Comprehensive Metabolomic Comparison of Five Cereal Vinegars Using Non-Targeted and Chemical Isotope Labeling LC-MS Analysis

 , and
, and
Abstract
:
1. Introduction
2. Results
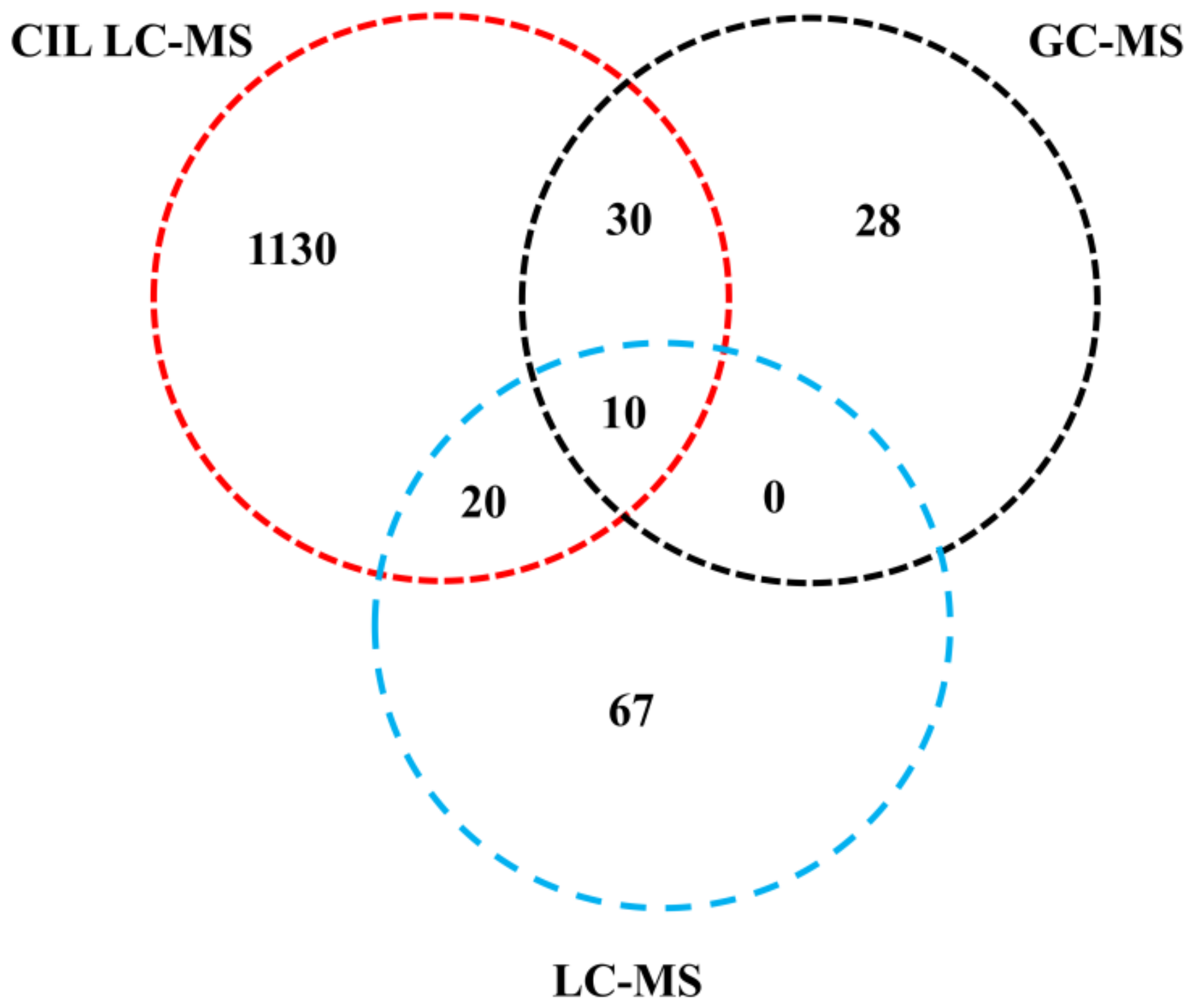
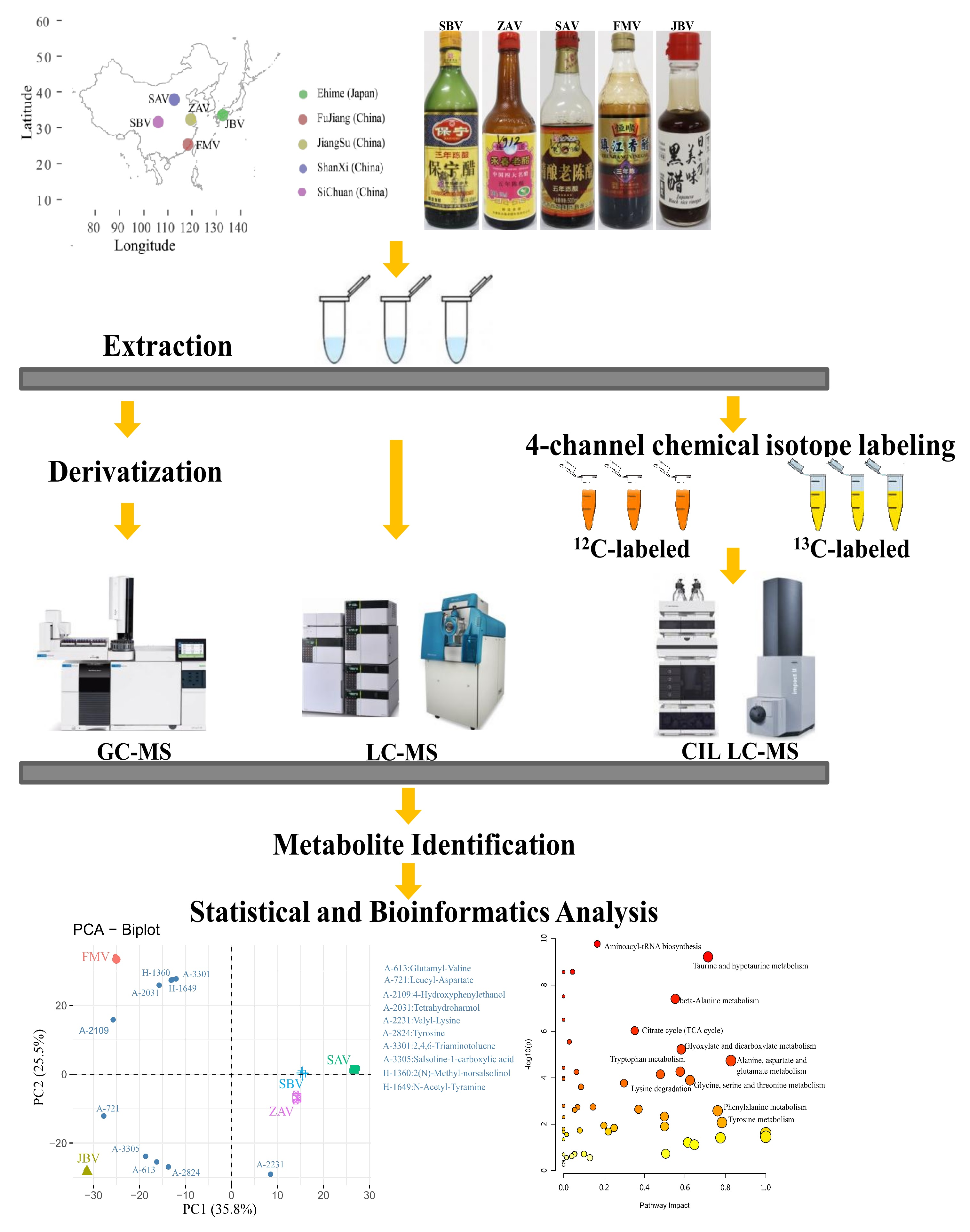
2.1. General Information of Identified Metabolites
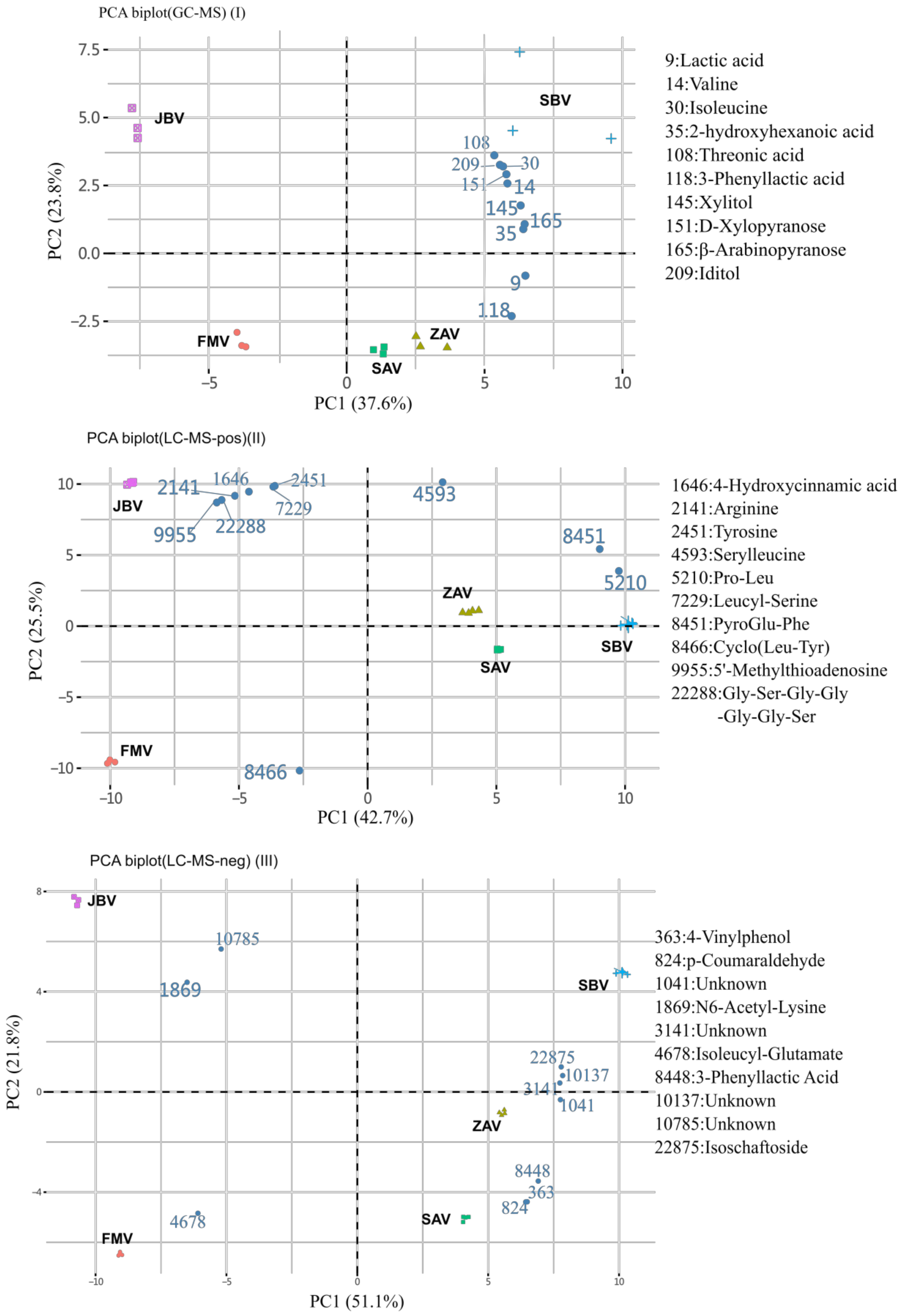
2.2. Non-Targeted-Based Analysis of Metabolites
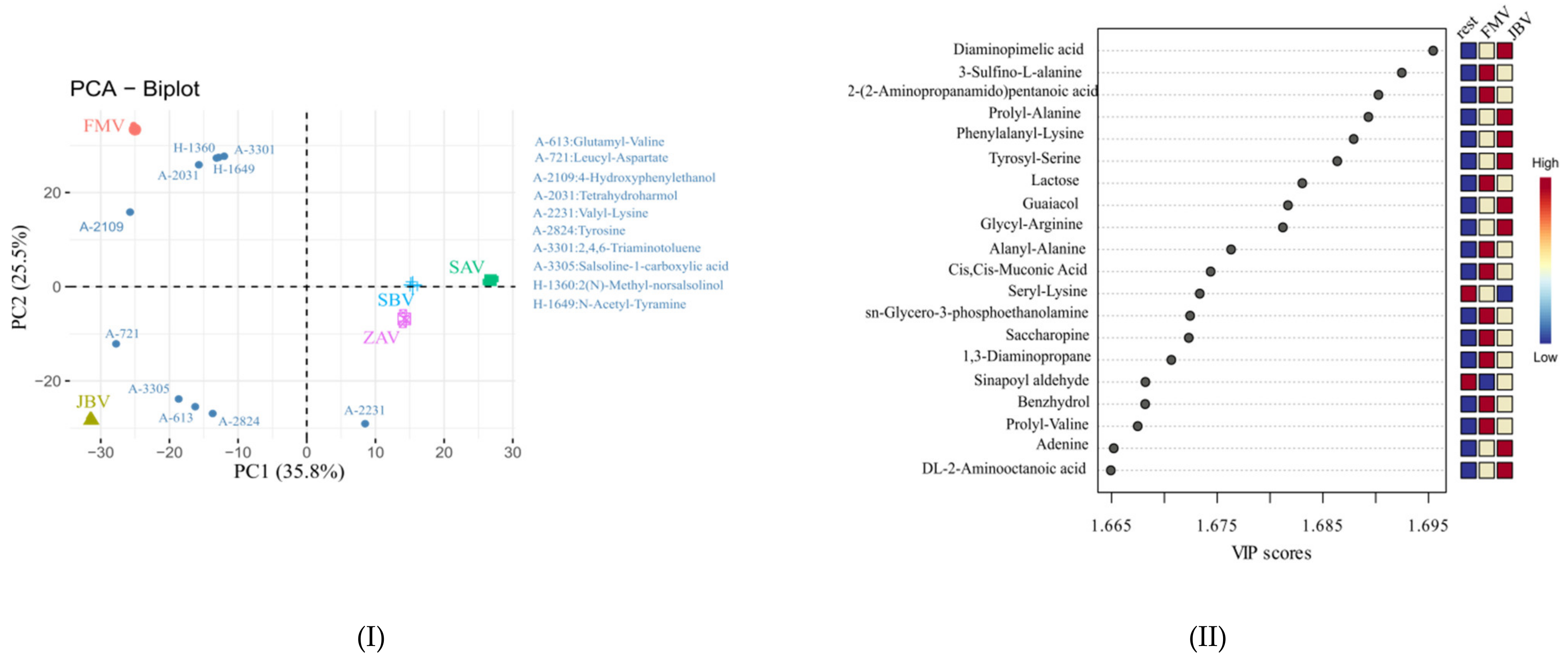
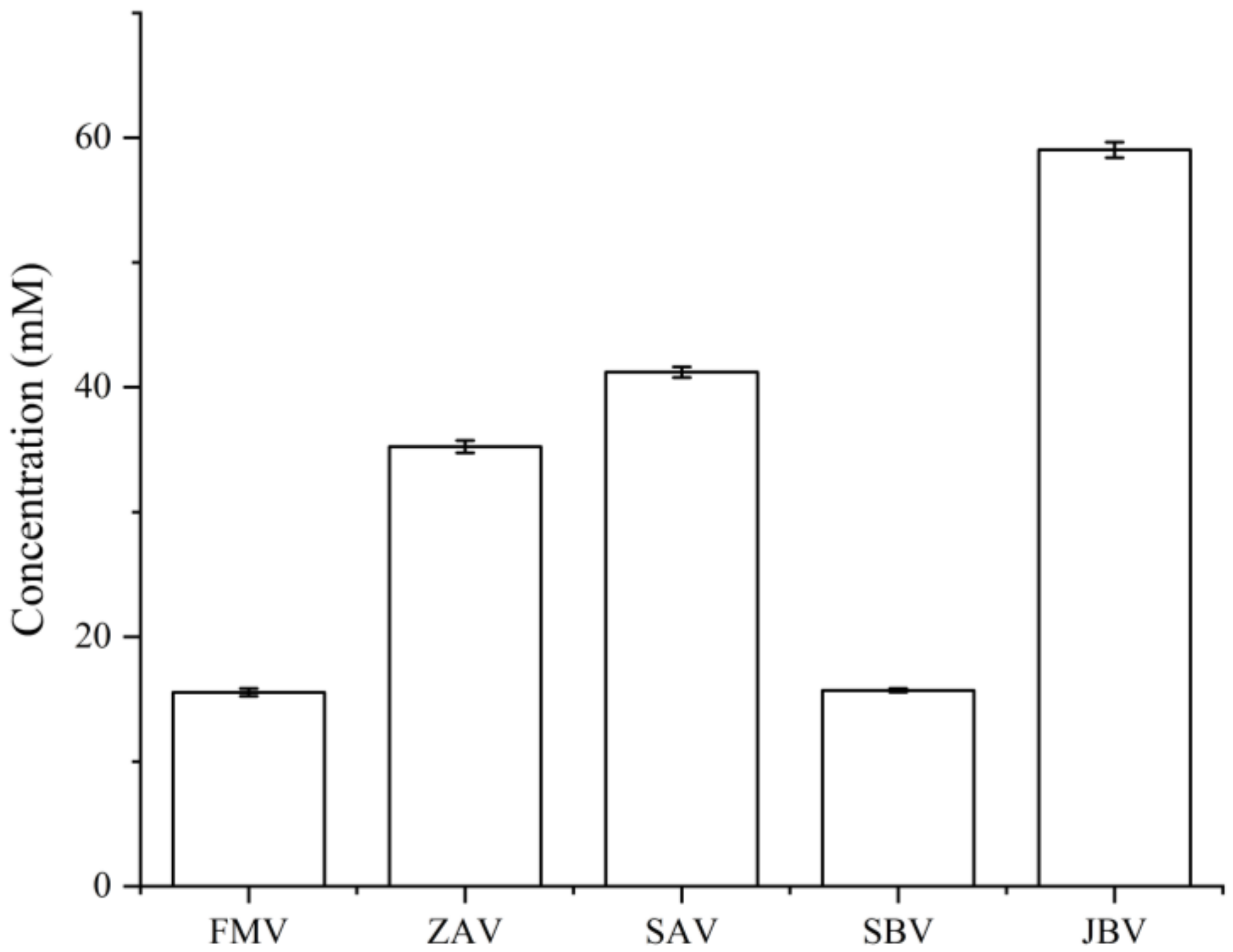
2.3. CIL LC-MS Metabolites and LC-UV Quantification
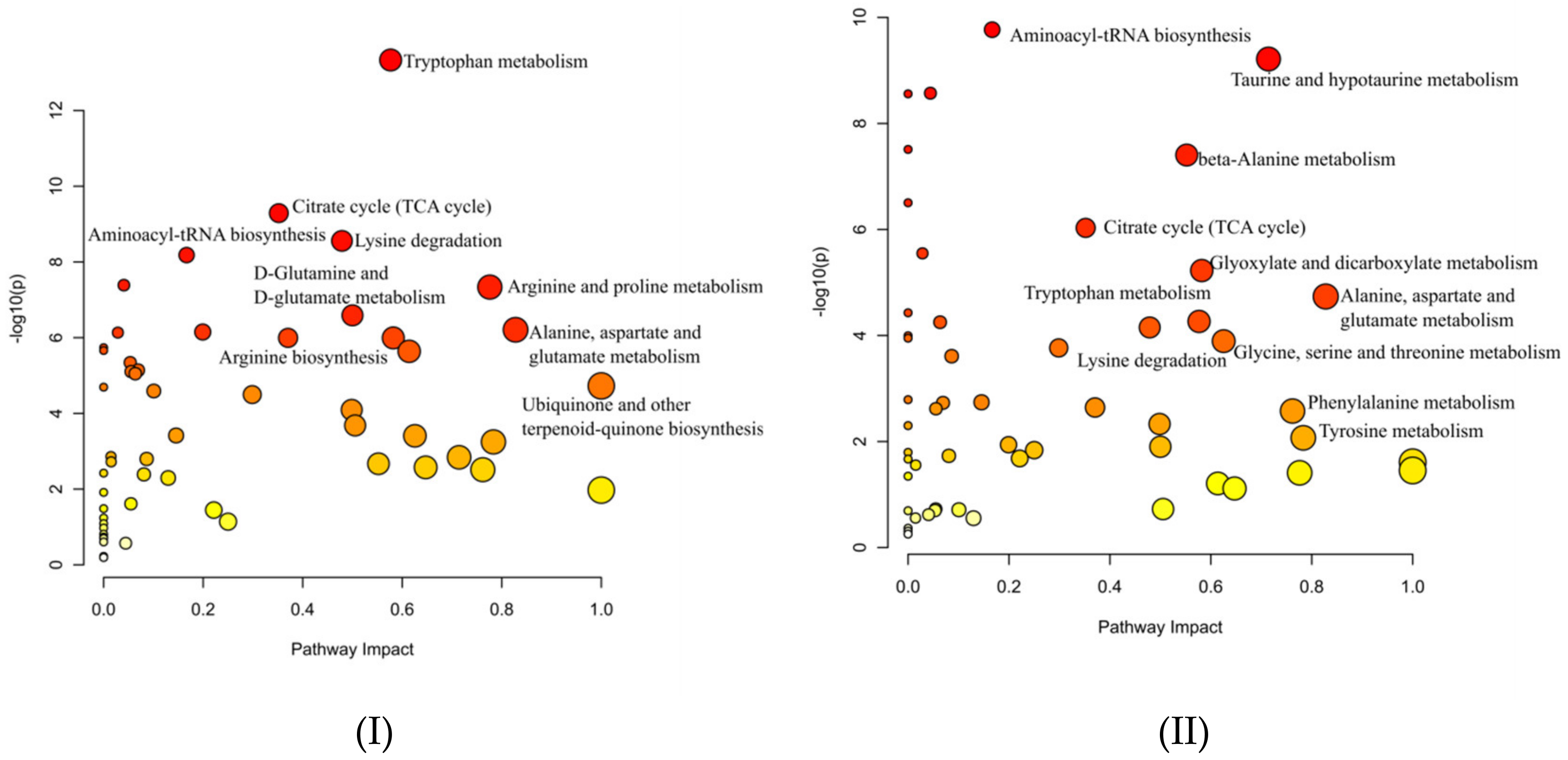
2.4. Pathway Annotation in Vinegar Metabolome
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. GC-MS Analysis
4.3. Conventional or Label-Free LC-MS Analysis
4.4. CIL LC-MS Analysis
4.5. Pathway Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.; He, Z.; Hao, W.; Zhu, H.; Liang, N.; Liu, J.; Zhang, C.; Ma, K.; He, W.; Yang, Y.; et al. Vinegars but not acetic acid are effective in reducing plasma cholesterol in hamsters fed a high-cholesterol diet. Food Funct. 2020, 11, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Zannini, E.; Wilkinson, S.; Daenen, L.; Arendt, E.K. Physiology of Acetic Acid Bacteria and Their Role in Vinegar and Fermented Beverages. Compr. Rev. Food Sci. Food Saf. 2019, 18, 587–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.M.; Wang, Z.M.; Zhang, X.J.; Mao, J.; Shi, J.S.; Xu, Z.H. Microbial ecology of cereal vinegar fermentation: Insights for driving the ecosystem function. Curr. Opin. Biotechnol. 2018, 49, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, T.; Giudici, P.; Chen, F. Vinegar Functions on Health: Constituents, Sources, and Formation Mechanisms. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1124–1138. [Google Scholar] [CrossRef]
- Ai, M.; Qiu, X.; Huang, J.; Wu, C.; Jin, Y.; Zhou, R. Characterizing the microbial diversity and major metabolites of Sichuan bran vinegar augmented by Monascus purpureus. Int. J. Food Microbiol. 2019, 292, 83–90. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, B.; Duan, W.; Zhang, J.; Wang, M. Nutrients and bioactive components from vinegar: A fermented and functional food. J. Funct. Foods 2020, 64, 103681. [Google Scholar] [CrossRef]
- Dai, W.; Qi, D.; Yang, T.; Lv, H.; Guo, L.; Zhang, Y. Non-targeted Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole-Time of Flight Mass Spectrometry Uncovers the Effects of Harvest Season on the Metabolites and Taste Quality in Tea (Camellia sinensis L.). J. Agric. Food Chem. 2015, 63, 9869–9878. [Google Scholar] [CrossRef]
- Xia, Y.; Yu, J.; Miao, W.; Shuang, Q. A UPLC-Q-TOF-MS-based metabolomics approach for the evaluation of fermented mare’s milk to koumiss. Food Chem. 2020, 320, 126619. [Google Scholar] [CrossRef]
- Zhao, S.; Li, L. Chemical derivatization in LC-MS-based metabolomics study. TrAC-Trends Anal. Chem. 2020, 131, 115988. [Google Scholar] [CrossRef]
- Zhao, S.; Li, H.; Han, W.; Chan, W.; Li, L. Metabolomic Coverage of Chemical-Group-Submetabolome Analysis: Group Classification and Four-Channel Chemical Isotope Labeling LC-MS. Anal. Chem. 2019, 91, 12108–12115. [Google Scholar] [CrossRef]
- del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernandez, M.; MMartin, M.C.; Alvarez, M.A. The biogenic amines putrescine and cadaverine show in vitro cytotoxicity at concentrations that can be found in foods. Sci. Rep. 2019, 9, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Martínez, N.; Martín, M.C.; Fernández, M.; Alvarez, M.A. qPCR for quantitative detection of tyramine-producing bacteria in dairy products. Food Res. Int. 2010, 43, 289–295. [Google Scholar] [CrossRef]
- Shi, C.; Liu, M.; Zhao, H.; Liang, L.; Zhang, B. Formation and Control of Biogenic Amines in Sufu-A Traditional Chinese Fermented Soybean Product: A Critical Review. Food Rev. Int. 2021, 1–22. [Google Scholar] [CrossRef]
- Otsuka, Y.; Shimamura, T.; Sakaji, M.; Arita, H.; Kashiwagi, T.; Ukeda, H. Quantification of proline-containing cyclic dipeptides by LC-MS/MS. Anal. Sci. 2020, 36, 977–980. [Google Scholar] [CrossRef] [Green Version]
- Mitsuhashi, S. Current topics in the biotechnological production of essential amino acids, functional amino acids, and dipeptides. Curr. Opin. Biotechnol. 2014, 26, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Gong, M.; Zhou, Z.; Yu, Y.; Liu, S.; Zhu, S.; Jian, D.; Cui, P.; Zhong, F.; Mao, J. Investigation of the 5-hydroxymethylfurfural and furfural content of Chinese traditional fermented vinegars from different regions and its correlation with the saccharide and amino acid content. LWT-Food Sci. Technol. 2020, 124, 109175. [Google Scholar] [CrossRef]
- Chaudhari, S.S.; Gokhale, D.V. Phenyllactic Acid: A Potential Antimicrobial Compound in Lactic acid Bacteria. J. Bacteriol. Mycol. 2016, 2, 121–125. [Google Scholar]
- Jung, S.; Hwang, H.; Lee, J.H. Effect of lactic acid bacteria on phenyllactic acid production in kimchi. Food Control 2019, 106, 106701. [Google Scholar] [CrossRef]
- Peters, A.; Krumbholz, P.; Jäger, E.; Heintz-Buschart, A.; Çakir, M.V.; Rothemund, S.; Gaudl, A.; Ceglarek, U.; Schöneberg, T.; Stäubert, C. Metabolites of lactic acid bacteria present in fermented foods are highly potent agonists of human hydroxycarboxylic acid receptor 3. PLoS Genet. 2019, 15, e1008145. [Google Scholar]
- Yang, C.; Hu, Z.; Lu, M.; Li, P.; Tan, J.; Chen, M.; Lv, H. Application of metabolomics pro fi ling in the analysis of metabolites and taste quality in di ff erent subtypes of white tea. Food Res. Int. 2018, 106, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Kaunietis, A.; Buivydas, A.; Čitavičius, D.J.; Kuipers, O.P. Heterologous biosynthesis and characterization of a glycocin from a thermophilic bacterium. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.; Kamble, D.; Lokhande, P. Iodine-catalyzed chemoselective dehydrogenation and aromatization of tetrahydro- b -carbolines: A short synthesis of Kumujian-C, Eudistomin-U. Tetrahedron Lett. 2018, 59, 2387–2392. [Google Scholar] [CrossRef]
- Kim, D.C.; Quang, T.H.; Yoon, C.S.; Ngan NT, T.; Lim SIl Lee, S.Y.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory activities of indole alkaloids from kanjang (Korean fermented soy source) in lipopolysaccharide-induced BV2 microglial cells. Food Chem. 2016, 213, 69–75. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen Rearrangement Rules: Computational MS/MS Fragmentation and Structure Elucidation Using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Mung, D.; Li, L. Applying quantitative metabolomics based on chemical isotope labeling LC-MS for detecting potential milk adulterant in human milk. Anal. Chim. Acta 2018, 1001, 78–85. [Google Scholar] [CrossRef]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 1–7. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | PubChem | Relative Ion Intensity in Different Types of Vinegar (×106) | Method b | ||||
|---|---|---|---|---|---|---|---|
| FMV | ZAV | SAV | SBV | JBV | |||
| Benzene and Substituted Derivatives (2) c | |||||||
| Tyramine | 5610 | 3.72 ± 0.17 | 0.71 ± 0.05 | 0.7 ± 0.07 | 0.73 ± 0.42 | 0.14 ± 0.02 | G, LP |
| 4-Vinylphenol | 62,453 | 1.27 ± 0.01 | 1.83 ± 0.08 | 1.84 ± 0.03 | 1.72 ± 0.05 | 0.16 ± 0 | LP, LN |
| Carboxylic acids and derivatives (14) | |||||||
| Pyroglutamic acid | 7405 | 4.81 ± 0.23 | 18.04 ± 1.56 | 15.01 ± 0.26 | 18.12 ± 1.36 | 4.29 ± 0.56 | G, LN, LP |
| Glycine | 750 | 6.2 ± 0.28 | 7.93 ± 0.4 | 4.92 ± 0.32 | 3.35 ± 1.23 | 2.82 ± 0.19 | G |
| Leucine | 6106 | 5.47 ± 0.11 | 7.86 ± 1.42 | 4.48 ± 0.13 | 6.55 ± 0.99 | 2.4 ± 0.12 | G |
| Succinic acid | 1110 | 4.59 ± 0.22 | 7.88 ± 0.27 | 3.76 ± 0.22 | 5.75 ± 1.29 | 2.05 ± 0.11 | G, LN |
| Valine | 6287 | 0.2 ± 0.06 | 2.66 ± 0.44 | 1.17 ± 0.21 | 5.52 ± 0.82 | 0.55 ± 0.06 | G |
| Alanine | 5950 | 8.99 ± 0.54 | 7.28 ± 1.2 | 6.88 ± 0.44 | 8.61 ± 1.25 | 1.12 ± 0.16 | G |
| Citric acid | 311 | 0.12 ± 0.02 | 2.06 ± 0.25 | 3.48 ± 0.01 | 0.14 ± 0.01 | 0.67 ± 0.05 | G, LN |
| 4-Aminobutyric acid(GABA) | 119 | 3.88 ± 0.32 | 2.44 ± 0.21 | 4.64 ± 0.25 | 1.07 ± 0.63 | 0.39 ± 0.02 | G |
| Phenylalanine | 6140 | 0.29 ± 0.01 | 0.38 ± 0.01 | 0.27 ± 0.01 | 0.4 ± 0.01 | 0.52 ± 0.01 | LP, G, LN |
| Tyrosine | 6057 | 0.87 ± 0.01 | 2.76 ± 0.1 | 2.41 ± 0.12 | 3.46 ± 0.12 | 0.61 ± 0.02 | LP, G |
| N-(1-Deoxy-1-fructosyl)phenylalanine | 101,039,148 | 2.99 ± 0.18 | 10.35 ± 0.27 | 0.95 ± 0.06 | 26.55 ± 1.19 | 25.14 ± 0.92 | LP, LN |
| Isoleucine | 6306 | 1.1 ± 0.04 | 1.74 ± 0.06 | 1.34 ± 0.06 | 1.42 ± 0.07 | 1.53 ± 0.04 | LP, LN, G |
| Cyclo(Pro-Leu) | 102,892 | 0.65 ± 0.03 | 1.15 ± 0.02 | 7.23 ± 0.19 | 1.06 ± 0.05 | 0.11 ± 0.01 | LP |
| Cyclo(Phe-Pro) | 99,895 | 8.08 ± 0.46 | 30.36 ± 0.75 | 48.52 ± 1.12 | 44.23 ± 2.11 | 8.9 ± 0.29 | LP |
| Hydroxy acids and derivatives (1) | |||||||
| Lactic acid | 107,689 | 33.05 ± 0.89 | 63.18 ± 0.61 | 51.33 ± 1.41 | 75.58 ± 0.5 | 5.68 ± 0.23 | G, LN |
| Phenylpropanoic acids (2) | |||||||
| 3-Phenyllactic acid | 3848 | 27.02 ± 1.05 | 33.97 ± 1.56 | 33.58 ± 1.39 | 32.9 ± 0.62 | 3.35 ± 0.03 | LN, G |
| Hydroxyphenyllactic acid | 9378 | 16.49 ± 0.88 | 9.41 ± 0.14 | 4.99 ± 0.11 | 4.19 ± 0.1 | 0.51 ± 0 | LN |
| Fatty Acyls (2) | |||||||
| isohexonic acid | 12,344 | 0.68 ± 0.06 | 0.85 ± 0.07 | 0.3 ± 0.04 | 0.56 ± 0.05 | 4.5 ± 0.74 | G |
| 9,10,13-Trihydroxystearic acid | 45,359,277 | 1.65 ± 0.05 | 14.97 ± 0.24 | 4.39 ± 2.57 | 39.55 ± 0.93 | 0 ± 0 | LP, LN |
| Harmala alkaloids (2) | |||||||
| Tetrahydroharman-3-carboxylic acid | 73,530 | 1.03 ± 0.07 | 4.29 ± 0.08 | 2.32 ± 0.03 | 0.83 ± 0.05 | 4.55 ± 0.23 | LP, LN |
| Harmalan | 160,510 | 25.51 ± 0.53 | 0.19 ± 0.01 | 3.32 ± 0.15 | 11.97 ± 0.31 | 0.21 ± 0.02 | LP |
| Organooxygen compounds (14) | |||||||
| Glucose | 5793 | 44.94 ± 2.4 | 46.47 ± 0.97 | 6.57 ± 0.83 | 35.19 ± 23.77 | 61.87 ± 1.51 | G |
| Glycerol | 753 | 10.08 ± 0.35 | 25.73 ± 1.13 | 16.79 ± 0.61 | 28.82 ± 2.64 | 20.75 ± 0.54 | G |
| 2,3-Butanediol | 262 | 4.63 ± 0.19 | 8 ± 0.16 | 6.73 ± 0.19 | 5.91 ± 0.78 | 4.74 ± 0.15 | G |
| Fructose | 439,163 | 37.55 ± 3.58 | 27.07 ± 3.12 | 1.35 ± 0.08 | 45.83 ± 3.17 | 4.34 ± 0.4 | G |
| Inositol | 892 | 0.96 ± 0.22 | 16.33 ± 1.59 | 13.32 ± 1.77 | 11.16 ± 1.43 | 3.46 ± 0.31 | G |
| Ribofuranose | 5779 | 0.17 ± 0.02 | 11.72 ± 0.62 | 9.49 ± 1.43 | 5.52 ± 0.92 | 2.06 ± 0.15 | G |
| Iditol | 5,460,044 | 0.46 ± 0.4 | 5.38 ± 0.65 | 1.19 ± 0.07 | 10.67 ± 1.01 | 0.78 ± 0.3 | G |
| Xylopyranose | 135,191 | 0.03 ± 0 | 6.91 ± 0.78 | 10.98 ± 1.06 | 24.04 ± 2.77 | 0.58 ± 0.03 | G |
| Mannitol | 6251 | 0.47 ± 0.25 | 13.24 ± 1.38 | 4.71 ± 0.48 | 2.98 ± 0.29 | 0.39 ± 0.05 | G |
| Xylitol | 6912 | 0.07 ± 0.01 | 4.72 ± 0.12 | 4.27 ± 0.38 | 9.13 ± 0.86 | 0.24 ± 0.03 | G |
| Threitol | 222,285 | 2.66 ± 0.12 | 1.9 ± 0.16 | 5.23 ± 0.56 | 10.68 ± 0.9 | 0.14 ± 0.02 | G |
| Glyceric acid | 439,194 | 0.28 ± 0.01 | 2.14 ± 0.14 | 4.11 ± 0.28 | 1.65 ± 0.33 | 0.13 ± 0.01 | G |
| Lyxose | 439,240 | 0.01 ± 0 | 1.71 ± 0.31 | 2.28 ± 0.17 | 6.17 ± 0.89 | 0.09 ± 0.01 | G |
| 1,3-Propanediol | 347,971 | 6.64 ± 0.27 | 0.11 ± 0.02 | 0.69 ± 0.01 | 0.91 ± 0.11 | 0.02 ± 0 | G |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhao, C.; Dong, L.; Huan, Y.; Yoshimoto, M.; Zhu, Y.; Tada, I.; Wang, X.; Zhao, S.; Zhang, F.; et al. Comprehensive Metabolomic Comparison of Five Cereal Vinegars Using Non-Targeted and Chemical Isotope Labeling LC-MS Analysis. Metabolites 2022, 12, 427. https://doi.org/10.3390/metabo12050427
Li Z, Zhao C, Dong L, Huan Y, Yoshimoto M, Zhu Y, Tada I, Wang X, Zhao S, Zhang F, et al. Comprehensive Metabolomic Comparison of Five Cereal Vinegars Using Non-Targeted and Chemical Isotope Labeling LC-MS Analysis. Metabolites. 2022; 12(5):427. https://doi.org/10.3390/metabo12050427
Chicago/Turabian StyleLi, Zhihua, Chi Zhao, Ling Dong, Yu Huan, Miwa Yoshimoto, Yongqing Zhu, Ipputa Tada, Xiaohang Wang, Shuang Zhao, Fengju Zhang, and et al. 2022. "Comprehensive Metabolomic Comparison of Five Cereal Vinegars Using Non-Targeted and Chemical Isotope Labeling LC-MS Analysis" Metabolites 12, no. 5: 427. https://doi.org/10.3390/metabo12050427
APA StyleLi, Z., Zhao, C., Dong, L., Huan, Y., Yoshimoto, M., Zhu, Y., Tada, I., Wang, X., Zhao, S., Zhang, F., Li, L., & Arita, M. (2022). Comprehensive Metabolomic Comparison of Five Cereal Vinegars Using Non-Targeted and Chemical Isotope Labeling LC-MS Analysis. Metabolites, 12(5), 427. https://doi.org/10.3390/metabo12050427