
Specific Changes in Arabidopsis thaliana Rosette Lipids during Freezing Can Be Associated with Freezing Tolerance

 , , , , , , , , , , , and
, , , , , , , , , , , and 
Abstract
1. Introduction
2. Results and Discussion
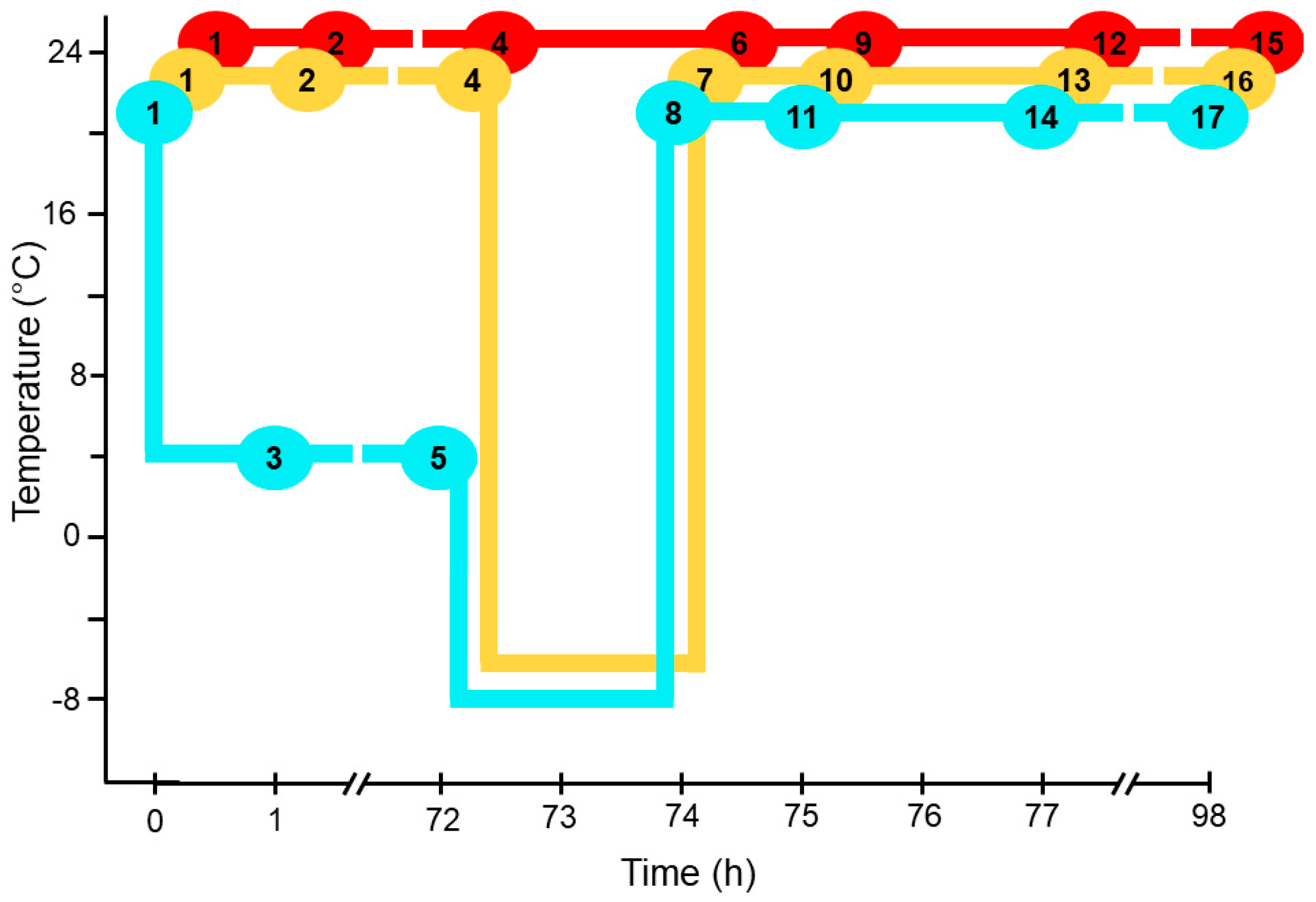
2.1. Experimental Design
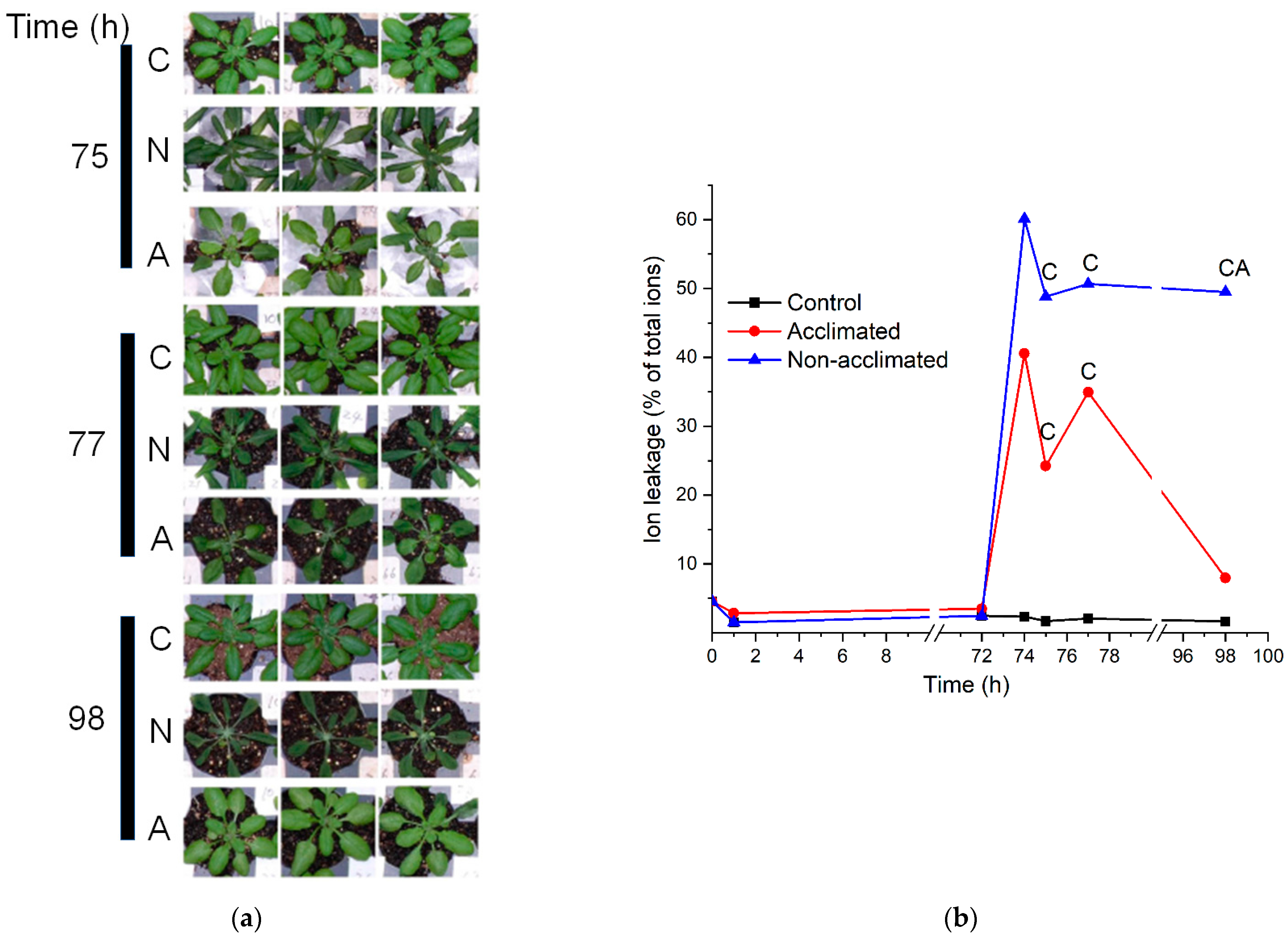
2.2. Plant Response to Treatment
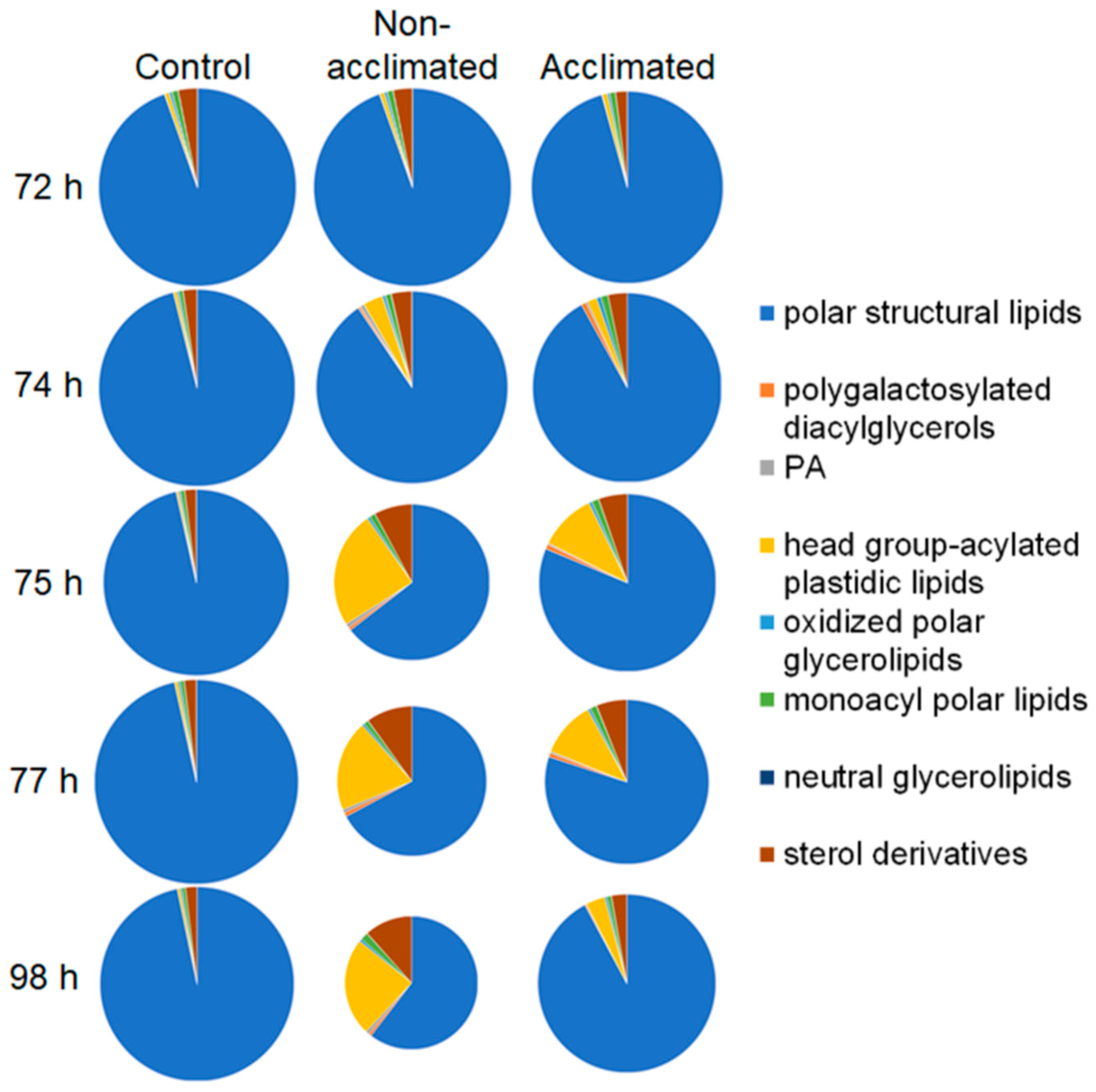
2.3. Overview of Lipid Changes in Wild-Type Plants
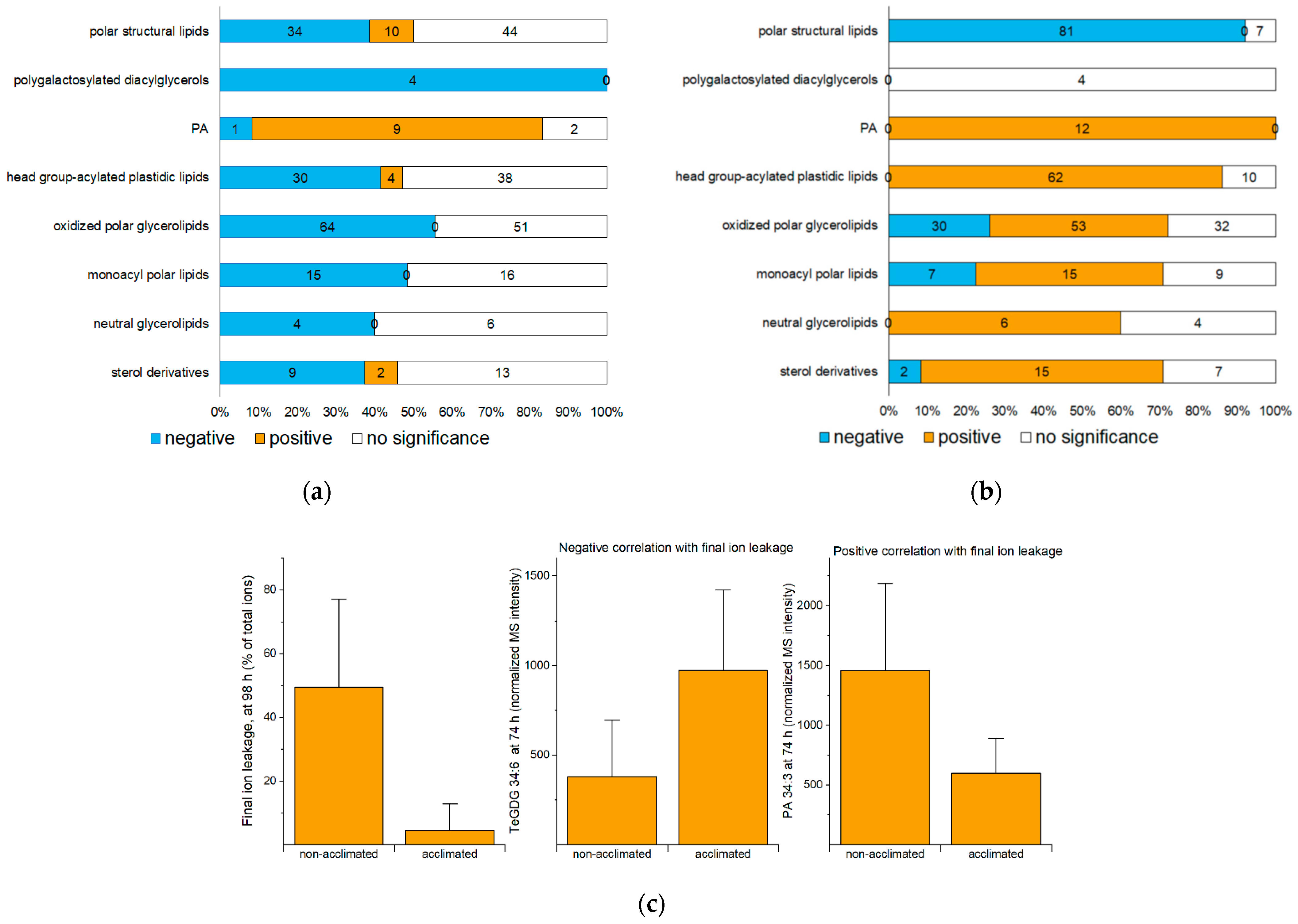
2.4. Correlation Analysis as a Predictor of the Role of Lipids in Freezing Response
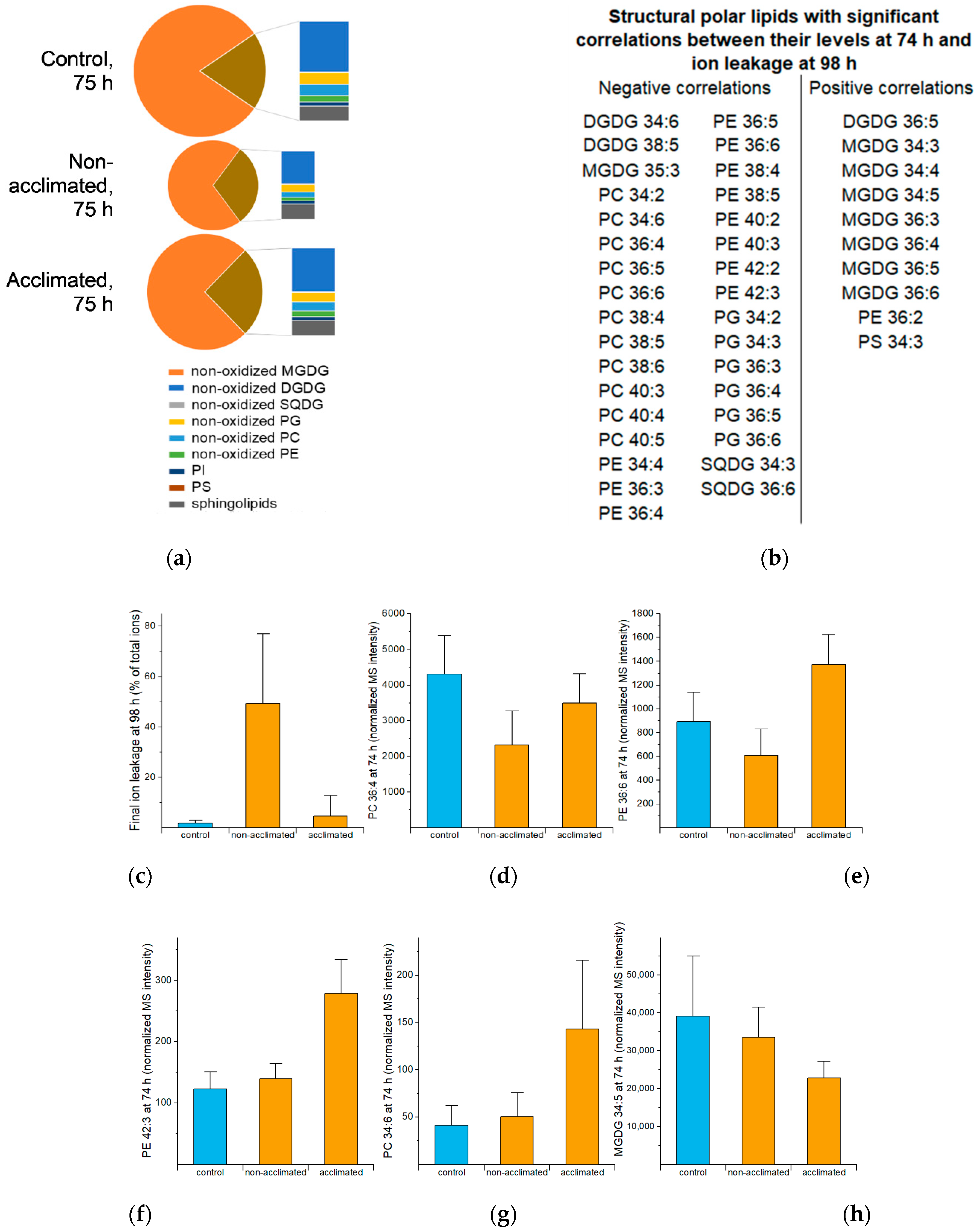
2.5. Structural Polar Lipids
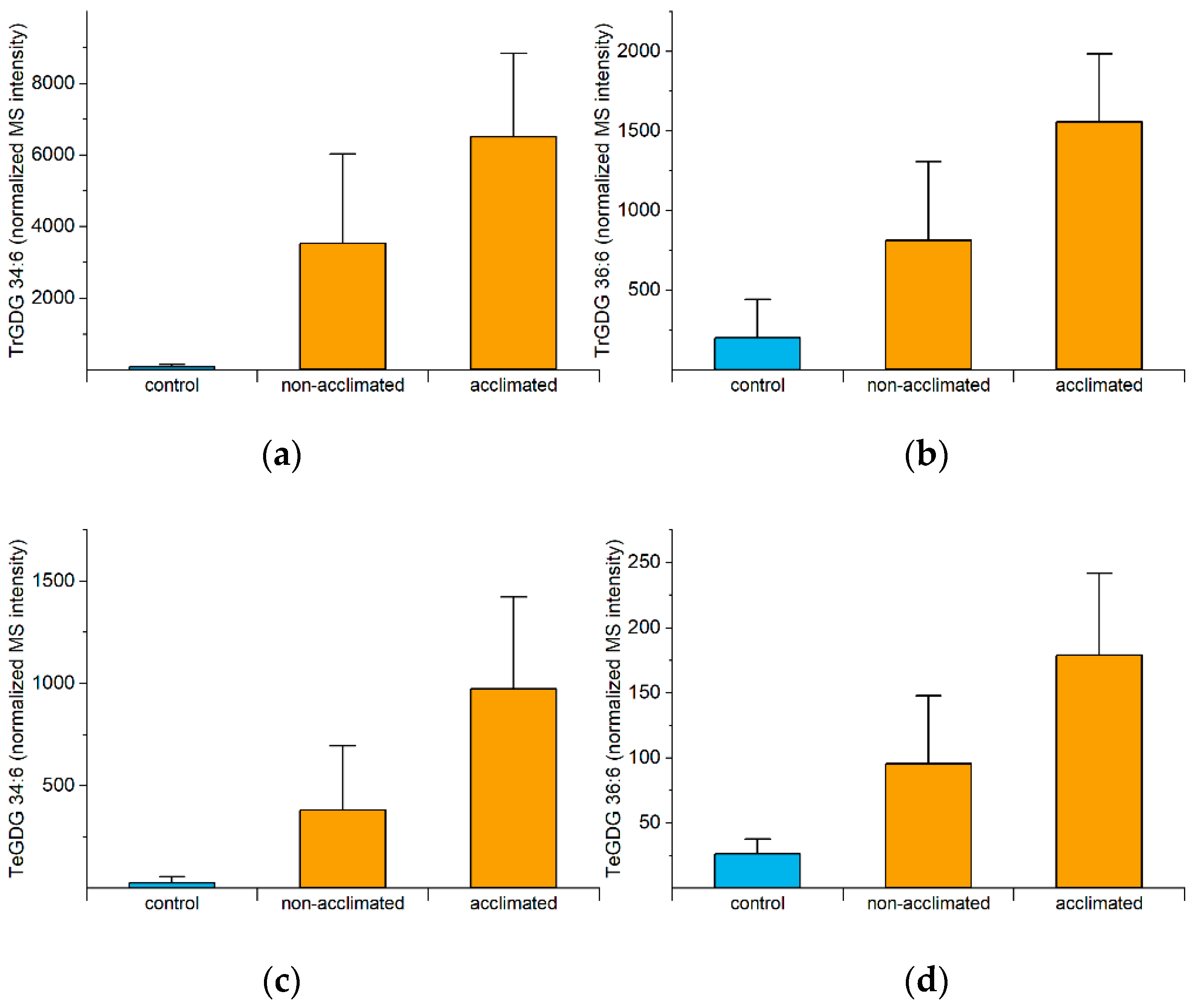
2.6. Polygalactosylated Diacylglycerols
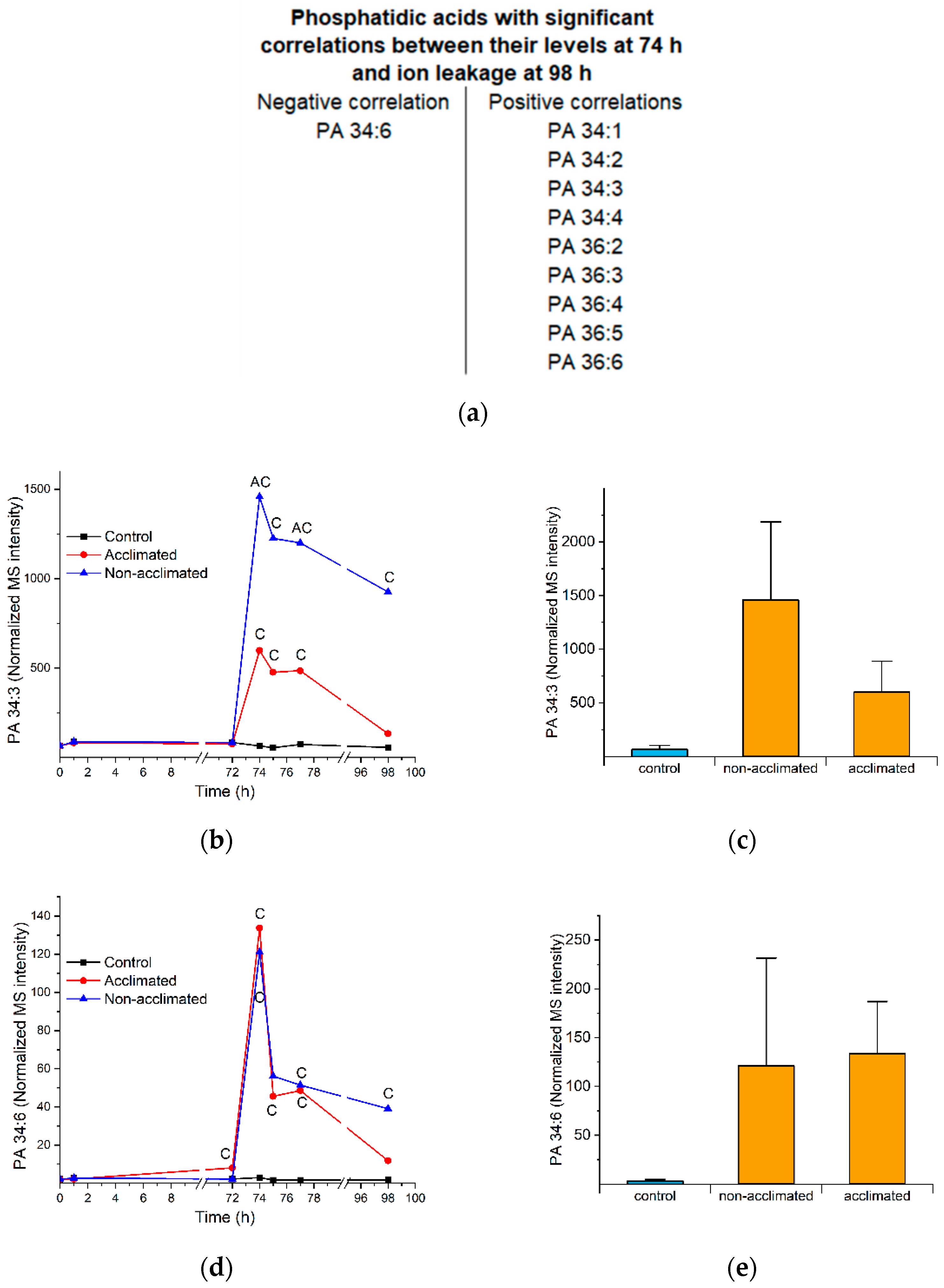
2.7. Phosphatidic Acid
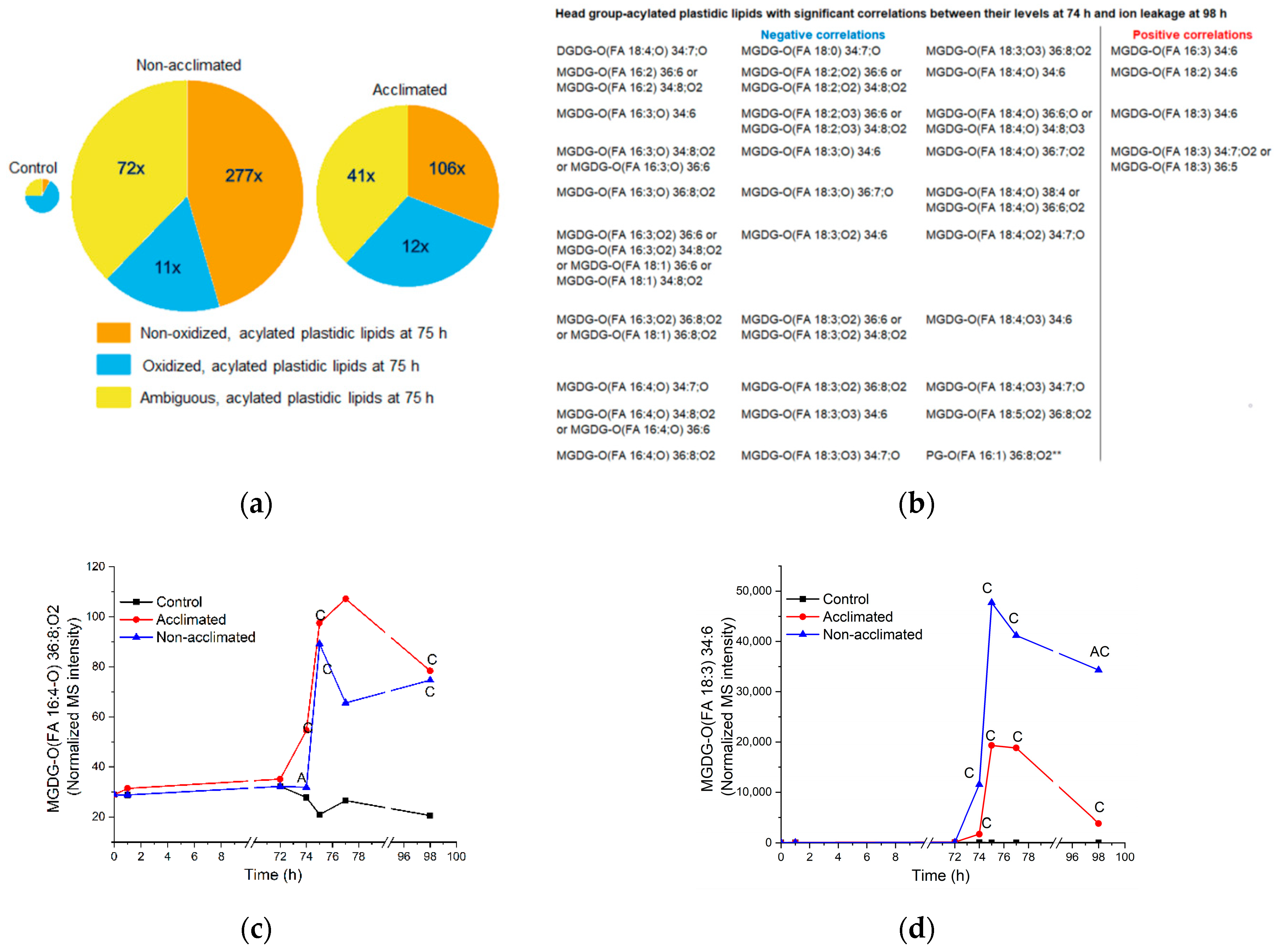
2.8. Head Group-Acylated Plastidic Lipids
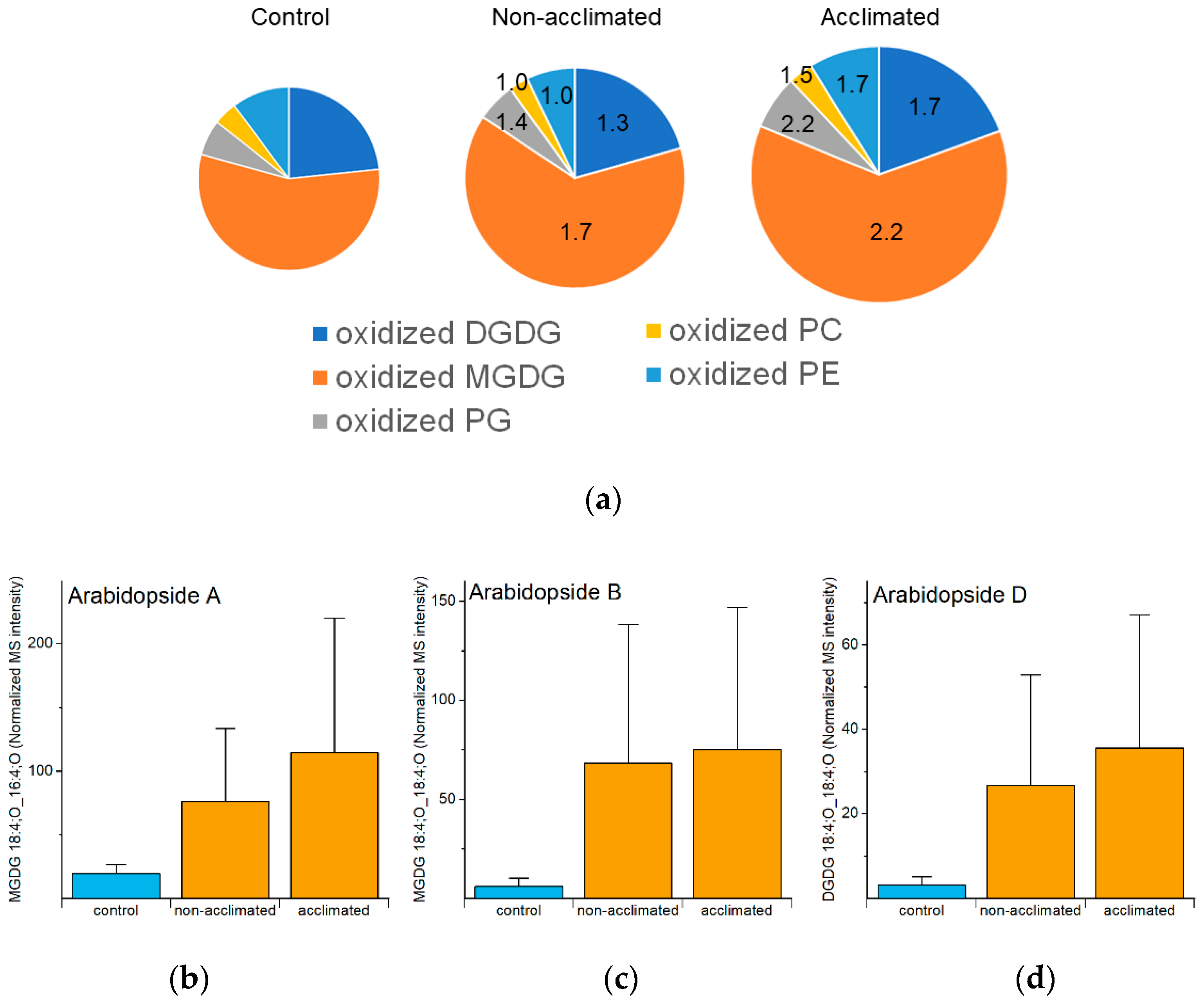
2.9. Oxidized Polar Diacylglycerolipids
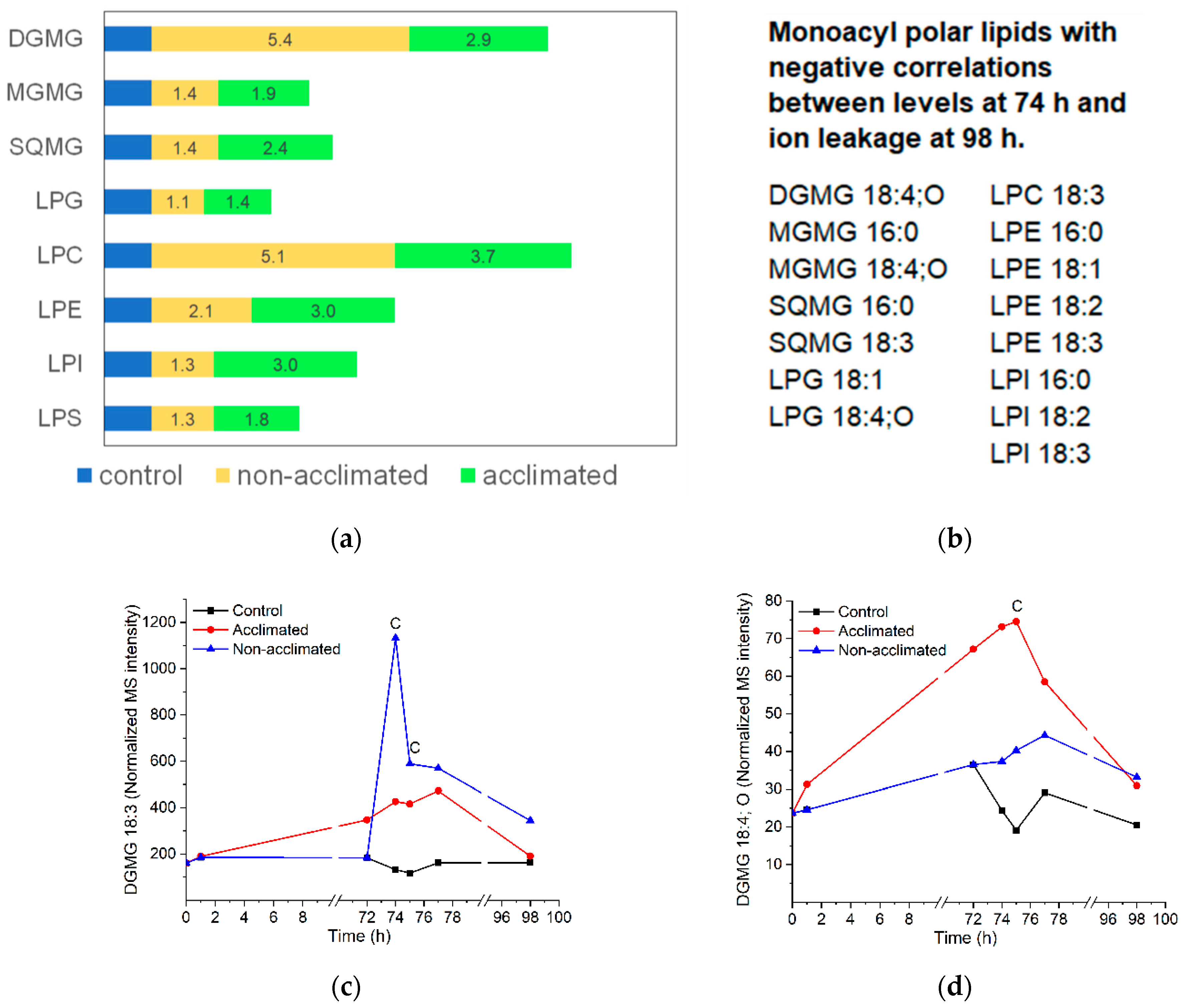
2.10. Monoacyl Polar Lipids
2.11. Neutral Glycerolipids
2.12. Sterol Derivatives
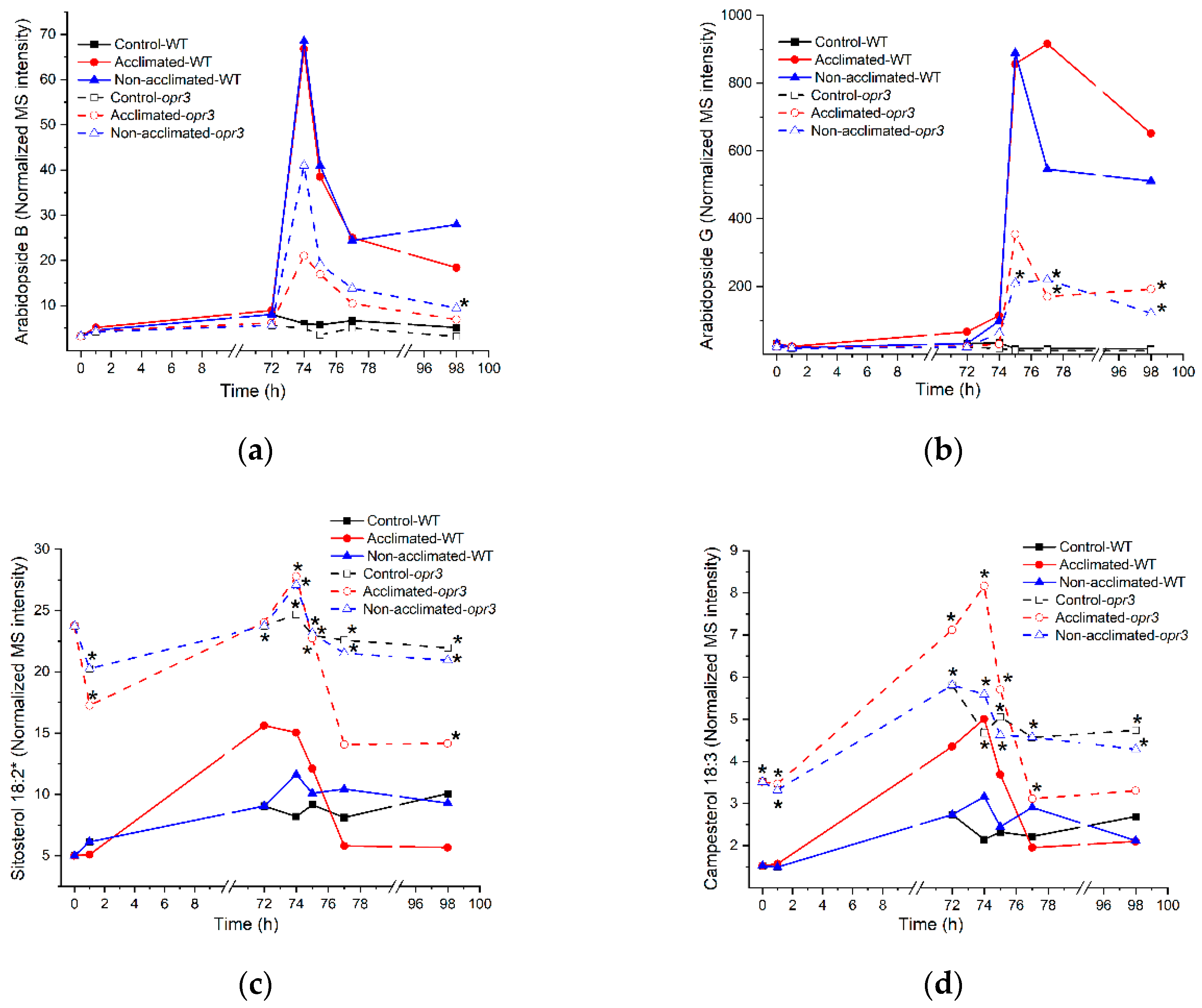
2.13. Lipids of Plants with Mutations in Lipid-Related Genes
2.14. Differences in Sizes of Mutant Plants Compared to Wild-Type Plants
2.15. Summary of Hypotheses Generated from Correlation Analsysis
3. Materials and Methods
3.1. Experimental Design
3.2. Plant Growth
3.3. Cold Acclimation and Freezing Treatment
3.4. Sampling, Lipid Extraction, and Profiling by ESI Triple Quadrupole Mass Spectrometry
3.5. Mass Spectrometry Data Collection and Processing
3.6. Phenotype Analysis
3.7. Statistical Analysis
3.7.1. Comparison of Lipid Levels and Ion Leakage of Wild-Type Plants at Each Time Point as a Function of Treatment
3.7.2. Correlation Analysis of Lipid Levels and Ion Leakage in Wild-Type Plants
3.7.3. Comparison of Lipid Levels and Ion Leakage at Each Time Point as a Function of Genotype (Wild Type vs. Mutant)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steponkus, P.L.; Lynch, D.V. Freeze/thaw-induced destabilization of the plasma membrane and the effects of cold acclimation. J. Bioenerg. Biomembr. 1989, 21, 21–41. [Google Scholar] [CrossRef] [PubMed]
- Hugly, S.; Somerville, C. A role for membrane lipid polyunsaturation in chloroplast biogenesis at low temperature. Plant Physiol. 1992, 99, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Miquel, M.; James, D., Jr.; Dooner, H.; Browse, J. Arabidopsis requires polyunsaturated lipids for low-temperature survival. Proc. Natl. Acad. Sci. USA 1993, 90, 6208–6212. [Google Scholar] [CrossRef] [PubMed]
- Welti, R.; Li, W.; Li, M.; Sang, Y.; Biesiada, H.; Zhou, H.E.; Rajashekar, C.B.; Williams, T.D.; Wang, X. Profiling membrane lipids in plant stress responses. Role of phospholipase Dα in freezing-induced lipid changes in Arabidopsis. J. Biol. Chem. 2002, 277, 31994–32002. [Google Scholar] [CrossRef] [PubMed]
- Moellering, E.R.; Muthan, B.; Benning, C. Freezing tolerance in plants requires lipid remodeling at the outer chloroplast membrane. Science 2010, 330, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.S.; Tamura, P.; Galeva, N.A.; Chaturvedi, R.; Roth, M.R.; Williams, T.D.; Wang, X.; Shah, J.; Welti, R. Direct infusion mass spectrometry of oxylipin-containing Arabidopsis membrane lipids reveals varied patterns in different stress responses. Plant Physiol. 2012, 158, 324–339. [Google Scholar] [CrossRef]
- Vu, H.S.; Roth, M.R.; Tamura, P.; Samarakoon, T.; Shiva, S.; Honey, S.; Lowe, K.; Schmelz, E.A.; Williams, T.D.; Welti, R. Head-group acylation of monogalactosyldiacylglycerol is a common stress response, and the acyl-galactose acyl composition varies with the plant species and applied stress. Physiol. Plant. 2014, 150, 517–528. [Google Scholar] [CrossRef]
- Tarazona, P.; Feussner, K.; Feussner, I. An enhanced plant lipidomics method based on multiplexed liquid chromatography-mass spectrometry reveals additional insights into cold- and drought-induced membrane remodeling. Plant J. 2015, 84, 621–633. [Google Scholar] [CrossRef]
- Vijayan, P.; Browse, J. Photoinhibition in mutants of Arabidopsis deficient in thylakoid unsaturation. Plant Physiol. 2002, 129, 876–885. [Google Scholar] [CrossRef]
- Thorlby, G.; Fourrier, N.; Warren, G. The SENSITIVE TO FREEZING2 gene, required for freezing tolerance in Arabidopsis thaliana, encodes a beta-glucosidase. Plant Cell 2004, 16, 2192–2203. [Google Scholar] [CrossRef]
- Tan, W.J.; Yang, Y.C.; Zhou, Y.; Huang, L.P.; Xu, L.; Chen, Q.F.; Yu, L.J.; Xiao, S. DIACYLGLYCEROL ACYLTRANSFERASE and DIACYLGLYCEROL KINASE modulate triacylglycerol and phosphatidic acid production in the plant response to freezing stress. Plant Physiol. 2018, 177, 1303–1318, Erratum in Plant Physiol. 2018, 178, 1424–1425. [Google Scholar] [CrossRef] [PubMed]
- Arisz, S.A.; Heo, J.Y.; Koevoets, I.T.; Zhao, T.; van Egmond, P.; Meyer, A.J.; Zeng, W.; Niu, X.; Wang, B.; Mitchell-Olds, T.; et al. DIACYLGLYCEROL ACYLTRANSFERASE1 contributes to freezing tolerance. Plant Physiol. 2018, 177, 1410–1424. [Google Scholar] [CrossRef] [PubMed]
- Arisz, S.A.; van Wijk, R.; Roels, W.; Zhu, J.K.; Haring, M.A.; Munnik, T. Rapid phosphatidic acid accumulation in response to low temperature stress in Arabidopsis is generated through diacylglycerol kinase. Front. Plant Sci. 2013, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.S.; Shiva, S.; Roth, M.R.; Tamura, P.; Zheng, L.; Li, M.; Sarowar, S.; Honey, S.; McEllhiney, D.; Hinkes, P.; et al. Lipid changes after leaf wounding in Arabidopsis thaliana: Expanded lipidomic data form the basis for lipid co-occurrence analysis. Plant J. 2014, 80, 728–743. [Google Scholar] [CrossRef] [PubMed]
- Shiva, S.; Samarakoon, T.; Lowe, K.A.; Roach, C.; Vu, H.S.; Colter, M.; Porras, H.; Hwang, C.; Roth, M.R.; Tamura, P.; et al. Leaf lipid alterations in response to heat stress of Arabidopsis thaliana. Plants 2020, 9, 845. [Google Scholar] [CrossRef] [PubMed]
- Liebisch, G.; Fahy, E.; Aoki, J.; Dennis, E.A.; Durand, T.; Ejsing, C.S.; Fedorova, M.; Feussner, I.; Griffiths, W.J.; Köfeler, H.; et al. Update on LIPID MAPS classification, nomenclature, and shorthand notation for MS-derived lipid structures. J. Lipid Res. 2020, 61, 1539–1555. [Google Scholar] [CrossRef]
- Degenkolbe, T.; Giavalisco, P.; Zuther, E.; Seiwert, B.; Hincha, D.K.; Willmitzer, L. Differential remodeling of the lipidome during cold acclimation in natural accessions of Arabidopsis thaliana. Plant J. 2012, 72, 972–982. [Google Scholar] [CrossRef]
- Zuther, E.; Schulz, E.; Childs, L.H.; Hincha, D.K. Clinal variation in the non-acclimated and cold-acclimated freezing tolerance of Arabidopsis thaliana accessions. Plant Cell Environ. 2012, 35, 1860–1878. [Google Scholar] [CrossRef]
- Devaiah, S.P.; Roth, M.R.; Baughman, E.; Li, M.; Tamura, P.; Jeannotte, R.; Welti, R.; Wang, X. Quantitative profiling of polar glycerolipid species from organs of wild-type Arabidopsis and a phospholipase Dα1 knockout mutant. Phytochemistry 2006, 67, 1907–1924. [Google Scholar] [CrossRef]
- Chen, L.; Hu, W.; Mishra, N.; Wei, J.; Lu, H.; Hou, Y.; Qiu, X.; Yu, S.; Wang, C.; Zhang, H.; et al. AKR2A interacts with KCS1 to improve VLCFAs contents and chilling tolerance of Arabidopsis thaliana. Plant J. 2020, 103, 1575–1589. [Google Scholar] [CrossRef]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar] [PubMed]
- Li, W.; Li, M.; Zhang, W.; Welti, R.; Wang, X. The plasma membrane-bound phospholipase Ddelta enhances freezing tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2004, 22, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, R.; Li, M.; Li, L.; Wang, C.; Welti, R.; Wang, X. Differential degradation of extraplastidic and plastidic lipids during freezing and post-freezing recovery in Arabidopsis thaliana. J. Biol. Chem. 2008, 283, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Wang, X. Phosphatidic acid: An emerging versatile class of cellular mediators. Essays Biochem. 2020, 64, 533–546. [Google Scholar] [PubMed]
- Fan, J.; Yan, C.; Roston, R.; Shanklin, J.; Xu, C. Arabidopsis lipins, PDAT1 acyltransferase, and SDP1 triacylglycerol lipase synergistically direct fatty acids toward β-oxidation, thereby maintaining membrane lipid homeostasis. Plant Cell 2014, 26, 4119–4134. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.K.; Johansson, O.N.; Fahlberg, P.; Kommuri, M.; Töpel, M.; Bodin, L.J.; Sikora, P.; Modarres, M.; Ekengren, S.; Nguyen, C.T.; et al. Acylated monogalactosyl diacylglycerol: Prevalence in the plant kingdom and identification of an enzyme catalyzing galactolipid head group acylation in Arabidopsis thaliana. Plant J. 2015, 84, 1152–1166. [Google Scholar] [CrossRef]
- Nilsson, A.K.; Fahlberg, P.; Johansson, O.N.; Hamberg, M.; Andersson, M.X.; Ellerström, M. The activity of HYDROPEROXIDE LYASE 1 regulates accumulation of galactolipids containing 12-oxo-phytodienoic acid in Arabidopsis. J. Exp. Bot. 2016, 67, 5133–5144. [Google Scholar] [CrossRef]
- Andersson, M.X.; Hamberg, M.; Kourtchenko, O.; Brunnström, A.; McPhail, K.L.; Gerwick, W.H.; Göbel, C.; Feussner, I.; Ellerström, M. Oxylipin profiling of the hypersensitive response in Arabidopsis thaliana. Formation of a novel oxo-phytodienoic acid-containing galactolipid, arabidopside E. J. Biol. Chem. 2006, 281, 31528–31537. [Google Scholar]
- Buseman, C.M.; Tamura, P.; Sparks, A.A.; Baughman, E.J.; Maatta, S.; Zhao, J.; Roth, M.R.; Esch, S.W.; Shah, J.; Williams, T.D.; et al. Wounding stimulates the accumulation of glycerolipids containing oxophytodienoic acid and dinor-oxophytodienoic acid in Arabidopsis leaves. Plant Physiol. 2006, 142, 28–39. [Google Scholar] [CrossRef]
- Ibrahim, A.; Schütz, A.L.; Galano, J.M.; Herrfurth, C.; Feussner, K.; Durand, T.; Brodhun, F.; Feussner, I. The alphabet of galactolipids in Arabidopsis thaliana. Front. Plant Sci. 2011, 2, 95. [Google Scholar] [CrossRef]
- Zuther, E.; Schaarschmidt, S.; Fischer, A.; Erban, A.; Pagter, M.; Mubeen, U.; Giavalisco, P.; Kopka, J.; Sprenger, H.; Hincha, D.K. Molecular signatures associated with increased freezing tolerance due to low temperature memory in Arabidopsis. Plant Cell Environ. 2019, 42, 854–873. [Google Scholar] [PubMed]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate regulates the inducer of cbf expression-C-repeat binding factor/DRE binding factor1 cascade and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed]
- Pigolev, A.V.; Miroshnichenko, D.N.; Pushin, A.S.; Terentyev, V.V.; Boutanayev, A.M.; Dolgov, S.V.; Savchenko, T.V. Overexpression of Arabidopsis OPR3 in hexaploid wheat (Triticum aestivum L.) alters plant development and freezing tolerance. Int. J. Mol. Sci. 2018, 19, 3989. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.F.; Xu, L.; Tan, W.J.; Chen, L.; Qi, H.; Xie, L.J.; Chen, M.X.; Liu, B.Y.; Yu, L.J.; Yao, N.; et al. Disruption of the Arabidopsis defense regulator genes SAG101, EDS1, and PAD4 confers enhanced freezing tolerance. Mol. Plant 2015, 8, 1536–1549. [Google Scholar] [CrossRef]
- Zoeller, M.; Stingl, N.; Krischke, M.; Fekete, A.; Waller, F.; Berger, S.; Mueller, M.J. Lipid profiling of the Arabidopsis hypersensitive response reveals specific lipid peroxidation and fragmentation processes: Biogenesis of pimelic and azelaic acid. Plant Physiol. 2012, 160, 365–378. [Google Scholar] [CrossRef]
- Hisamatsu, Y.; Goto, N.; Hasegawa, K.; Shigemori, H. Arabidopsides A and B, two new oxylipins from Arabidopsis thaliana. Tetrahedron Lett. 2003, 44, 5553–5556. [Google Scholar] [CrossRef]
- Glauser, G.; Grata, E.; Rudaz, S.; Wolfender, J.L. High-resolution profiling of oxylipin containing galactolipids in Arabidopsis extracts by ultraperformance liquid 9 chromatography/time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2018, 22, 3154–3160. [Google Scholar] [CrossRef]
- Stelmach, B.A.; Muller, A.; Hennig, P.; Gebhardt, S.; Schubert-Zsilavecz, M.; Weiler, E.W. A novel class of oxylipins, sn1-O-(12-oxophytodienoyl)-sn2-O- (hexadecatrienoyl)-monogalactosyl diglyceride, from Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 12832–12838. [Google Scholar] [CrossRef]
- Hisamatsu, Y.; Goto, N.; Sekiguchi, M.; Hasegawa, K.; Shigemori, H. Oxylipins arabidopsides C and D from Arabidopsis thaliana. J. Nat. Prod. 2005, 68, 600–603. [Google Scholar] [CrossRef]
- Grun, G.; Berger, S.; Matthes, D.; Mueller, M.J. Early accumulation of nonenzymatically synthesized oxylipins in Arabidopsis thaliana after infection with Pseudomonas syringae. Funct. Plant Biol. 2007, 34, 65–71. [Google Scholar] [CrossRef]
- Chen, G.; Greer, M.S.; Weselake, R.J. Plant phospholipase A: Advances in molecular biology, biochemistry, and cellular function. Biomol. Concepts 2013, 4, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Narusaka, Y.; Narusaka, M.; Seki, M.; Fujita, M.; Ishida, J.; Nakashima, M.; Enju, A.; Sakurai, T.; Satou, M.; Kamiya, A.; et al. Expression profiles of Arabidopsis phospholipase A IIA gene in response to biotic and abiotic stresses. Plant Cell Physiol. 2003, 44, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Y.; Zheng, Y.; Bahn, S.C.; Pan, X.Q.; Li, M.Y.; Vu, H.S.; Roth, M.R.; Scheu, B.; Welti, R.; Hong, Y.Y.; et al. The patatin-containing phospholipase A pPLAIIα modulates oxylipin formation and water loss in Arabidopsis thaliana. Mol. Plant 2012, 5, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Falk, A.; Feys, B.J.; Frost, L.N.; Jones, J.D.; Daniels, M.J.; Parker, J.E. EDS1, an essential component of R gene-mediated disease resistance in Arabidopsis has homology to eukaryotic lipases. Proc. Natl. Acad. Sci. USA 1999, 96, 3292–3297. [Google Scholar] [CrossRef]
- Jirage, D.; Tootle, T.L.; Reuber, T.L.; Frost, L.N.; Feys, B.J.; Parker, J.E.; Ausubel, F.M.; Glazebrook, J. Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc. Natl. Acad. Sci. USA 1999, 96, 13583–13588. [Google Scholar] [CrossRef]
- He, Y.; Gans, S. A gene encoding an acyl hydrolase is involved in leaf senescence in Arabidopsis. Plant Cell 2002, 14, 805–815. [Google Scholar] [CrossRef]
- Heinz, E. Uber die enzymatische Bildung von Acylgalaktosyldiglycerid [On the enzymatic formation of acylgalactosyldiglyceride]. Biochim. Biophys. Acta 1967, 144, 333–343. [Google Scholar] [CrossRef]
- Mueller, S.P.; Unger, M.; Guender, L.; Fekete, A.; Mueller, M.J. Phospholipid:diacylglycerol acyltransferase-mediated triacylglycerol synthesis augments basal thermotolerance. Plant Physiol. 2017, 175, 486–497. [Google Scholar] [CrossRef]
- Demski, K.; Łosiewska, A.; Jasieniecka-Gazarkiewicz, K.; Klińska, S.; Banaś, A. Phospholipid:diacylglycerol acyltransferase1 overexpression delays senescence and enhances post-heat and cold exposure fitness. Front. Plant Sci. 2020, 11, 611897. [Google Scholar] [CrossRef]
- Uemura, M.; Steponkus, P.L. A contrast of the plasma membrane lipid composition of oat and rye leaves in relation to freezing tolerance. Plant Physiol. 1994, 104, 479–496. [Google Scholar] [CrossRef]
- Mishra, M.K.; Singh, G.; Tiwari, S.; Singh, R.; Kumari, N.; Misra, P. Characterization of Arabidopsis sterol glycosyltransferase TTG15/UGT80B1 role during freeze and heat stress. Plant Signal. Behav. 2015, 10, e1075682. [Google Scholar] [CrossRef] [PubMed]
- Dunn, O.J. Multiple comparisons among means. J. Am. Stat. Assoc. 1961, 56, 52–64. [Google Scholar] [CrossRef]
- Etymologia: Bonferroni correction. Emerg. Infect. Dis. 2015, 21, 289.
- Buseman, C.M. New Arabidopsis Galactolipids Containing Esterified Oxylipins. Master’s Thesis, Kansas State University, Manhattan, KS, USA, 2005. [Google Scholar]
- Telfer, A.; Bollman, K.M.; Poethig, R.S. Phase change and the regulation of trichome distribution in Arabidopsis thaliana. Development 1997, 124, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Available online: https://www.metaboanalyst.ca/ (accessed on 7 November 2021).
- Janková, J.; van de Geer, S. Confidence intervals for high-dimensional inverse covariance estimation. Electron. J. Stat. 2015, 9, 1205–1229. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; Anderson de Lima Morais, D.; Chang, l.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Nalam, V.J.; Keeretaweep, J.; Sarowar, S.; Shah, J. Root-derived oxylipins promote green peach aphid performance on Arabidopsis foliage. Plant Cell 2012, 24, 1643–1653. [Google Scholar] [CrossRef]
- Bi, D.; Cheng, Y.T.; Li, X.; Zhang, Y. Activation of plant immune responses by a gain-of-function mutation in an atypical receptor-like kinase. Plant Physiol. 2010, 153, 1771–1779. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Putative Identification | Reference(s) to Fatty Acid in Plants and/or in Complex Lipids |
|---|---|---|
| 7:1;O | 7-oxoheptanoic acid | [35] |
| 7:1;O2 | pimelic acid | [35] |
| 9:1;O | 9-oxononanoic acid | [35] |
| 9:1;O2 | azelaic acid | [35] |
| 16:4;O | dinorOPDA | [28,29,36,37] |
| 16:3;O | hydroxy 16:3 | |
| 18:5;O2 | unknown | |
| 18:4;O | OPDA, keto 18:3 | [28,29,30,36,37,38,39] |
| 18:4,O3 | unknown | |
| 18:3;O | hydroxy 18:3, keto fatty acid | [30] |
| 18:3;O2 | ketol fatty acid, hydroperoxy 18:3, dihydroxy 18:3 | [29,30] |
| 18:3;O3 | phytoprostane | [40] |
| 18:2;O | hydroxy 18:2 |
| Leaf Number at 20 Days | Leaf Number at 27 Days | Rosette Dry Mass | |
|---|---|---|---|
| wild type | 5.16 ± 0.82 | 9.25 ± 1.21 | 9.04 ± 5.69 |
| pplaIIα | 5.47 ± 0.78 | 10.10 ± 0.98 H | 9.40 ± 5.73 |
| pplaIIα × pplaIIIβ | 5.71 ± 0.67 H | 10.08 ± 1.00 H | 10.38 ± 5.78 |
| lox1-1 | 4.33 ± 0.89 L | 7.71 ± 1.25 L | 6.23 ± 3.72 L |
| lox5-1 | 4.50 ± 0.87 L | 8.35 ± 1.68 L | 7.79 ± 5.53 |
| lox1-1 × lox5-1 | 4.47 ± 0.99 L | 8.37 ± 1.48 L | 7.96 ± 8.44 |
| opr3-2 | 4.54 ± 0.77 L | 8.37 ± 1.00 L | 8.93 ± 6.29 |
| Part 1. Negative Correlations of Level at 74 h with Final Ion Leakage, i.e., Associated with Good Outcome. | |||
|---|---|---|---|
| Hypothesis: Increasing the Amounts of “Lipid or Lipid group” Produced During Freezing Will Increase Freezing Tolerance. | |||
| Lipid or Lipid Group | Possible Role in Freezing Sensitivity or Tolerance | Supporting Evidence from Previous Work/Comments | Status |
| Many structural polar lipids, especially those with polyunsaturated fatty acyl chains (Figure 5) | Form and stabilize membranes | Extensive biophysical evidence indicates most lipids in this group participate in bilayer structures. This group of lipids is the only group at 98 h negatively correlated with ion leakage at 98 h (Figure 4b), consistent with these lipids being present when plants are recovered. | |
| Structural lipids with very long-chain fatty acyl chains (Figure 5) | May reduce propensity for non-bilayer phases caused by dehydration during ice formation | Genetic manipulation of very long-chain fatty acid content shows higher content of very long-chain fatty acids is associated with chilling tolerance [20]. | Some evidence in chilling |
| Polygalactosylated diacylglycerols (Figure 6) | Stabilize the chloroplast envelope | The sfr2 mutant has a poor outcome upon freezing compared to wild type [5]. | Good evidence |
| PA 34:6 (i.e., PA 18:3_16:3) (Figure 7) | Is a byproduct of polygalactosylated diacylglycerol synthesis | Synthesis of PA 34:6 after wounding is highly correlated with polygalactosylated diacylglycerol synthesis. It may be a side product rather than causative [14]. | No direct evidence |
| Oxidized head group-acylated plastidic lipids (Figure 8), oxidized polar diacyl lipids (Figure 9), and OPDA-containing sterol derivatives (Figure 12) | Some may serve as a sink for reactive oxygen species; some might serve as signaling molecules. | OPDA-containing species are formed by the same pathway as jasmonates, which activate the ICE-CBF/DREB1 pathway leading to freezing tolerance [32]. | No direct evidence about the role of the measured oxidized lipids |
| Monoacyl polar lipids (Figure 10) | May be intermediates in lipid remodeling | DGMG and MGMG can be formed during head-group acylation of plastidic lipids | No direct evidence |
| Triacylglycerols (Figure 11) | Sequester free fatty acids removed during lipid remodeling | DGAT1 overexpression increases freezing tolerance [11,12]. The role of PDAT1 remains to be tested. | Moderate evidence |
| Sterol esters (Figure 12) | May serve as a reservoir for a small amount of fatty acids removed from membranes | No evidence | |
| Part 2. Positive Correlations of Level at 74 h with Final Ion Leakage, i.e., Associated with Poor Outcome. | |||
| Hypothesis: Decreasing the amounts of “Lipid or lipid group” produced during freezing will increase freezing tolerance. | |||
| Lipid or Lipid Group | Possible Role in Freezing Sensitivity or Tolerance | Supporting Evidence from Previous Work/Comments | Status |
| Some MGDGs without full unsaturation (Figure 5) | May cause chloroplast membranes to be too rigid at low temperatures, causing photoinhibition | Defects in chloroplast desaturases lead to photoinhibition and poor growth in cold [2,9]. | Good evidence for effect in cold, but effect in freezing is less clear |
| PAs not containing 16:3 (Figure 7) | May destabilize extra-plastidic membranes by facilitating hexagonal phase formation | PLDα1 suppression increases freezing survival and decreases ion leakage [4]. Knockout of DGKs decreases freezing tolerance [11]. | Good evidence for effect, but mechanism of PA action is unproven |
| Non-oxidized head group-acylated MGDGs (Figure 8) | May destabilize membrane structure when present at high levels, as occurs after freezing | No evidence | |
| Non-oxidized acyl sterol hexosides (Figure 12) | May destabilize extra-plastidic membranes by facilitating hexagonal phase formation | Acyl sterol hexosides decrease in cold acclimation and are present at lower levels in the plasma membrane of a freeze-tolerant rye than in a freeze-susceptible oat [50]. | Associative evidence only |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, H.S.; Shiva, S.; Samarakoon, T.; Li, M.; Sarowar, S.; Roth, M.R.; Tamura, P.; Honey, S.; Lowe, K.; Porras, H.; et al. Specific Changes in Arabidopsis thaliana Rosette Lipids during Freezing Can Be Associated with Freezing Tolerance. Metabolites 2022, 12, 385. https://doi.org/10.3390/metabo12050385
Vu HS, Shiva S, Samarakoon T, Li M, Sarowar S, Roth MR, Tamura P, Honey S, Lowe K, Porras H, et al. Specific Changes in Arabidopsis thaliana Rosette Lipids during Freezing Can Be Associated with Freezing Tolerance. Metabolites. 2022; 12(5):385. https://doi.org/10.3390/metabo12050385
Chicago/Turabian StyleVu, Hieu Sy, Sunitha Shiva, Thilani Samarakoon, Maoyin Li, Sujon Sarowar, Mary R. Roth, Pamela Tamura, Samuel Honey, Kaleb Lowe, Hollie Porras, and et al. 2022. "Specific Changes in Arabidopsis thaliana Rosette Lipids during Freezing Can Be Associated with Freezing Tolerance" Metabolites 12, no. 5: 385. https://doi.org/10.3390/metabo12050385
APA StyleVu, H. S., Shiva, S., Samarakoon, T., Li, M., Sarowar, S., Roth, M. R., Tamura, P., Honey, S., Lowe, K., Porras, H., Prakash, N., Roach, C. A., Stuke, M., Wang, X., Shah, J., Gadbury, G., Wang, H., & Welti, R. (2022). Specific Changes in Arabidopsis thaliana Rosette Lipids during Freezing Can Be Associated with Freezing Tolerance. Metabolites, 12(5), 385. https://doi.org/10.3390/metabo12050385