 |
| LacCer | 12.4 | 9.8 |
| GM3 | 11.2 | 11.3 |
| Sph | 9.6 | 13.9 |
| S1P | 9.4 | 8.3 |
| dhSph | 9.9 | 8.0 |
| dhS1P | 7.7 | 10.0 |
| Conc (µM) | Min | Max | Mean ± SD (n = 20) | % |
|---|---|---|---|---|
| Cer | 8.1 | 14.6 | 11.2 ± 2.0 | 4.5 |
| dhCer | 0.2 | 0.8 | 0.54 ± 0.20 | 0.2 |
| SM | 181.7 | 248.9 | 217.9 ± 19.3 | 87.2 |
| HexCer | 3.4 | 10.7 | 7.6 ± 1.9 | 3.0 |
| LacCer | 2.9 | 8.5 | 5.5 ± 1.4 | 2.2 |
| GM3 | 2.6 | 5.7 | 4.7 ± 0.7 | 1.9 |
| Sph | 0.04 | 0.2 | 0.07 ± 0.034 | 0.03 |
| S1P | 0.9 | 3.3 | 2.0 ± 0.593 | 0.8 |
| dhSph | 0.003 | 0.03 | 0.01 ± 0.006 | 0.004 |
| dhS1P | 0.23 | 0.80 | 0.45 ± 0.177 | 0.2 |
| Conc (µM) | Min | Max | Mean ± SD (n = 20) | % |
|---|---|---|---|---|
| Cer | 59.9 | 148.5 | 92.8 ± 21.1 | 5.9 |
| dhCer | 9.6 | 39.3 | 22.7 ± 8.0 | 1.4 |
| SM | 854.6 | 2000 | 1387 ± 237.3 | 87.4 |
| HexCer | 3.3 | 8.5 | 6.7 ± 1.5 | 0.4 |
| LacCer | 21.6 | 115.9 | 64.2 ± 23.8 | 4.0 |
| GM3 | 1.5 | 4.6 | 3.3 ± 0.9 | 0.2 |
| Sph | 1.7 | 4.0 | 2.5 ± 0.64 | 0.1 |
| S1P | 2.6 | 6.4 | 4.1 ± 1.041 | 0.3 |
| dhSph | 0.1 | 0.4 | 0.2 ± 0.112 | 0.01 |
| dhS1P | 1.5 | 3.7 | 2.2 ± 0.563 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morano, C.; Zulueta, A.; Caretti, A.; Roda, G.; Paroni, R.; Dei Cas, M. An Update on Sphingolipidomics: Is Something Still Missing? Some Considerations on the Analysis of Complex Sphingolipids and Free-Sphingoid Bases in Plasma and Red Blood Cells. Metabolites 2022, 12, 450. https://doi.org/10.3390/metabo12050450
Morano C, Zulueta A, Caretti A, Roda G, Paroni R, Dei Cas M. An Update on Sphingolipidomics: Is Something Still Missing? Some Considerations on the Analysis of Complex Sphingolipids and Free-Sphingoid Bases in Plasma and Red Blood Cells. Metabolites. 2022; 12(5):450. https://doi.org/10.3390/metabo12050450
Chicago/Turabian StyleMorano, Camillo, Aida Zulueta, Anna Caretti, Gabriella Roda, Rita Paroni, and Michele Dei Cas. 2022. "An Update on Sphingolipidomics: Is Something Still Missing? Some Considerations on the Analysis of Complex Sphingolipids and Free-Sphingoid Bases in Plasma and Red Blood Cells" Metabolites 12, no. 5: 450. https://doi.org/10.3390/metabo12050450
APA StyleMorano, C., Zulueta, A., Caretti, A., Roda, G., Paroni, R., & Dei Cas, M. (2022). An Update on Sphingolipidomics: Is Something Still Missing? Some Considerations on the Analysis of Complex Sphingolipids and Free-Sphingoid Bases in Plasma and Red Blood Cells. Metabolites, 12(5), 450. https://doi.org/10.3390/metabo12050450