
Untargeted Metabolomics Identify a Panel of Urinary Biomarkers for the Diagnosis of Urothelial Carcinoma of the Bladder, as Compared to Urolithiasis with or without Urinary Tract Infection in Dogs

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Subject Characteristics
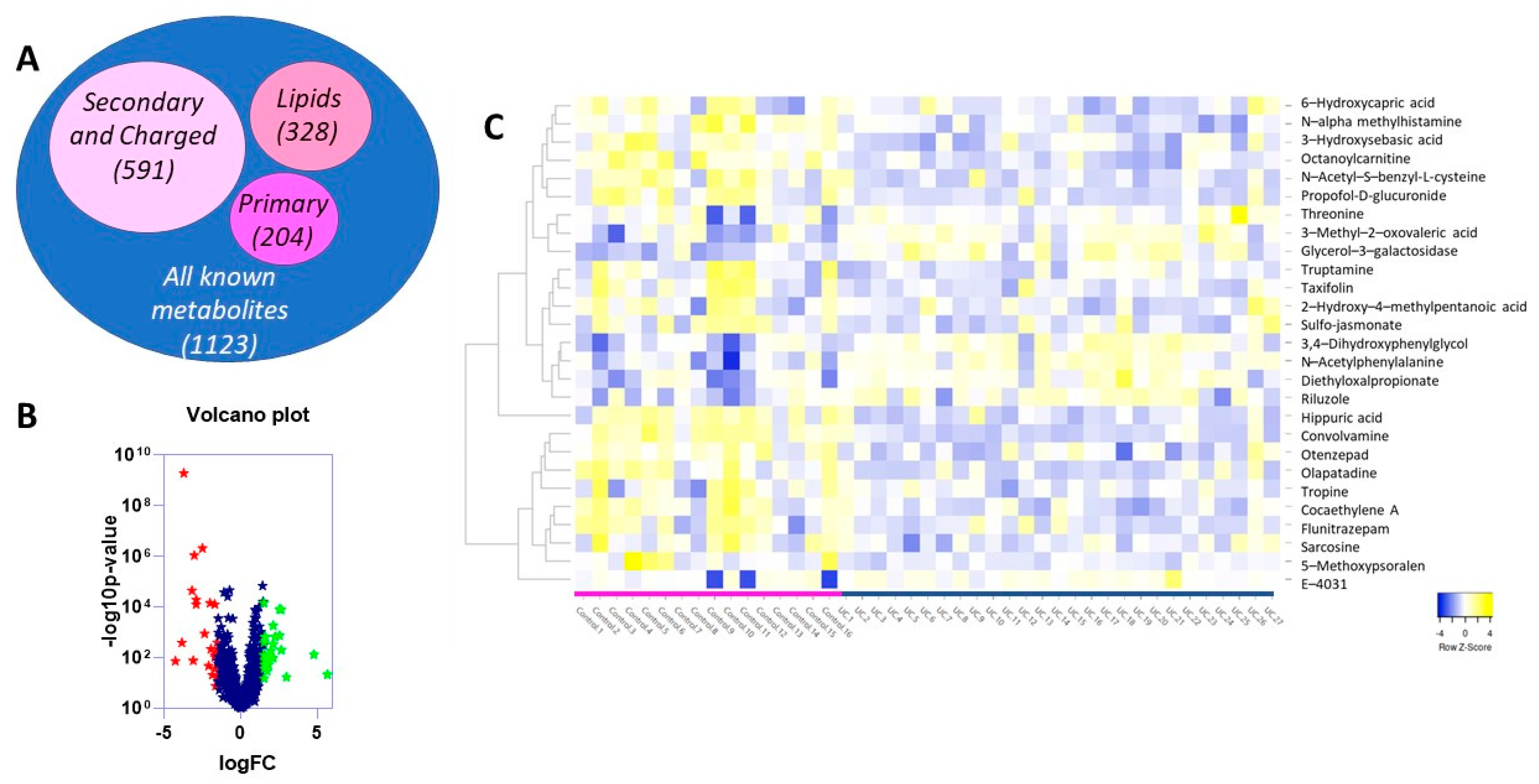
2.2. Differentially Expressed Metabolites in UC vs. Control Reflects Changes in Medication and Diet, as Well as Internal Genetic Alterations Due to Disease Progression
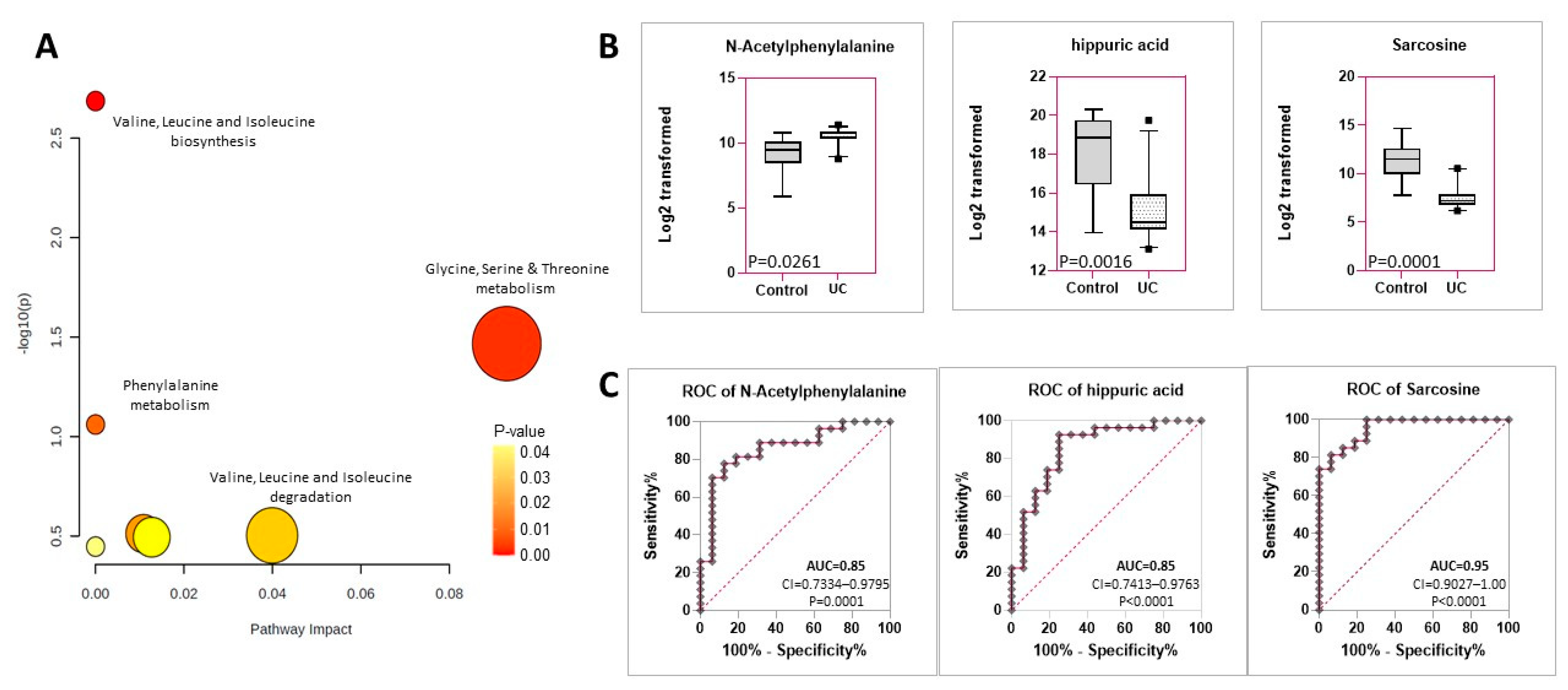
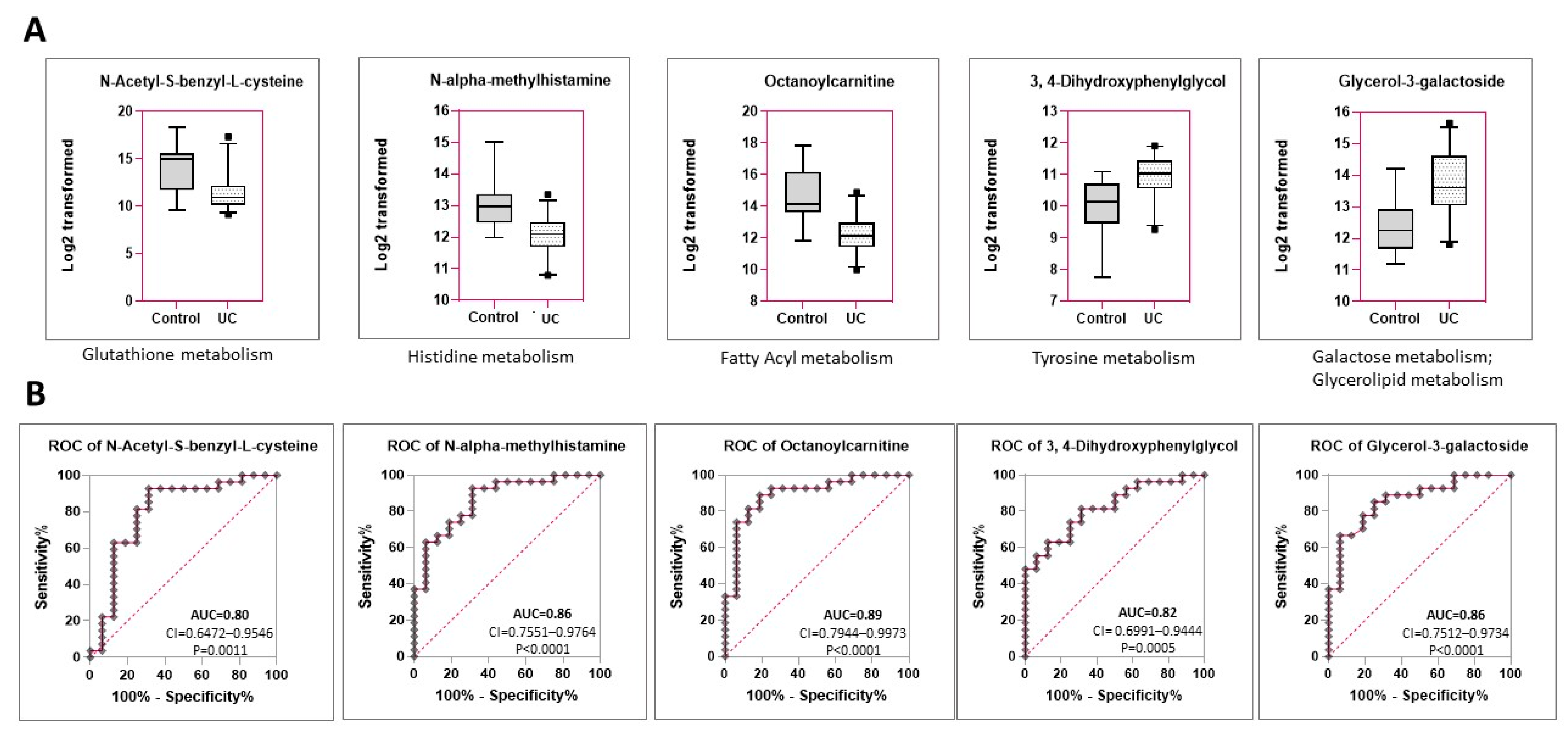
2.3. Identification of Six Metabolites That Are Distinctly Expressed in the Urine of Dogs with Urothelial Carcinoma vs. Urolithiasis with or without a UTI
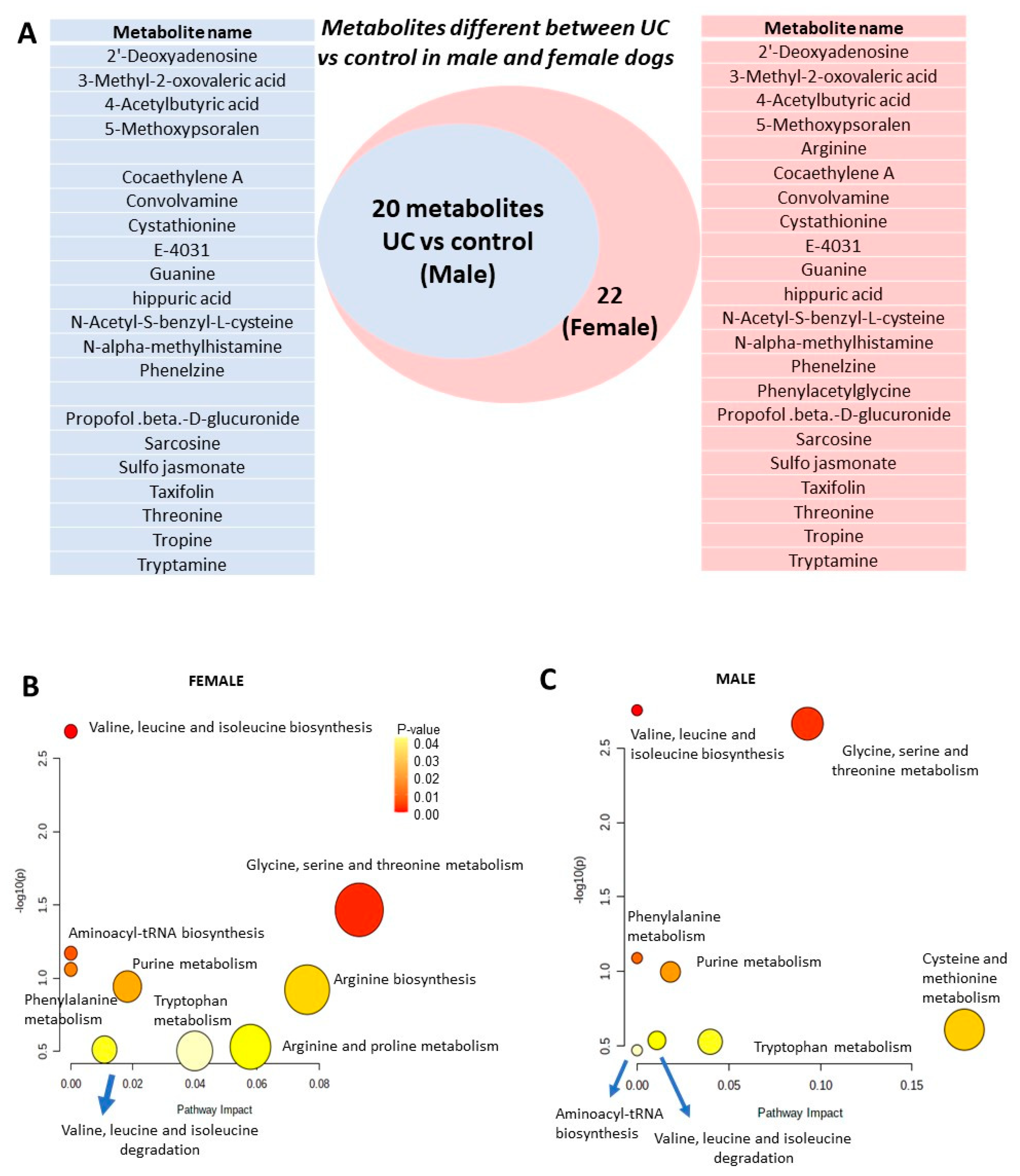
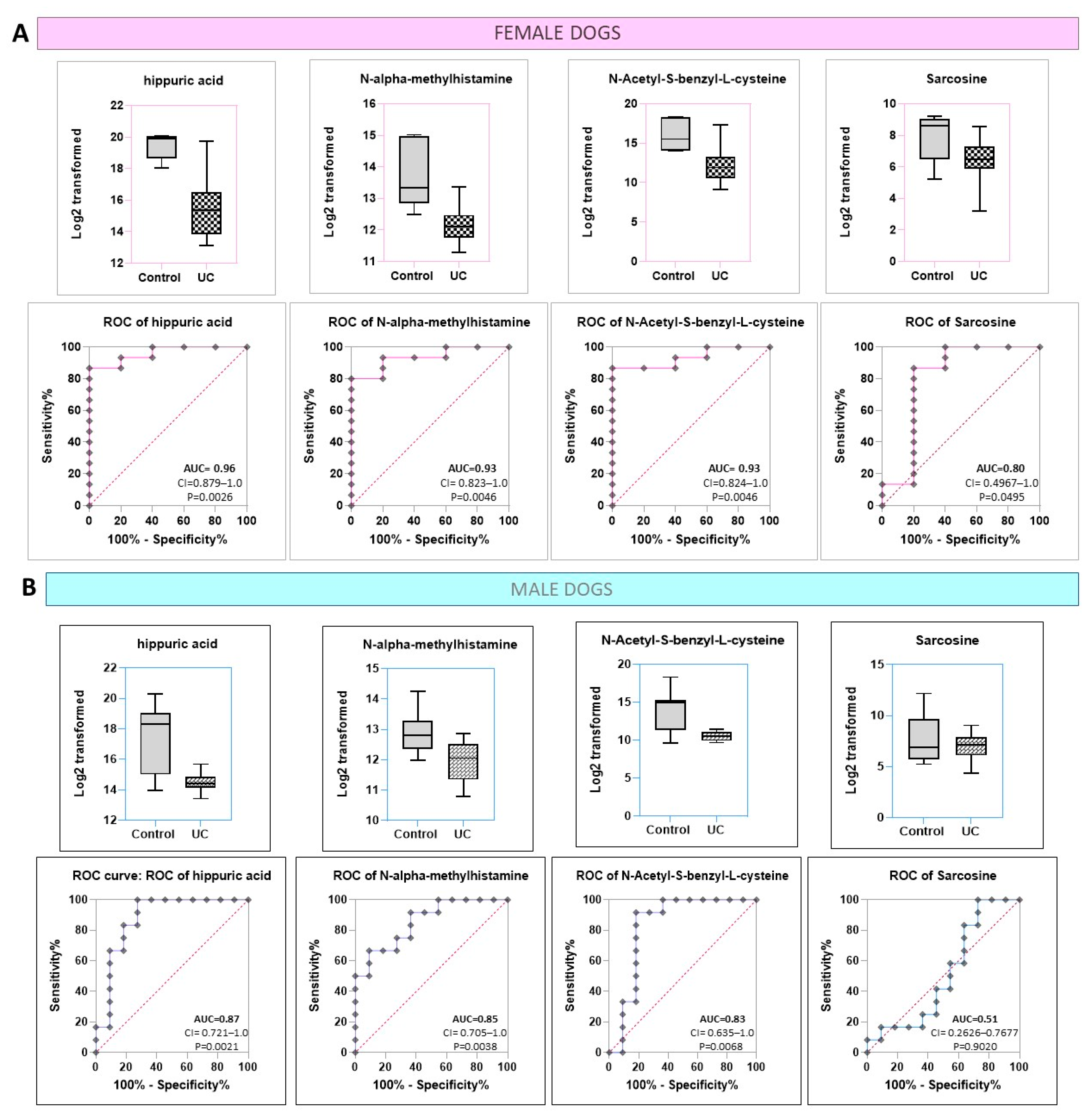
2.4. Hippuric Acid and NAMH Are Discriminatory between UC vs. Control in Both Male and Female Dogs
2.5. Breed-Related Differences in Metabolite Expression Show That Changes in Metabolites Observed Are Mostly Driven by Retrievers and Not Terriers
2.6. The Difference in These Metabolites Is Not Affected by Either NSAID or Antibiotic Use, or by Urethral Obstruction
3. Discussion
4. Materials and Methods
4.1. Urine Collection and Handling
4.2. Urinary Creatinine Levels
4.3. Metabolomic Analyses
- Primary metabolites: Using derivatization and GC-TOF MS, primary metabolites such as amino acids, sugars, and hydroxyl acids were profiled. The BinBase database used has over 1200 primary metabolites and the ability to annotate unknown metabolites [52].
- Complex lipids: >350 lipids covering 12 lipid classes (e.g., ceramides, cholesteryl esters, free fatty acids, phosphatidyl-cholines, -serines and -ethanolamines, lysophospholipids, sphingomyelins, and triacylglycerols) were quantified from an acetonitrile/isopropanol gradient on a charged surface hybrid (CSH) LC column and positive and negative electrospray ionization QTOF MS. The LipidBLAST library has 200,000 MS/MS lipid spectra.
- Secondary and charged metabolites: Charged, complex hydrophilic compounds such as cholines, nucleotides, nucleosides, methylated, and acetylated amines were measured using HILC QTOF MS; >100 secondary compounds were screened using the standards on this platform. Exposomes, such as metabolites from pharmaceutical drugs, were detected through untargeted analysis.
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fulkerson, C.M.; Knapp, D.W. Management of Transitional Cell Carcinoma of the Urinary Bladder in Dogs: A Review. Vet. J. 2015, 205, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Aupperle-Lellbach, H.; Grassinger, J.; Hohloch, C.; Kehl, A.; Pantke, P. Diagnostic value of the BRAF variant V595E in urine samples, smears and biopsies from canine transitional cell carcinoma. Tierarztl. Prax. Ausg. K Kleintiere Heimtiere 2018, 46, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, H.; Shapiro, S.G.; Breen, M. Detection of BRAF Mutation in Urine DNA as a Molecular Diagnostic for Canine Urothelial and Prostatic Carcinoma. PLoS ONE 2015, 10, e0144170. [Google Scholar] [CrossRef] [PubMed]
- Kent, M.S.; Zwingenberger, A.; Westropp, J.L.; Barrett, L.E.; Durbin-Johnson, B.P.; Ghosh, P.; Vinall, R.L. MicroRNA Profiling of Dogs with Transitional Cell Carcinoma of the Bladder Using Blood and Urine Samples. BMC Vet. Res. 2017, 13, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wei, S.; Liu, L.; Nagana Gowda, G.A.; Bonney, P.; Stewart, J.; Knapp, D.W.; Raftery, D. NMR-Based Metabolomics Study of Canine Bladder Cancer. Biochim. Biophys. Acta 2012, 1822, 1807–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiehn, O. Metabolomics—The Link between Genotypes and Phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Schmidt, D.R.; Patel, R.; Kirsch, D.G.; Lewis, C.A.; Vander Heiden, M.G.; Locasale, J.W. Metabolomics in Cancer Research and Emerging Applications in Clinical Oncology. Cancer J. Clin. 2021, 71, 333–358. [Google Scholar] [CrossRef]
- Dill, A.L.; Ifa, D.R.; Manicke, N.E.; Costa, A.B.; Ramos-Vara, J.A.; Knapp, D.W.; Cooks, R.G. Lipid Profiles of Canine Invasive Transitional Cell Carcinoma of the Urinary Bladder and Adjacent Normal Tissue by Desorption Electrospray Ionization Imaging Mass Spectrometry. Anal. Chem. 2009, 81, 8758–8764. [Google Scholar] [CrossRef] [Green Version]
- D’Hue, C.A.; Dhawan, D.; Peat, T.; Ramos-Vara, J.; Jarmusch, A.; Knapp, D.W.; Cooks, R.G. Fatty Acid Patterns Detected By Ambient Ionization Mass Spectrometry in Canine Invasive Urothelial Carcinoma From Dogs of Different Breeds. Bladder Cancer 2018, 4, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Rosenwasser, L.J.; O’Brien, T.; Weyne, J. Mast Cell Stabilization and Anti-Histamine Effects of Olopatadine Ophthalmic Solution: A Review of Pre-Clinical and Clinical Research. Curr. Med. Res. Opin. 2005, 21, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New Data Content and Improved Web Interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Sunil, C.; Xu, B. An Insight into the Health-Promoting Effects of Taxifolin (Dihydroquercetin). Phytochemistry 2019, 166, 112066. [Google Scholar] [CrossRef] [PubMed]
- Mitchelson, F.; Lanzafame, A.; Christopoulos, A. The Interaction of Otenzepad (AF-DX 116) with Allosteric Modulators and Competitive Antagonists at a Muscarinic M2 Receptor. Life Sci. 1999, 64, 559. [Google Scholar] [CrossRef]
- McNeely, W.; Goa, K.L. 5-Methoxypsoralen. A Review of Its Effects in Psoriasis and Vitiligo. Drugs 1998, 56, 667–690. [Google Scholar] [CrossRef]
- Balkrishna, A.; Thakur, P.; Varshney, A. Phytochemical Profile, Pharmacological Attributes and Medicinal Properties of Convolvulus Prostratus—A Cognitive Enhancer Herb for the Management of Neurodegenerative Etiologies. Front. Pharm. 2020, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.T.; Yuan, C.S.; Zhou, Z.; January, C.T. Enhancement of Delayed Afterdepolarizations and Triggered Activity by Class III Antiarrhythmic Drugs: Multiple Effects of E-4031 and Dofetilide. Methods Find Exp. Clin. Pharm. 2000, 22, 67–76. [Google Scholar] [CrossRef]
- Miller, R.G.; Mitchell, J.D.; Moore, D.H. Riluzole for Amyotrophic Lateral Sclerosis (ALS)/Motor Neuron Disease (MND). Cochrane Database Syst. Rev. 2012, 65, CD001447. [Google Scholar] [CrossRef]
- Andrews, P. Cocaethylene Toxicity. J. Addict. Dis. 1997, 16, 75–84. [Google Scholar] [CrossRef]
- Druid, H.; Holmgren, P.; Ahlner, J. Flunitrazepam: An Evaluation of Use, Abuse and Toxicity. Forensic Sci. Int. 2001, 122, 136–141. [Google Scholar] [CrossRef]
- Knapp, D.W.; Ramos-Vara, J.A.; Moore, G.E.; Dhawan, D.; Bonney, P.L.; Young, K.E. Urinary Bladder Cancer in Dogs, a Naturally Occurring Model for Cancer Biology and Drug Development. ILAR J. 2014, 55, 100–118. [Google Scholar] [CrossRef] [PubMed]
- Grassinger, J.M.; Merz, S.; Aupperle-Lellbach, H.; Erhard, H.; Klopfleisch, R. Correlation of BRAF Variant V595E, Breed, Histological Grade and Cyclooxygenase-2 Expression in Canine Transitional Cell Carcinomas. Vet. Sci. 2019, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drug-Induced Phospholipidosis-Pathological Aspects and Its Prediction. Available online: https://www.jstage.jst.go.jp/article/tox/21/1/21_1_9/_article/-char/ja/ (accessed on 9 January 2022).
- Pero, R.W. Health Consequences of Catabolic Synthesis of Hippuric Acid in Humans. Curr. Clin. Pharm. 2010, 5, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino Acids in Cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Rattray, N.J.W.; Zhang, Q.; Mironova, V.; Santos-Neto, A.; Muca, E.; Vollmar, A.K.R.; Hsu, K.-S.; Rattray, Z.; Cross, J.R.; et al. Tumor Tissue-Specific Biomarkers of Colorectal Cancer by Anatomic Location and Stage. Metabolites 2020, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-K.; Lin, C.-Y.; Kang, C.-J.; Liao, C.-T.; Wang, W.-L.; Chiang, M.-H.; Yen, T.-C.; Lin, G. Nuclear Magnetic Resonance Metabolomics Biomarkers for Identifying High Risk Patients with Extranodal Extension in Oral Squamous Cell Carcinoma. J. Clin. Med. 2020, 9, E951. [Google Scholar] [CrossRef] [Green Version]
- Pelayo, B.A.; Fu, Y.-M.; Meadows, G.G. Decreased Tissue Plasminogen Activator and Increased Plasminogen Activator Inhibitors and Increased Activator Protein-1 and Specific Promoter 1 Are Associated with Inhibition of Invasion in Human A375 Melanoma Deprived of Tyrosine and Phenylalanine. Int. J. Oncol. 2001, 18, 877–883. [Google Scholar] [CrossRef]
- Loras, A.; Martínez-Bisbal, M.C.; Quintás, G.; Gil, S.; Martínez-Máñez, R.; Ruiz-Cerdá, J.L. Urinary Metabolic Signatures Detect Recurrences in Non-Muscle Invasive Bladder Cancer. Cancers 2019, 11, 914. [Google Scholar] [CrossRef] [Green Version]
- Loras, A.; Trassierra, M.; Sanjuan-Herráez, D.; Martínez-Bisbal, M.C.; Castell, J.V.; Quintás, G.; Ruiz-Cerdá, J.L. Bladder Cancer Recurrence Surveillance by Urine Metabolomics Analysis. Sci. Rep. 2018, 8, 9172. [Google Scholar] [CrossRef] [Green Version]
- Łuczykowski, K.; Warmuzińska, N.; Operacz, S.; Stryjak, I.; Bogusiewicz, J.; Jacyna, J.; Wawrzyniak, R.; Struck-Lewicka, W.; Markuszewski, M.J.; Bojko, B. Metabolic Evaluation of Urine from Patients Diagnosed with High Grade (HG) Bladder Cancer by SPME-LC-MS Method. Molecules 2021, 26, 2194. [Google Scholar] [CrossRef]
- De Simone, G.; Balducci, C.; Forloni, G.; Pastorelli, R.; Brunelli, L. Hippuric Acid: Could Became a Barometer for Frailty and Geriatric Syndromes? Ageing Res. Rev. 2021, 72, 101466. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, S.S.; Gutzow, I.S. Hippuric Acid as a Significant Regulator of Supersaturation in Calcium Oxalate Lithiasis: The Physiological Evidence. BioMed Res. Int. 2013, 2013, 374950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amelio, I.; Cutruzzolá, F.; Antonov, A.; Agostini, M.; Melino, G. Serine and Glycine Metabolism in Cancer. Trends Biochem. Sci. 2014, 39, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, A.; Poisson, L.M.; Rajendiran, T.M.; Khan, A.P.; Cao, Q.; Yu, J.; Laxman, B.; Mehra, R.; Lonigro, R.J.; Li, Y.; et al. Metabolomic Profiles Delineate Potential Role for Sarcosine in Prostate Cancer Progression. Nature 2009, 457, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Walters, R.O.; Arias, E.; Diaz, A.; Burgos, E.S.; Guan, F.; Tiano, S.; Mao, K.; Green, C.L.; Qiu, Y.; Shah, H.; et al. Sarcosine Is Uniquely Modulated by Aging and Dietary Restriction in Rodents and Humans. Cell Rep. 2018, 25, 663–676.e6. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K. Targeting of NMDA Receptors in New Treatments for Schizophrenia. Expert Opin. Ther. Targets 2014, 18, 1049–1063. [Google Scholar] [CrossRef]
- Efficacy and Cognitive Effect of Sarcosine (N-Methylglycine) in Patients with Schizophrenia: A Systematic Review and Meta-Analysis of Double-Blind Randomised Controlled Trials. Available online: https://pubmed.ncbi.nlm.nih.gov/32122256/ (accessed on 9 January 2022).
- Saitoh, T.; Fukushima, Y.; Otsuka, H.; Ishikawa, M.; Tamai, M.; Takahashi, H.; Mori, H.; Asano, T.; Anai, M.; Ishikawa, T.; et al. Effects of N-Alpha-Methyl-Histamine on Human H(2) Receptors Expressed in CHO Cells. Gut 2002, 50, 786–789. [Google Scholar] [CrossRef] [Green Version]
- Millán-Guerrero, R.O.; Isais-Millán, R.; Guzmán-Chávez, B.; Castillo-Varela, G. N Alpha Methyl Histamine versus Propranolol in Migraine Prophylaxis. Can. J. Neurol. Sci. 2014, 41, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Simsekoglu, M.F.; Kaleler, I.; Onal, B.; Demirdag, C.; Citgez, S.; Uslu, E.; Erozenci, A.; Talat, Z. Do Urinary Mast Cell Mediators Predict Immune Response to BCG in Patients with Primary High-Grade Non-Muscle Invasive Bladder Cancer? Int. J. Clin. Pract. 2021, 75, e13959. [Google Scholar] [CrossRef]
- Dospinescu, V.-M.; Tiele, A.; Covington, J.A. Sniffing Out Urinary Tract Infection—Diagnosis Based on Volatile Organic Compounds and Smell Profile. Biosensors 2020, 10, 83. [Google Scholar] [CrossRef]
- Molendijk, J.; Robinson, H.; Djuric, Z.; Hill, M.M. Lipid Mechanisms in Hallmarks of Cancer. Mol. Omics 2020, 16, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Mounier, C.; Bouraoui, L.; Rassart, E. Lipogenesis in Cancer Progression (Review). Int. J. Oncol. 2014, 45, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prentki, M.; Madiraju, S.R.M. Glycerolipid Metabolism and Signaling in Health and Disease. Endocr. Rev. 2008, 29, 647–676. [Google Scholar] [CrossRef] [Green Version]
- Octanoylcarnitine—Bioblast. Available online: https://www.mitofit.org/index.php/Octanoylcarnitine (accessed on 21 November 2021).
- Schönfeld, P.; Wojtczak, L. Short- and Medium-Chain Fatty Acids in Energy Metabolism: The Cellular Perspective. J. Lipid. Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V. Cellular Fatty Acid Metabolism and Cancer. Cell Metab. 2013, 18, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Shin, Y.; Kim, T.H.; Kim, D.-H.; Lee, A. Plasma Metabolites as Possible Biomarkers for Diagnosis of Breast Cancer. PLoS ONE 2019, 14, e0225129. [Google Scholar] [CrossRef]
- Kind, T.; Cho, E.; Park, T.D.; Deng, N.; Liu, Z.; Lee, T.; Fiehn, O.; Kim, J. Interstitial Cystitis-Associated Urinary Metabolites Identified by Mass-Spectrometry Based Metabolomics Analysis. Sci. Rep. 2016, 6, 39227. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, B.M.; Stirdivant, S.M.; Mitchell, M.W.; Wulff, J.E.; McDunn, J.E.; Li, Z.; Dennis-Barrie, A.; Neri, B.P.; Milburn, M.V.; Lotan, Y.; et al. Bladder Cancer Biomarker Discovery Using Global Metabolomic Profiling of Urine. PLoS ONE 2014, 9, e115870. [Google Scholar] [CrossRef]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass Spectral and Retention Index Libraries for Metabolomics Based on Quadrupole and Time-of-Flight Gas Chromatography/Mass Spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-Enabled Heat Mapping for All. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N.; et al. HMDB 4.0: The Human Metabolome Database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UC (n = 27) | Urolithiasis Only (n = 8) | UTI and Urolithiasis (n = 8) | All Subjects (n = 43) | p-Value † | |
|---|---|---|---|---|---|
| Age | <0.001 | ||||
| Mean (SD) | 10.9 (1.9) | 7.2 (3.9) | 6.2 (4.6) | 9.3 (3.5) | |
| Median (Range) | 11 (6–14) | 6.5 (1–13) | 7 (0.3–12) | 10 (0.3–14) | |
| Gender | 0.00921 | ||||
| Female | 15 (55.6%) | 0 | 5 (62.5%) | 20 (46.5%) | |
| Male | 12 (44.4%) | 8 (100%) | 3 (37.5%) | 23 (53.5%) | |
| Breed | |||||
| Terrier | 5 (18.5%) | 4 (50%) | 3 (37.5%) | 12 (27.9%) | 0.20218 |
| Shepherd | 5 (18.5%) | 0 | 0 | 5 (11.6%) | 0.34365 |
| Retriever | 5 (18.5%) | 0 | 1 (12.5%) | 6 (14%) | 0.60082 |
| Others | 12 (44.4%) | 4 (50%) | 4 (50%) | 20 (46.5%) | |
| Conditions | |||||
| Castrated/Spayed | 25 (92.6%) | 6 (75%) | 6 (75%) | 37 (86%) | 0.17918 |
| Urethral obstruction | 5 (18.5%) | 1 (12.5%) | 1 (12.5%) | 7 (16.3%) | 1.00000 |
| Presence of metastasis | 3 (11.1%) | 0 | 0 | 3 (7%) | |
| Treatments | |||||
| Antibiotics | 7 (25.9%) | 0 | 1 (12.5%) | 8 (18.6%) | 0.30230 |
| NSAIDS | 12 (44.4%) | 3 (37.5%) | 0 | 15 (34.9%) | 0.07110 |
| Chemotherapy | 4 (14.8%) | 0 | 0 | 4 (9.3%) |
| Platform | Metabolite Name | logFC | AveExpr | p Value | adj.p.Val. |
|---|---|---|---|---|---|
| Primary metabolites (GCTOF MS) | glycerol-3-galactoside | 1.6722 | 13.4648 | 0.0006 | 0.0478 |
| hippuric acid | −3.5546 | 15.755 | <0.0001 | 0.0016 | |
| Secondary and charged metabolites (HILIC-QTOF MS) | E-4031 | 4.4226 | 6.8263 | 0.0014 | 0.0314 |
| Riluzole | 2.3073 | 10.9453 | 0.0013 | 0.0305 | |
| Diethyloxalpropionate | 1.8298 | 9.8314 | 0.0018 | 0.0387 | |
| 3-Methyl-2-oxovaleric acid | 1.8166 | 11.3829 | <0.0001 | 0.0022 | |
| Threonine | 1.8112 | 11.5928 | 0.0017 | 0.0378 | |
| N-Acetylphenylalanine | 1.6295 | 10.0706 | 0.001 | 0.0261 | |
| 3,4-Dihydroxyphenylglycol | 1.585 | 10.7024 | 0.0002 | 0.0111 | |
| N-alpha-methylhistamine | −1.5784 | 12.378 | 0.0001 | 0.0084 | |
| 3-Hydroxysebacic acid | −1.8382 | 11.246 | 0.0007 | 0.0217 | |
| 5-Methoxypsoralen | −1.9007 | 6.5929 | 0.0009 | 0.026 | |
| 6-Hydroxycaproic acid | −2.1693 | 12.4207 | 0.0005 | 0.0194 | |
| Octanoylcarnitine | −2.2014 | 12.6927 | 0.0002 | 0.0111 | |
| Otenzepad | −2.2737 | 8.5251 | 0.0007 | 0.0217 | |
| 2-Hydroxy-4-methylpentanoic acid | −2.3568 | 9.71 | 0.0005 | 0.0194 | |
| Taxifolin | −2.3879 | 9.4181 | 0.0004 | 0.0186 | |
| Sulfo jasmonate | −2.6733 | 10.5836 | 0.0008 | 0.0253 | |
| Tryptamine | −2.8279 | 9.4239 | 0.0001 | 0.0097 | |
| Flunitrazepam | −2.9725 | 7.2548 | 0.0002 | 0.0097 | |
| Sarcosine | −3.3428 | 7.0257 | <0.0001 | 0.0001 | |
| Cocaethylene A | −3.3942 | 7.7349 | <0.0001 | 0.0002 | |
| Propofol. beta.-D-glucuronide | −3.6922 | 11.0599 | 0.0002 | 0.0097 | |
| N-Acetyl-S-benzyl-L-cysteine | −3.8876 | 12.156 | 0.0001 | 0.0063 | |
| Olopatadine | −3.8901 | 8.6456 | 0.0002 | 0.0097 | |
| Convolvamine | −4.0141 | 8.3401 | <0.0001 | 0.0001 | |
| Tropine | −4.1392 | 9.54 | 0.0001 | 0.0084 |
| Pathway Name | Metabolites | p Value |
|---|---|---|
| Valine, Leucine and Isoleucine Biosynthesis | L-Threonine, 3-Methyl-2-Oxovaleric acid, 2-hydroxy-4-methylpentanoic acid | 0.00103 |
| Glycine, Serine and Threonine Metabolism | Sarcosine, L-Threonine | 0.00178 |
| Phenylalanine Metabolism | Hippuric acid, N-Acetylphenylalanine, 5-Methoxypsoralen, Taxifolin | 0.00629 |
| Valine, Leucine, and Isoleucine Degradation | 3-Methyl-2-Oxovaleric acid, L-Threonine | 0.231 |
| Tryptophan Metabolism | Tryptamine | 0.236 |
| Tyrosine Metabolism | 3-4-Dihydroxyphenylglycol | 0.241 |
| Aminoacyl-tRNA Biosynthesis | L-Threonine | 0.271 |
| Group | Breed | Defined by | UC (n = 27) | U/UTI (n = 16) |
|---|---|---|---|---|
| Herding | Australian Cattle Dog † | AKC | 1 | 0 |
| Australian Shepherd † | AKC | 2 | 0 | |
| Border Collie † | AKC | 1 | 0 | |
| Collie Mix | AKC | 1 | 0 | |
| German Shepherd | AKC | 1 | 0 | |
| Shepherd Mix | AKC | 2 | 0 | |
| Shetland Sheepdog † | AKC | 1 | 0 | |
| Hound | Beagle † | AKC | 1 | 0 |
| Dachshund | AKC | 1 | 0 | |
| Rhodesian Ridgeback | AKC | 1 | 0 | |
| Non-sporting | Bichon Frise † | AKC | 0 | 2 |
| Dalmatian | AKC | 1 | 0 | |
| French Bulldog | AKC | 0 | 1 | |
| Sporting | Brittany/Spaniel mix | AKC | 0 | 1 |
| Golden Retriever | AKC | 2 | 1 | |
| Labrador Retriever | AKC | 2 | 0 | |
| Labrador Retriever Mix | AKC | 1 | 0 | |
| Terrier | American Pit Bull Terrier | UKC | 0 | 1 |
| Miniature Schnauzer | AKC | 1 | 2 | |
| Terrier Mix | AKC | 1 | 3 | |
| West Highland White Terrier † | AKC | 3 | 1 | |
| Yorkshire Terrier | AKC | 0 | 1 | |
| Companion | Chihuahua mix | AKC | 0 | 1 |
| Miniature Pinscher | AKC | 2 | 0 | |
| Pomeranian | AKC | 1 | 0 | |
| Working | Great Pyrenees | AKC | 0 | 1 |
| Mastiff | AKC | 1 | 0 | |
| Rottweiler | AKC | 0 | 1 |
| Terrier vs. Non-Terrier | ||||||||
|---|---|---|---|---|---|---|---|---|
| UC | Control | |||||||
| Metabolite Name | logFC | AveExpr | p Value | adj.p.Val. | logFC | AveExpr | p Value | adj.p.Val. |
| Taxifolin | 3.32344 | 9.4180 | 6.57 × 10−5 | 0.0191 | 3.2452 | 9.4180 | 0.0001 | 0.0376 |
| 3-Hydroxykynurenine | −7.8520 | 13.8756 | 7.22 × 10−5 | 0.0191 | −7.7438 | 13.8756 | 0.0001 | 0.0376 |
| 4-Pyridoxic acid | −2.2640 | 12.0144 | 9.73 × 10−5 | 0.0191 | −2.2118 | 12.0144 | 0.0001 | 0.0376 |
| Homoarginine | 2.4386 | 8.33970 | 0.0002 | 0.0358 | ||||
| Tryptamine | 3.2555 | 9.4239 | 0.0003 | 0.0359 | ||||
| Flunitrazepam | 3.3556 | 7.2548 | 0.0004 | 0.0359 | ||||
| 2-[(4-Aminobenzoyl) amino]acetic acid | −2.4797 | 13.9770 | 0.0004 | 0.0359 | ||||
| N-Methylalanine | 3.35947 | 10.8914 | 0.0006 | 0.0461 | ||||
| Tropine | 4.2490 | 9.5399 | 0.0007 | 0.0468 | ||||
| Urethral Obstruction | No Urethral Obstruction | ||||||
|---|---|---|---|---|---|---|---|
| Metabolite Name | AveExpr | logFC | p Value | adj.p.Val | logFC | p Value | adj.p.Val |
| 2,3-Dihydroxybenzoic acid B | 10.63 | 2.59 | 0.0031 | 0.0473 | |||
| 2’-Deoxyadenosine | 9.38 | −1.52 | 0.0017 | 0.0341 | |||
| 2-Hydroxy-4-methylpentanoic acid | 9.71 | −2.11 | 0.0015 | 0.0319 | −2.45 | 0.0007 | 0.0218 |
| 3,4-Dihydroxyphenylglycol | 10.70 | 1.61 | 0.0004 | 0.0160 | |||
| 3-Hydroxysebacic acid | 11.25 | −1.92 | 0.0001 | 0.0064 | −2.08 | 0.0001 | 0.0057 |
| 3-Methyl-2-oxovaleric acid | 11.38 | 1.82 | 0.0000 | 0.0005 | 1.90 | 0.0000 | 0.0008 |
| 4-Acetylbutyric acid | 13.37 | 2.38 | 0.0009 | 0.0271 | 2.33 | 0.0022 | 0.0422 |
| 5-Methoxypsoralen | 6.59 | −1.83 | 0.0010 | 0.0271 | −2.05 | 0.0007 | 0.0218 |
| 6-Hydroxycaproic acid | 12.42 | −1.91 | 0.0015 | 0.0319 | −2.24 | 0.0006 | 0.0218 |
| Cer d36:1 | 9.78 | −2.09 | 0.0005 | 0.0472 | |||
| Cer d42:2 Isomer A | 10.95 | −1.64 | 0.0006 | 0.0472 | |||
| Cer-NS d34:1; Cer-NS d18:1/16:0; | 8.82 | −3.07 | 0.0005 | 0.0472 | |||
| Cocaethylene A | 7.73 | −3.19 | 0.0000 | 0.0005 | −3.45 | 0.0000 | 0.0006 |
| Convolvamine | 8.34 | −4.17 | 0.0000 | 0.0000 | −4.15 | 0.0000 | 0.0000 |
| D-Glucosamine | 9.50 | −1.63 | 0.0025 | 0.0424 | |||
| Diethyloxalpropionate | 9.83 | 1.88 | 0.0010 | 0.0271 | 1.99 | 0.0011 | 0.0255 |
| E-4031 | 6.83 | 4.48 | 0.0012 | 0.0294 | 4.70 | 0.0013 | 0.0307 |
| FA 24:1 (nervonic acid) | 10.66 | 1.72 | 0.0004 | 0.0472 | |||
| Flunitrazepam | 7.25 | −2.98 | 0.0001 | 0.0064 | −3.24 | 0.0001 | 0.0072 |
| glycerol-3-galactoside | 13.46 | 1.69 | 0.0003 | 0.0296 | 1.81 | 0.0003 | 0.0299 |
| Haloperidol | 7.69 | −3.14 | 0.0026 | 0.0424 | |||
| hippuric acid | 15.75 | −3.68 | 0.0000 | 0.0003 | −3.72 | 0.0000 | 0.0009 |
| Lobelanidine | 9.60 | −1.52 | 0.0016 | 0.0321 | −1.69 | 0.0011 | 0.0255 |
| Metanephrine | 10.03 | −1.62 | 0.0021 | 0.0393 | |||
| N-Acetylphenylalanine | 10.07 | 1.72 | 0.0003 | 0.0138 | 1.77 | 0.0005 | 0.0196 |
| N-Acetyl-S-benzyl-L-cysteine | 12.16 | −3.96 | 0.0000 | 0.0020 | −4.12 | 0.0000 | 0.0031 |
| N-alpha-methylhistamine | 12.38 | −1.52 | 0.0001 | 0.0064 | −1.54 | 0.0003 | 0.0111 |
| N-Methylalanine | 10.89 | −2.15 | 0.0019 | 0.0376 | −2.27 | 0.0022 | 0.0422 |
| Octanoylcarnitine | 12.69 | −2.22 | 0.0001 | 0.0064 | −2.32 | 0.0002 | 0.0077 |
| Olopatadine | 8.65 | −3.95 | 0.0001 | 0.0064 | −4.35 | 0.0001 | 0.0047 |
| Otenzepad | 8.53 | −2.22 | 0.0007 | 0.0227 | −2.33 | 0.0008 | 0.0225 |
| Palatinose | 11.85 | 3.38 | 0.0023 | 0.0418 | |||
| Phenylacetylglycine | 12.62 | 1.50 | 0.0008 | 0.0225 | |||
| Propofol. beta.-D-glucuronide | 11.06 | −4.03 | 0.0000 | 0.0010 | −4.24 | 0.0000 | 0.0014 |
| Riluzole | 10.95 | 2.25 | 0.0026 | 0.0487 | |||
| Sarcosine | 7.03 | −3.22 | 0.0000 | 0.0001 | −3.46 | 0.0000 | 0.0001 |
| Sulfo jasmonate | 10.58 | −2.64 | 0.0008 | 0.0246 | −2.81 | 0.0008 | 0.0225 |
| Taxifolin | 9.42 | −2.49 | 0.0001 | 0.0064 | −2.68 | 0.0001 | 0.0072 |
| Tropine | 9.54 | −4.28 | 0.0000 | 0.0012 | −4.41 | 0.0000 | 0.0021 |
| Tryptamine | 9.42 | −2.74 | 0.0002 | 0.0069 | −2.92 | 0.0002 | 0.0077 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsamouri, M.M.; Durbin-Johnson, B.P.; Culp, W.T.N.; Palm, C.A.; Parikh, M.; Kent, M.S.; Ghosh, P.M. Untargeted Metabolomics Identify a Panel of Urinary Biomarkers for the Diagnosis of Urothelial Carcinoma of the Bladder, as Compared to Urolithiasis with or without Urinary Tract Infection in Dogs. Metabolites 2022, 12, 200. https://doi.org/10.3390/metabo12030200
Tsamouri MM, Durbin-Johnson BP, Culp WTN, Palm CA, Parikh M, Kent MS, Ghosh PM. Untargeted Metabolomics Identify a Panel of Urinary Biomarkers for the Diagnosis of Urothelial Carcinoma of the Bladder, as Compared to Urolithiasis with or without Urinary Tract Infection in Dogs. Metabolites. 2022; 12(3):200. https://doi.org/10.3390/metabo12030200
Chicago/Turabian StyleTsamouri, Maria Malvina, Blythe P. Durbin-Johnson, William T. N. Culp, Carrie A. Palm, Mamta Parikh, Michael S. Kent, and Paramita M. Ghosh. 2022. "Untargeted Metabolomics Identify a Panel of Urinary Biomarkers for the Diagnosis of Urothelial Carcinoma of the Bladder, as Compared to Urolithiasis with or without Urinary Tract Infection in Dogs" Metabolites 12, no. 3: 200. https://doi.org/10.3390/metabo12030200
APA StyleTsamouri, M. M., Durbin-Johnson, B. P., Culp, W. T. N., Palm, C. A., Parikh, M., Kent, M. S., & Ghosh, P. M. (2022). Untargeted Metabolomics Identify a Panel of Urinary Biomarkers for the Diagnosis of Urothelial Carcinoma of the Bladder, as Compared to Urolithiasis with or without Urinary Tract Infection in Dogs. Metabolites, 12(3), 200. https://doi.org/10.3390/metabo12030200