
Plasma Lipid Profiles Change with Increasing Numbers of Mild Traumatic Brain Injuries in Rats

 , , and
, , and
Abstract
:1. Introduction
2. Results
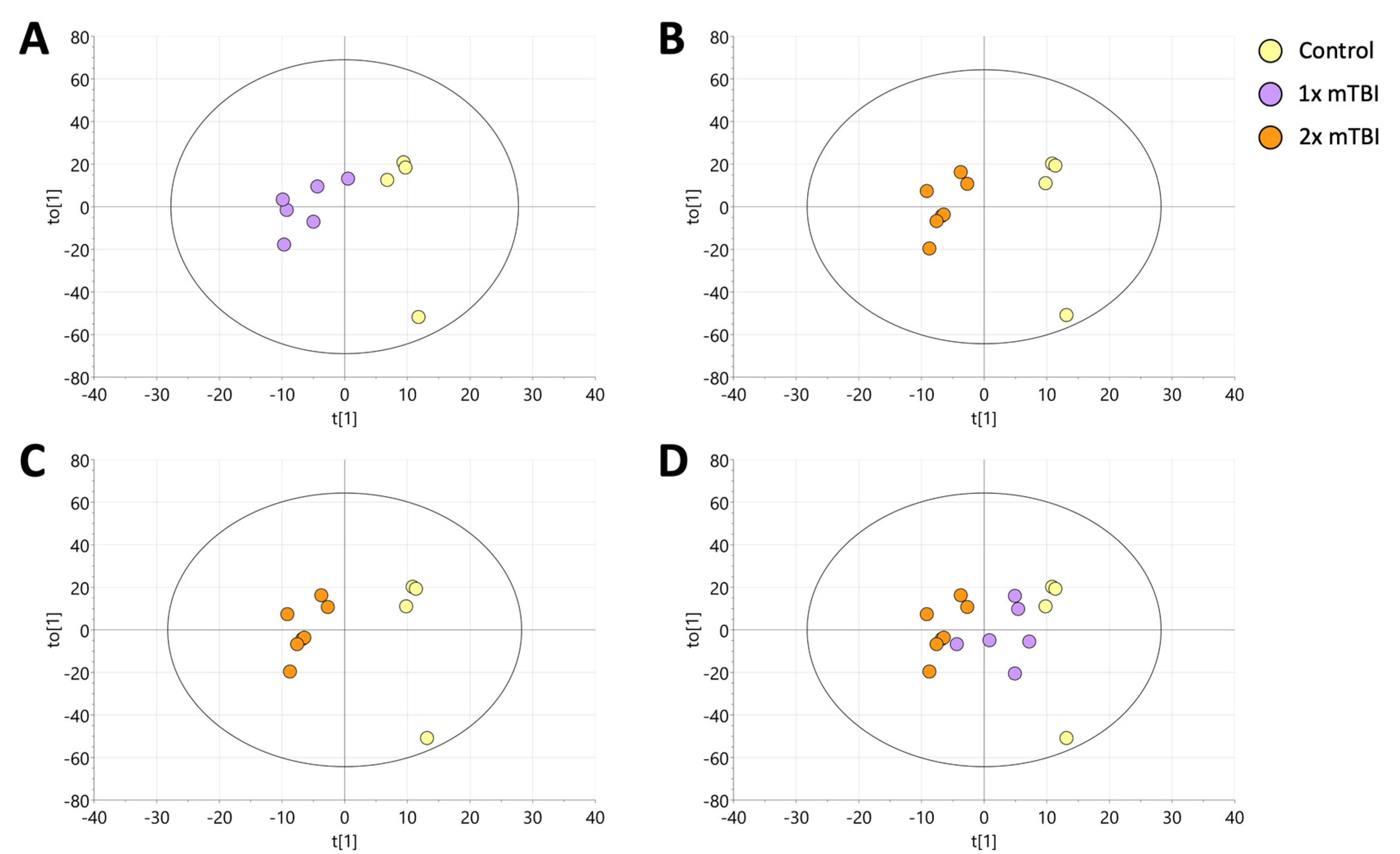
2.1. Orthogonal Projection to Latent Structure-Discriminant Analysis (OPLS-DA) of Plasma Lipids Discriminates Sham, 1× mTBI and 2× mTBI
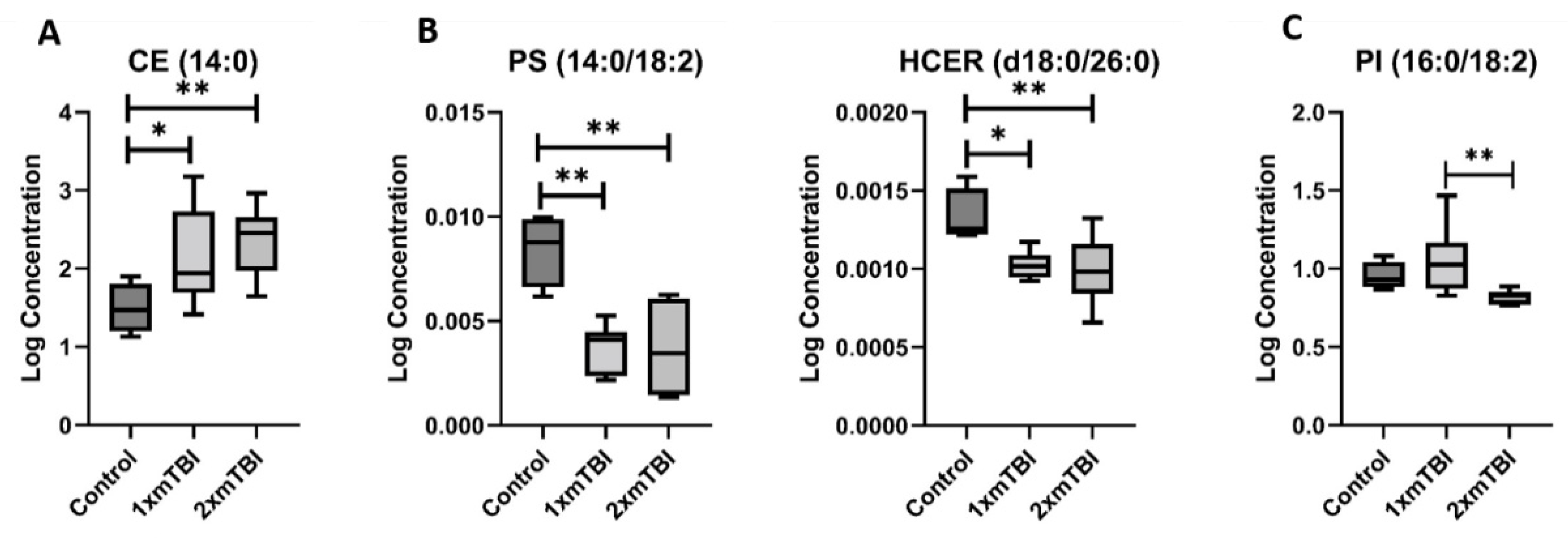
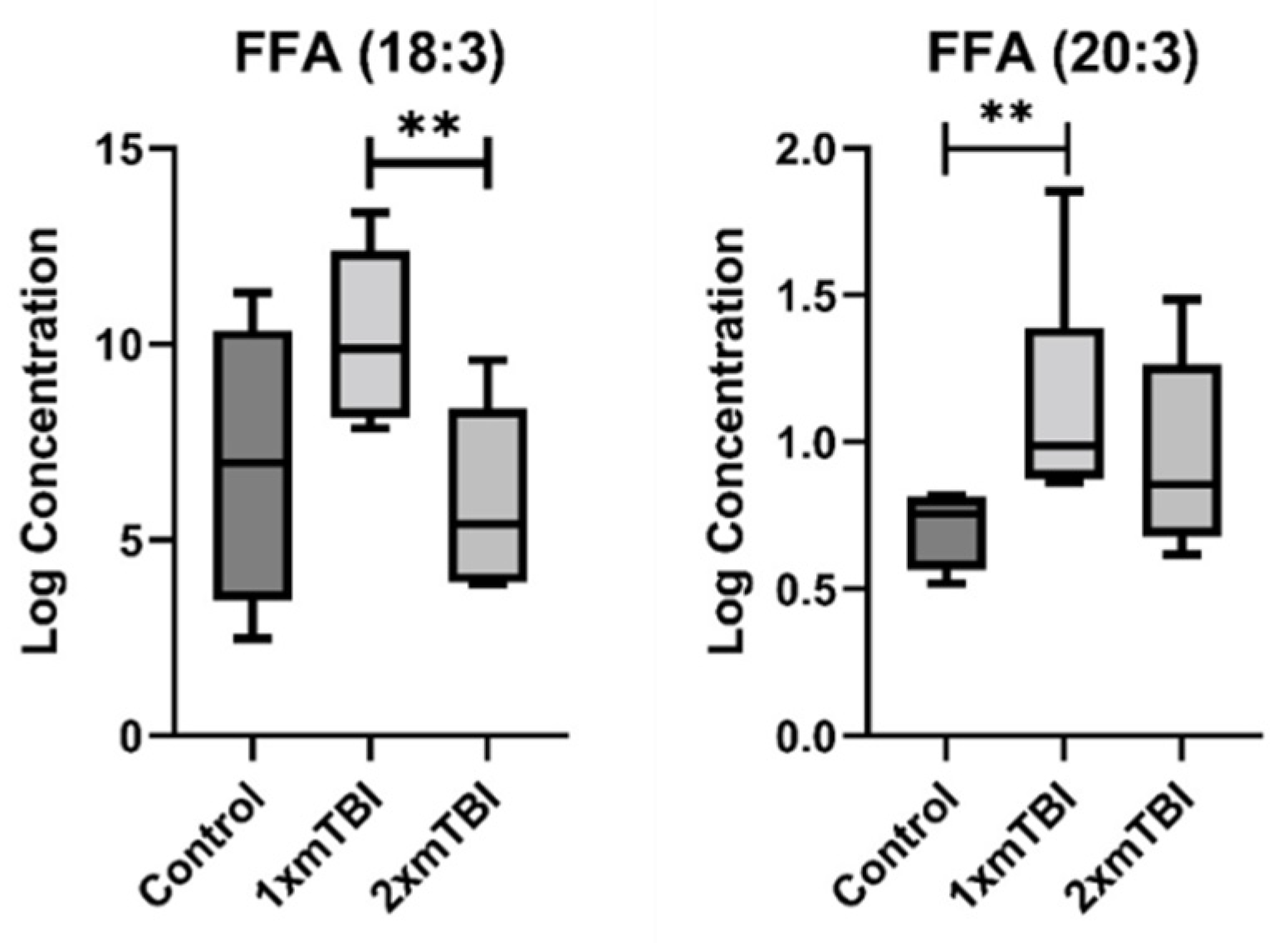
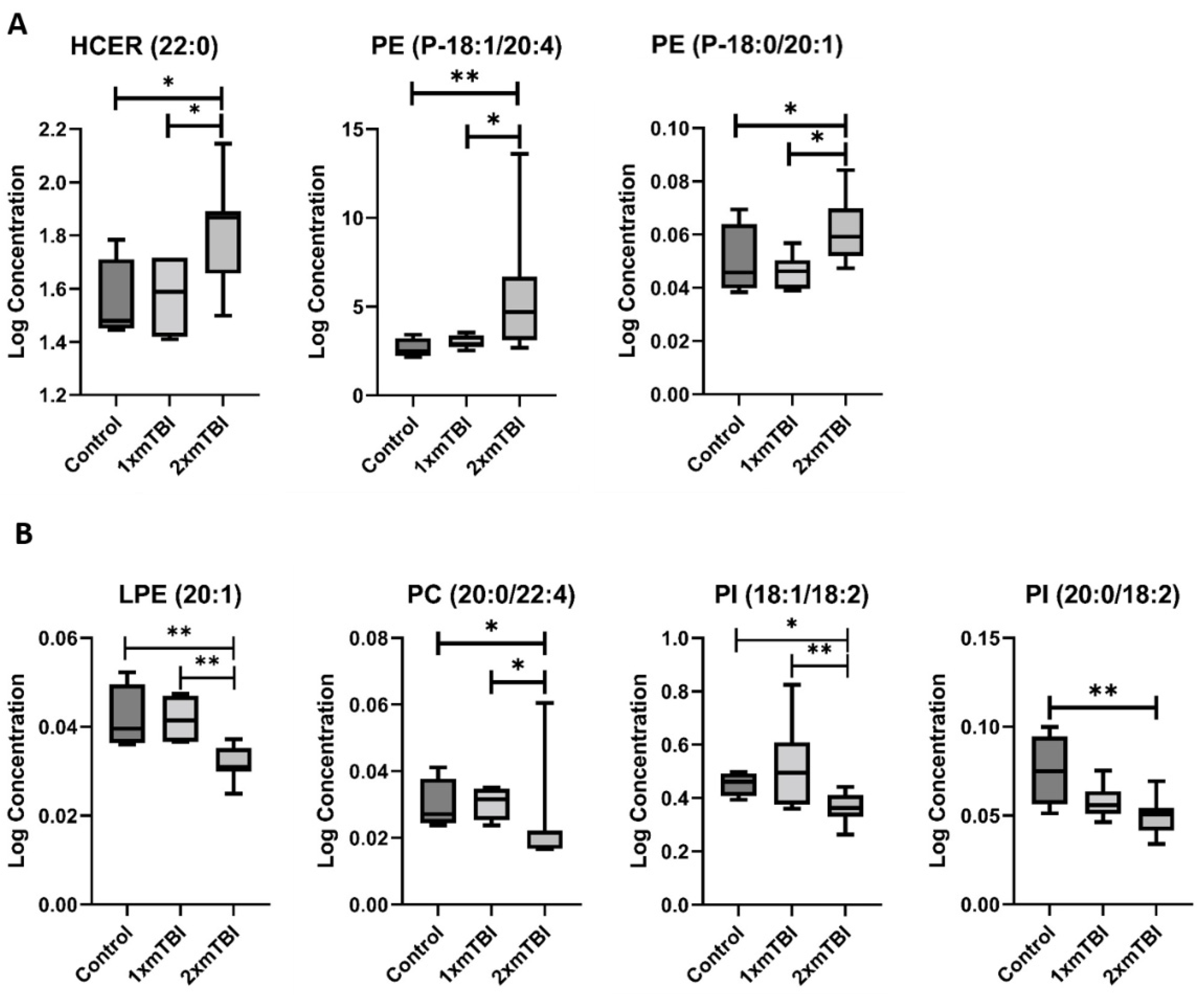
2.2. Individual Plasma Metabolites Are Altered Differentially in Response to 1× or 2× mTBI
3. Discussion
4. Materials and Methods
4.1. Animals and Ethics
4.2. Closed Head Weight-Drop Nodel of Repeated Mild Traumatic Brain Injury
4.3. Plasma Collection for Lipidomics
4.4. Sample Preparation for Liquid Chromatography–Mass Spectrometry (LC-MS)
4.5. Liquid Chromatography Mass Spectrometry (LC-MS)
4.6. Data Pre-Processing
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CE | Cholesterolester |
| FFA | Free fatty acid |
| HCER | Hexosylceramide |
| LC-MS | Liquid chromatography-mass spectrometry |
| LPE | Lysophosphatidylethanolamine |
| LPG | Lysophosphoglycerol |
| mTBI | Mild traumatic brain injury |
| OPLS-DA | Orthogonal projection to latent structure-discriminant analysis |
| PC | Phosphocholine |
| PCA | Principal components analysis |
| PE | Phosphoethanolamine |
| PI | Phosphoinositol |
| PS | Phosphoserine |
| QC | Quality control |
| rmTBI | Repeated mild traumatic brain injury |
| SM | Sphingomyelin |
| TBI | Traumatic brain injury |
References
- Thomas, E.; Fitzgerald, M.; Cowen, G. Post-concussion states: How do we improve our patients’ outcomes? An Australian perspective. J. Concussion 2020, 4, 205970022096031. [Google Scholar] [CrossRef]
- Belanger, H.G.; Spiegel, E.; Vanderploeg, R.D. Neuropsychological performance following a history of multiple self-reported concussions: A meta-analysis. J. Int. Neuropsychol. Soc. 2010, 16, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Martini, D.N.; Broglio, S.P. Long-term effects of sport concussion on cognitive and motor performance: A review. Int. J. Psychophysiol. 2018, 132, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Omalu, B.; Bailes, J.; Hamilton, R.L.; Kamboh, M.I.; Hammers, J.; Case, M.; Fitzsimmons, R. Emerging Histomorphologic Phenotypes of Chronic Traumatic Encephalopathy in American Athletes. Neurosurgery 2011, 69, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Decq, P.; Gault, N.; Blandeau, M.; Kerdraon, T.; Berkal, M.; Elhelou, A.; Dusfour, B.; Peyrin, J.-C. Long-term consequences of recurrent sports concussion. Acta Neurochir. (Wien) 2016, 158, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Giza, C.C.; Hovda, D.A. The New Neurometabolic Cascade of Concussion. Neurosurgery 2014, 75 (Suppl. 4), S24–S33. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Muneer, P.; Schuetz, H.; Wang, F.; Skotak, M.; Jones, J.; Gorantla, S.; Zimmerman, M.C.; Chandra, N.; Haorah, J. Induction of Oxidative and Nitrosative Damage Leads to Cerebrovascular Inflammation in an Animal Model of Mild Traumatic brain Injury Induced by Primary Blast. Free Radic. Biol. Med. 2013, 60, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fresco, G.P.; Sousa, A.D.; Pillai, A.M.; Moy, S.S.; Crawley, J.N.; Tessarollo, L.; Dupree, J.L.; Bhat, M.A. Disruption of Axo–Glial Junctions Causes Cytoskeletal Disorganization and Degeneration of Purkinje Neuron Axons. Proc. Natl. Acad. Sci. USA 2006, 103, 5137–5142. [Google Scholar] [CrossRef] [Green Version]
- Nessel, I.; Michael-Titus, A.T. Lipid profiling of brain tissue and blood after traumatic brain injury: A review of human and experimental studies. Semin. Cell Dev. Biol. 2021, 112, 145–156. [Google Scholar] [CrossRef]
- Emmerich, T.; Abdullah, L.; Ojo, J.; Mouzon, B.; Nguyen, T.; Laco, G.S.; Crynen, G.; Evans, J.E.; Reed, J.; Mullan, M.; et al. Mild TBI Results in a Long-Term Decrease in Circulating Phospholipids in a Mouse Model of Injury. NeuroMolecular Med. 2016, 19, 122–135. Available online: https://link.springer.com/article/10.1007/s12017-016-8436-4 (accessed on 29 September 2021). [CrossRef] [PubMed]
- Bozek, K.; Wei, Y.; Yan, Z.; Liu, X.; Xiong, J.; Sugimoto, M.; Tomita, M.; Pääbo, S.; Sherwood, C.C.; Hof, P.R.; et al. Organization and Evolution of Brain Lipidome Revealed by Large-Scale Analysis of Human, Chimpanzee, Macaque, and Mouse Tissues. Neuron 2015, 85, 695–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunshine, H.; Iruela-Arispe, M.L. Membrane Lipids and Cell Signaling. Curr. Opin. Lipidol. 2017, 28, 408–413. Available online: https://pubmed.ncbi.nlm.nih.gov/28692598/ (accessed on 22 October 2021). [CrossRef] [PubMed]
- Smith, Q.R.; Nagura, H. Fatty Acid Uptake and Incorporation in Brain: Studies with the Perfusion Model. J. Mol. Neurosci. 2001, 16, 167–172. [Google Scholar] [CrossRef]
- Rapoport, S.I.; Chang, M.J.C.; Spector, A.A. Delivery and Turnover of Plasma-Derived Essential PUFAs in Mammalian Brain. J. Lipid Res. 2001, 42, 678–685. Available online: http://www.jlr.org/article/S0022227520316291/fulltext (accessed on 1 October 2021). [CrossRef]
- Daley, M.; Dekaban, G.A.; Bartha, R.; Brown, A.; Stewart, T.C.; Doherty, T.; Fischer, L.; Holmes, J.; Menon, R.; Rupar, C.A.; et al. Metabolomics Profiling of Concussion in Adolescent Male Hockey Players: A Novel Diagnostic Method. Metabolomics 2016, 12, 185. Available online: https://link.springer.com/article/10.1007/s11306-016-1131-5 (accessed on 1 October 2021). [CrossRef]
- Emmerich, M.T.; Abdullah, L.; Crynen, G.; Dretsch, M.N.; Evans, J.; Ait-Ghezala, G.; Reed, J.; Montague, M.H.; Chaytow, H.; Mathura, V.S.; et al. Plasma Lipidomic Profiling in a Military Population of Mild Traumatic Brain Injury and Post-Traumatic Stress Disorder with Apolipoprotein E ɛ4-Dependent Effect. J. Neurotrauma 2016, 33, 1331–1348. Available online: https://www.liebertpub.com/doi/abs/10.1089/neu.2015.4061 (accessed on 1 October 2021). [CrossRef] [PubMed]
- Ojo, J.O.; Algamal, M.; Leary, P.; Abdullah, L.; Mouzon, B.; Evans, J.E.; Mullan, M.; Crawford, F. Disruption in Brain Phospholipid Content in a Humanized Tau Transgenic Model Following Repetitive Mild Traumatic Brain Injury. Front. Neurosci. 2018, 12, 893. [Google Scholar] [CrossRef] [Green Version]
- Meaney, D.F.; Smith, D.H. Biomechanics of Concussion. Clin. Sports Med. 2011, 30, 19–31. Available online: https://pubmed.ncbi.nlm.nih.gov/21074079/ (accessed on 13 March 2022). [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Black, A.M.B.; Milbourn, H.; Krakonja, S.; Nesbit, M.; Bartlett, C.A.; Fehily, B.; Takechi, R.; Yates, N.J.; Fitzgerald, M. The Effects of a Combination of Ion Channel Inhibitors in Female Rats Following Repeated Mild Traumatic Brain Injury. Int. J. Mol. Sci. 2018, 19, 3408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyaegbu, C.C.; Mao, Y.; McGonigle, T.; Raja, S.; Clarke, T.; Black, A.M.B.; Solomon, T.; Fuller, K.; Fitzgerald, M. Simultaneous flow cytometric characterization of multiple cell types and metabolic states in the rat brain after repeated mild traumatic brain injury. J. Neurosci. Methods 2021, 359, 109223. [Google Scholar] [CrossRef]
- Yates, N.; Lydiard, S.; Fehily, B.; Weir, G.; Chin, A.; Bartlett, C.A.; Alderson, J.; Fitzgerald, M. Repeated mild traumatic brain injury in female rats increases lipid peroxidation in neurons. Exp. Brain Res. 2017, 235, 2133–2149. [Google Scholar] [CrossRef] [Green Version]
- Fehily, B.; Bartlett, C.A.; Lydiard, S.; Archer, M.; Milbourn, H.; Majimbi, M.; Hemmi, J.M.; Dunlop, S.A.; Yates, N.; Fitzgerald, M. Differential responses to increasing numbers of mild traumatic brain injury in a rodent closed-head injury model. J. Neurochem. 2019, 149, 660–678. [Google Scholar] [CrossRef]
- Emmerich, T.; Zakirova, Z.; Klimas, N.; Sullivan, K.; Shetty, A.K.; Evans, J.E.; Ait-Ghezala, G.; Laco, G.S.; Hattiangady, B.; Shetty, G.A.; et al. Phospholipid profiling of plasma from GW veterans and rodent models to identify potential biomarkers of Gulf War Illness. PLoS ONE 2017, 12, e0176634. [Google Scholar] [CrossRef] [PubMed]
- Fiandaca, M.S.; Mapstone, M.; Mahmoodi, A.; Gross, T.; Macciardi, F.; Cheema, A.K.; Merchant-Borna, K.; Bazarian, J.; Federoff, H.J. Plasma metabolomic biomarkers accurately classify acute mild traumatic brain injury from controls. PLoS ONE 2018, 13, e0195318. [Google Scholar] [CrossRef] [Green Version]
- Hogan, S.R.; Phan, J.H.; Alvarado-Velez, M.; Wang, M.D.; Bellamkonda, R.V.; Fernández, F.M.; LaPlaca, M.C. Discovery of Lipidome Alterations Following Traumatic Brain Injury via High-Resolution Metabolomics. J. Proteome Res. 2018, 17, 2131–2143. Available online: https://pubmed.ncbi.nlm.nih.gov/29671324/ (accessed on 11 October 2021). [CrossRef]
- Huguenard, C.J.C.; Cseresznye, A.; Evans, J.E.; Oberlin, S.; Langlois, H.; Ferguson, S.; Darcey, T.; Nkiliza, A.; Dretsch, M.; Mullan, M.; et al. Plasma Lipidomic Analyses in Cohorts With mTBI and/or PTSD Reveal Lipids Differentially Associated with Diagnosis and APOE ε4 Carrier Status. Front. Physiol. 2020, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Poblete, R.A.; Arenas, M.; Sanossian, N.; Freeman, W.D.; Louie, S.G. The Role of Bioactive Lipids in Attenuating the Neuroinflammatory Cascade in Traumatic Brain Injury. Ann. Clin. Transl. Neurol. 2020, 7, 2524–2534. [Google Scholar] [CrossRef]
- Roux, A.; Muller, L.; Jackson, S.N.; Post, J.; Baldwin, K.; Hoffer, B.; Balaban, C.; Barbacci, D.; Schultz, J.A.; Gouty, S.; et al. Mass Spectrometry Imaging of Rat Brain Lipid Profile Changes over Time Following Traumatic Brain Injury. J. Neurosci. Methods 2016, 272, 19–32. Available online: https://pubmed.ncbi.nlm.nih.gov/26872743/ (accessed on 22 October 2021). [CrossRef] [PubMed] [Green Version]
- Ye, L.; Sun, Y.; Jiang, Z.; Wang, G. L-Serine, an Endogenous Amino Acid, Is a Potential Neuroprotective Agent for Neurological Disease and Injury. Front. Mol. Neurosci. 2021, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Harizi, H.; Corcuff, J.-B.; Gualde, N. Arachidonic-Acid-Derived Eicosanoids: Roles in Biology and Immunopathology. Trends Mol. Med. 2008, 14, 461–469. Available online: https://pubmed.ncbi.nlm.nih.gov/18774339/ (accessed on 22 October 2021). [CrossRef]
- Anthonymuthu, T.S.; Kenny, E.M.; Amoscato, A.A.; Lewis, J.; Kochanek, P.M.; Kagan, V.E.; Bayır, H. Global assessment of oxidized free fatty acids in brain reveals an enzymatic predominance to oxidative signaling after trauma. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 2601–2613. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.I.; Higgs, G.A. Eicosanoids and Inflammation. J. Pathol. 1988, 156, 101–110. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/path.1711560204 (accessed on 22 October 2021). [CrossRef] [PubMed]
- Fabelo, N.; Martín, V.; Santpere, G.; Marín, R.; Torrent, L.; Ferrer, I.; Díaz, M. Severe Alterations in Lipid Composition of Frontal Cortex Lipid Rafts from Parkinson’s Disease and Incidental Parkinson’s Disease. Mol. Med. 2011, 17, 1107–1118. Available online: https://pubmed.ncbi.nlm.nih.gov/21717034/ (accessed on 21 October 2021). [CrossRef]
- Han, X.; Holtzman, D.M.; McKeel, D.W., Jr. Plasmalogen Deficiency in Early Alzheimer’s Disease Subjects and in Animal Models: Molecular Characterization Using Electrospray Ionization mass Spectrometry. J. Neurochem. 2001, 77, 1168–1180. Available online: https://pubmed.ncbi.nlm.nih.gov/11359882/ (accessed on 21 October 2021). [CrossRef]
- Mallah, K.; Quanico, J.; Raffo-Romero, A.; Cardon, T.; Aboulouard, S.; Devos, D.; Kobeissy, F.; Zibara, K.; Salzet, M.; Fournier, I. Matrix-Assisted Laser Desorption/Ionization-Mass Spectrometry Imaging of Lipids in Experimental Model of Traumatic Brain Injury Detecting Acylcarnitines as Injury Related Markers. Anal. Chem. 2019, 91, 11879–11887. [Google Scholar] [CrossRef]
- Abdullah, L.; Evans, J.E.; Ferguson, S.; Mouzon, B.; Montague, H.; Reed, J.; Crynen, G.; Emmerich, T.; Crocker, M.; Pelot, R.; et al. Lipidomic Analyses Identify Injury-Specific Phospholipid Changes 3 mo after Traumatic Brain Injury. FASEB J. 2014, 28, 5311–5321. [Google Scholar] [CrossRef] [PubMed]
- Gier, E.C.; Pulliam, A.N.; Gaul, D.A.; Moore, S.G.; LaPlaca, M.C.; Fernández, F.M. Lipidome Alterations following Mild Traumatic Brain Injury in the Rat. Metabolites 2022, 12, 150. Available online: https://www.mdpi.com/2218-1989/12/2/150/htm (accessed on 5 February 2022). [CrossRef]
- Ojo, J.O.; Algamal, M.; Leary, P.; Abdullah, L.; Mouzon, B.; Evans, J.E.; Mullan, M.; Crawford, F. Converging and Differential Brain Phospholipid Dysregulation in the Pathogenesis of Repetitive Mild Traumatic Brain Injury and Alzheimer’s Disease. Front. Neurosci. 2019, 13, 103. [Google Scholar] [CrossRef] [Green Version]
- Patel, D.; Witt, S.N. Ethanolamine and Phosphatidylethanolamine: Partners in Health and Disease. Oxidative Med. Cell. Longev. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaba, T.; Murate, M.; Tomishige, N.; Lee, Y.F.; Hullin-Matsuda, F.; Pollet, B.; Humbert, N.; Mély, Y.; Sako, Y.; Greimel, P.; et al. Formation of Tubules and Helical Ribbons by Ceramide Phosphoethanolamine-Containing Membranes. Sci. Rep. 2019, 9, 5812. Available online: https://www.nature.com/articles/s41598-019-42247-1 (accessed on 22 October 2021). [CrossRef] [PubMed] [Green Version]
- Hisano, K.; Yoshida, H.; Kawase, S.; Mimura, T.; Haniu, H.; Tsukahara, T.; Kurihara, T.; Matsuda, Y.; Saito, N.; Uemura, T. Abundant Oleoyl-Lysophosphatidylethanolamine in Brain Stimulates Neurite Outgrowth and Protects against Glutamate Toxicity in Cultured Cortical Neurons. J. Biochem. 2021, 170, 327–336. Available online: https://academic.oup.com/jb/article/170/3/327/6211200 (accessed on 22 October 2021). [CrossRef] [PubMed]
- Raghu, P.; Joseph, A.; Krishnan, H.; Singh, P.; Saha, S. Phosphoinositides: Regulators of Nervous System Function in Health and Disease. Front. Mol. Neurosci. 2019, 12, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javaid, S.; Farooq, T.; Rehman, Z.; Afzal, A.; Ashraf, W.; Rasool, M.F.; Alqahtani, F.; Alsanea, S.; Alasmari, F.; Alanazi, M.M.; et al. Dynamics of Choline-Containing Phospholipids in Traumatic Brain Injury and Associated Comorbidities. Int. J. Mol. Sci. 2021, 22, 11313. [Google Scholar] [CrossRef] [PubMed]
- Reed, T.T. Lipid Peroxidation and Neurodegenerative Disease. Free Radic. Biol. Med. 2011, 51, 1302–1319. Available online: https://pubmed.ncbi.nlm.nih.gov/21782935/ (accessed on 13 March 2022). [CrossRef] [PubMed]
- Gaschler, M.M.; Stockwell, B.R. Lipid Peroxidation in Cell Death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. Available online: https://pubmed.ncbi.nlm.nih.gov/28212725/ (accessed on 13 March 2022). [CrossRef] [PubMed]
- Calderon, R.O.; Attema, B.; DeVries, G.H. Lipid Composition of Neuronal Cell Bodies and Neurites from Cultured Dorsal Root Ganglia. J. Neurochem. 2002, 64, 424–429. Available online: https://pubmed.ncbi.nlm.nih.gov/7798942/ (accessed on 13 March 2022). [CrossRef]
- Gupte, R.P.; Brooks, W.; Vukas, R.; Pierce, J.D.; Harris, J.L. Sex Differences in Traumatic Brain Injury: What We Know and What We Should Know. J. Neurotrauma 2019, 36, 3063–3091. Available online: https://www.liebertpub.com/doi/abs/10.1089/neu.2018.6171 (accessed on 13 March 2022). [CrossRef]
- Brotfain, E.; Gruenbaum, S.E.; Boyko, M.; Kutz, R.; Zlotnik, A.; Klein, M. Neuroprotection by Estrogen and Progesterone in Traumatic Brain Injury and Spinal Cord Injury. Curr. Neuropharmacol. 2016, 14, 641–653. Available online: https://pubmed.ncbi.nlm.nih.gov/26955967/ (accessed on 13 March 2022). [CrossRef] [Green Version]
- Gray, N.; Lawler, N.; Zeng, A.; Ryan, M.; Bong, S.; Boughton, B.; Bizkarguenaga, M.; Bruzzone, C.; Embade, N.; Wist, J.; et al. Diagnostic Potential of the Plasma Lipidome in Infectious Disease: Application to Acute SARS-CoV-2 Infection. Metabolites 2021, 11, 467. Available online: https://www.mdpi.com/2218-1989/11/7/467/htm (accessed on 28 October 2021). [CrossRef]
- Adams, K.J.; Pratt, B.; Bose, N.; Dubois, L.G.; John-Williams, L.S.; Perrott, K.M.; Ky, K.; Kapahi, P.; Sharma, V.; MacCoss, M.J.; et al. Skyline for Small Molecules: A Unifying Software Package for Quantitative Metabolomics. J. Proteome Res. 2020, 19, 1447–1458. Available online: https://pubs.acs.org/doi/full/10.1021/acs.jproteome.9b00640 (accessed on 28 October 2021). [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Luan, H.; Ji, F.; Chen, Y.; Cai, Z. statTarget: A streamlined Tool for Signal Drift Correction and Interpretations of Quantitative Mass Spectrometry-Based Omics Data. Anal. Chim. Acta 2018, 1036, 66–72. Available online: https://europepmc.org/article/med/30253838 (accessed on 28 October 2021). [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Class | Lipid Species | Kruskal-Wallis | Post-Hoc Dunn’s Test | Control Mean (95% CI) | 1× mTBI Mean (95% CI) | 2× mTBI Mean (95% CI) | |||
|---|---|---|---|---|---|---|---|---|---|
| p | eta2 | p (Control vs. 1× mTBI) | p (Control vs. 2× mTBI) | p (1× mTBI vs. 2× mTBI) | |||||
| LPE | LPE(20:1) | 0.008 | 0.554 | 0.459 | 0.008 | 0.002 | 0.042 (0.030,0.054) | 0.042 (0.036,0.048) | 0.032 (0.028,0.036) |
| LPE | LPE(22:6) | 0.009 | 0.537 | 0.076 | 0.103 | 0.001 | 0.038 (0.022,0.054) | 0.049 (0.039,0.059) | 0.029 (0.023,0.035) |
| PI | PI(16:0/18:2) | 0.014 | 0.471 | 0.449 | 0.013 | 0.004 | 0.952 (0.806,1.098) | 1.050 (0.815,1.285) | 0.819 (0.777,0.861) |
| LPE | LPE(22:5) | 0.016 | 0.443 | 0.092 | 0.12 | 0.002 | 0.015 (0.007,0.023) | 0.021 (0.013,0.029) | 0.010 (0.006,0.014) |
| LPG | LPG(18:2) | 0.016 | 0.443 | 0.13 | 0.084 | 0.002 | 0.016 (0.011,0.021) | 0.018 (0.015,0.021) | 0.012 (0.009,0.015) |
| PS | PS(14:0/18:2) | 0.018 | 0.43 | 0.006 | 0.005 | 0.48 | 0.008 (0.005,0.011) | 0.004 (0.003,0.005) | 0.004 (0.002,0.006) |
| PE | PE(P-18:1/20:4) | 0.025 | 0.384 | 0.166 | 0.005 | 0.036 | 2.640 (1.771,3.511) | 2.990 (2.596,3.376) | 5.630 (2.151,9.101) |
| PI | PI(16:0/18:3) | 0.029 | 0.363 | 0.076 | 0.189 | 0.004 | 0.006 (0.005,0.007) | 0.009 (0.006,0.012) | 0.005 (0.004,0.006) |
| HCER | HCER(d18:0/26:0) | 0.032 | 0.348 | 0.015 | 0.006 | 0.387 | 0.001 (0.001,0.001) | 0.001 (0.001,0.001) | 0.001 (0.001,0.001) |
| FFA | FFA(20:3) | 0.038 | 0.324 | 0.005 | 0.057 | 0.118 | 0.712 (0.492,0.932) | 1.140 (0.738,1.532) | 0.954 (0.663,1.245) |
| PI | PI(18:1/18:2) | 0.039 | 0.319 | 0.429 | 0.03 | 0.01 | 0.454 (0.383,0.525) | 0.516 (0.341,0.691) | 0.361 (0.306,0.416) |
| HCER | HCER(22:0) | 0.04 | 0.317 | 0.469 | 0.021 | 0.013 | 1.550 (1.292,1.800) | 1.570 (1.431,1.715) | 1.820 (1.627,2.003) |
| CE | CE(14:0) | 0.041 | 0.312 | 0.043 | 0.006 | 0.198 | 1.490 (0.988,2.000) | 2.140 (1.475,2.805) | 2.370 (1.967,2.775) |
| FFA | FFA(18:3) | 0.049 | 0.288 | 0.054 | 0.322 | 0.008 | 6.930 (1.147,12.715) | 10.200 (7.815,12.607) | 5.940 (3.900,7.970) |
| PI | PI(20:0/18:2) | 0.049 | 0.287 | 0.399 | 0.039 | 0.011 | 0.030 (0.018,0.042) | 0.03 (0.025,0.035) | 0.024 (0.009,0.039) |
| PC | PC(20:0/22:4) | 0.049 | 0.287 | 0.399 | 0.039 | 0.011 | 0.050 (0.028,0.072) | 0.046 (0.039,0.053) | 0.061 (0.049,0.073) |
| PE | PE(P-18:0/20:1) | 0.049 | 0.288 | 0.159 | 0.009 | 0.064 | 0.075 (0.043,0.107) | 0.058 (0.048,0.068) | 0.049 (0.039,0.059) |
| PI | PI(18:1/18:1) | 0.05 | 0.286 | 0.237 | 0.081 | 0.008 | 0.498 (0.278,0.718) | 0.610 (0.371,0.849) | 0.385 (0.301,0.469) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anyaegbu, C.C.; Szemray, H.; Hellewell, S.C.; Lawler, N.G.; Leggett, K.; Bartlett, C.; Lins, B.; McGonigle, T.; Papini, M.; Anderton, R.S.; et al. Plasma Lipid Profiles Change with Increasing Numbers of Mild Traumatic Brain Injuries in Rats. Metabolites 2022, 12, 322. https://doi.org/10.3390/metabo12040322
Anyaegbu CC, Szemray H, Hellewell SC, Lawler NG, Leggett K, Bartlett C, Lins B, McGonigle T, Papini M, Anderton RS, et al. Plasma Lipid Profiles Change with Increasing Numbers of Mild Traumatic Brain Injuries in Rats. Metabolites. 2022; 12(4):322. https://doi.org/10.3390/metabo12040322
Chicago/Turabian StyleAnyaegbu, Chidozie C., Harrison Szemray, Sarah C. Hellewell, Nathan G. Lawler, Kerry Leggett, Carole Bartlett, Brittney Lins, Terence McGonigle, Melissa Papini, Ryan S. Anderton, and et al. 2022. "Plasma Lipid Profiles Change with Increasing Numbers of Mild Traumatic Brain Injuries in Rats" Metabolites 12, no. 4: 322. https://doi.org/10.3390/metabo12040322
APA StyleAnyaegbu, C. C., Szemray, H., Hellewell, S. C., Lawler, N. G., Leggett, K., Bartlett, C., Lins, B., McGonigle, T., Papini, M., Anderton, R. S., Whiley, L., & Fitzgerald, M. (2022). Plasma Lipid Profiles Change with Increasing Numbers of Mild Traumatic Brain Injuries in Rats. Metabolites, 12(4), 322. https://doi.org/10.3390/metabo12040322