
Structural Diversity, LC-MS-MS Analysis and Potential Biological Activities of Brevibacillus laterosporus Extract

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation Conditions of Halotolerant Bacteria
2.2. Antibacterial Potencies against Pathogenic Bacteria
2.2.1. Paper Disc Assay
2.2.2. Agar Well Diffusion Assay
2.3. Molecular Identification of the Isolated Bacteria
2.4. Optimizing Culture Conditions of the B. laterosporus BF202 Antibacterial Activity
2.4.1. Different Types of Growth Media
2.4.2. Different Incubation Periods
2.4.3. Different pH Values
2.4.4. Different Temperature Degrees
2.4.5. Miscellaneous Carbon and Nitrogen Sources
2.5. Method for Preparing the Crude Extract
2.6. Characterization of Methanol Extract Using LC-MS/MS and GNPS
2.7. In Vitro Anticancer Study
2.7.1. Maintenance of Cell Lines
2.7.2. Cytotoxicity Assay
2.7.3. Characterization of Apoptosis Using Annexin V/Propidium Iodide (PI) Labeling
2.7.4. Cell Cycle Analysis Using Flow Cytometer
3. Results
3.1. Isolation of Halotolerant Bacteria and the Estimation of Their Antibacterial Activities
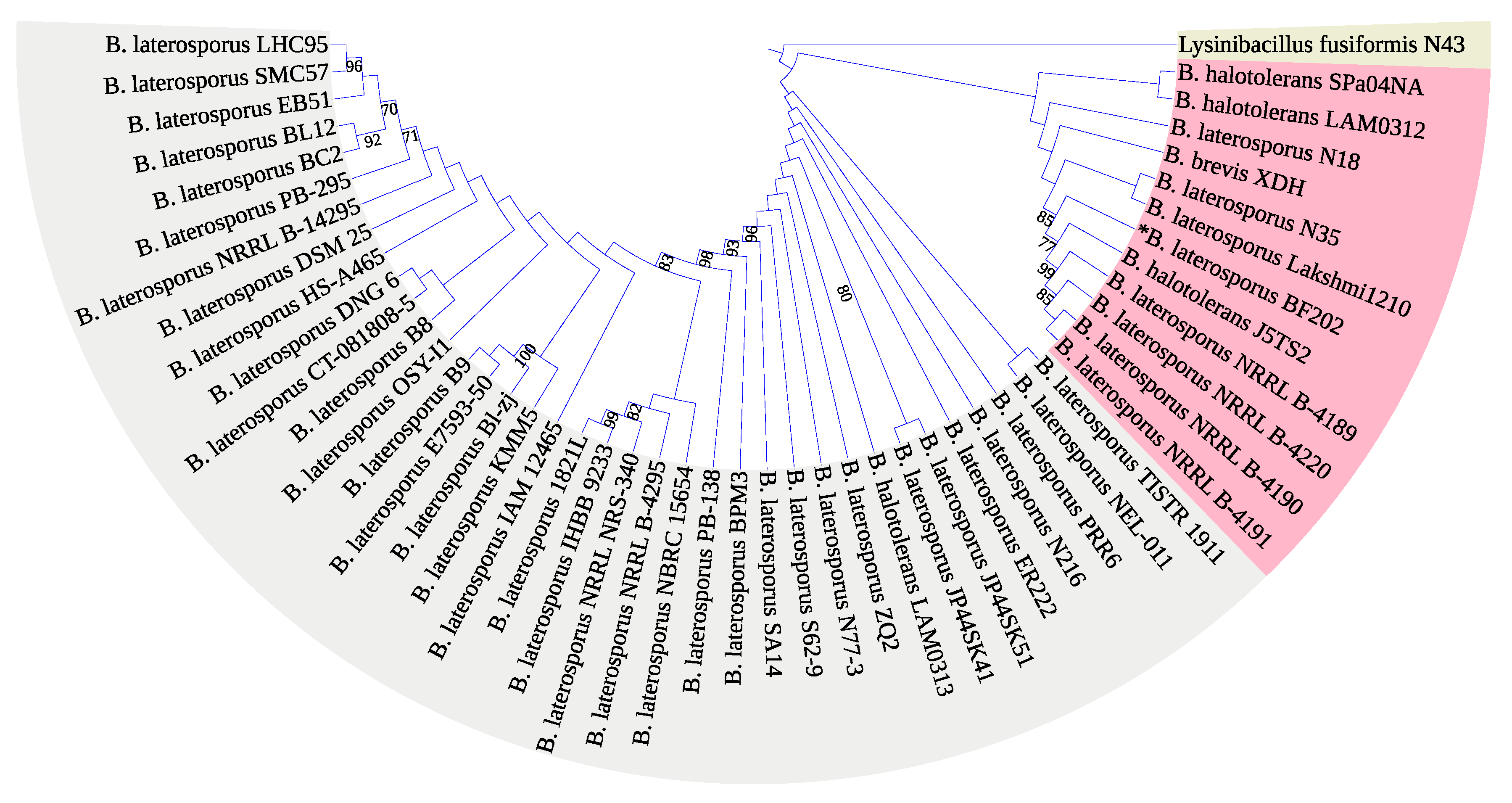
3.2. Molecular Identification of the Bacterial Isolate
3.3. Optimization of the Culture Conditions of the B. laterosporus BF202
3.4. LC-MS/MS Analysis of B. laterosporus Bioactive Extract
3.5. Cytotoxicity of the B. laterosporus Crude Extract
3.6. Quantification of Apoptosis
3.7. Cell Cycle Analysis Using Flow Cytometer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deslouches, B.; Di, Y.P. Antimicrobial Peptides with Selective Antitumor Mechanisms: Prospect for Anticancer Applications. Oncotarget 2017, 8, 46635. [Google Scholar] [CrossRef] [PubMed]
- Kamenshchikova, A.; Wolffs, P.F.G.; Hoebe, C.; Horstman, K. Anthropocentric Framings of One Health: An Analysis of International Antimicrobial Resistance Policy Documents. Crit. Public Health 2021, 31, 306–315. [Google Scholar] [CrossRef]
- Eric, N.; Janet, L.; Steven, K.G. Inflammation, Immune Activation, and Cardiovascular Disease in HIV. AIDS 2016, 30, 1495. [Google Scholar]
- Hale, J.D.F.; Hancock, R.E.W. Alternative Mechanisms of Action of Cationic Antimicrobial Peptides on Bacteria. Expert Rev. Anti Infect. Ther. 2007, 5, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Vogelmann, R.; Amieva, M.R. The Role of Bacterial Pathogens in Cancer. Curr. Opin. Microbiol. 2007, 10, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Alipour, M. Molecular Mechanism of Helicobacter Pylori-Induced Gastric Cancer. J. Gastrointest. Cancer 2021, 52, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Burns, J.L.; Montelaro, R.C. Engineered Cationic Antimicrobial Peptides to Overcome Multidrug Resistance by ESKAPE Pathogens. Antimicrob. Agents Chemother. 2015, 59, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Siegal, R.; Miller, K.D.; Jemal, A. Cancer Statistics, 2012. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Ting, C.-H.; Huang, H.-N.; Huang, T.-C.; Wu, C.-J.; Chen, J.-Y. The Mechanisms by Which Pardaxin, a Natural Cationic Antimicrobial Peptide, Targets the Endoplasmic Reticulum and Induces c-FOS. Biomaterials 2014, 35, 3627–3640. [Google Scholar] [CrossRef]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Figueiredo, C.R.; Matsuo, A.L.; Massaoka, M.H.; Polonelli, L.; Travassos, L.R. Anti-Tumor Activities of Peptides Corresponding to Conserved Complementary Determining Regions from Different Immunoglobulins. Peptides 2014, 59, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Krachkovskii, S.A.; Sobol, A.G.; Ovchinnikova, T.V.; Tagaev, A.A.; Yakimenko, Z.A.; Azizbekyan, R.R.; Kuznetsova, N.I.; Shamshina, T.N.; Arseniev, A.S. Isolation, Biological Properties, and Spatial Structure of Antibiotic Loloatin A. Russ. J. Bioorganic Chem. 2002, 28, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, L.; Liu, Y.; Han, P.; Hong, D.; Li, S.; Ma, A.; Jia, Y. Brevilaterin B from Brevibacillus Laterosporus Has Selective Antitumor Activity and Induces Apoptosis in Epidermal Cancer. World J. Microbiol. Biotechnol. 2022, 38, 201. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, T.; Senthil Kumar, P.; Hemavathy, R.V.; Swetha, V.; Karishma Sri, R. Isolation, Structure Elucidation and Anticancer Activity from Brevibacillus Brevis EGS 9 That Combats Multi Drug Resistant Actinobacteria. Microb. Pathog. 2018, 115, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ji, C.; Sui, J.; Sa, R.; Wang, X.; Liu, X.; Guo, T.L. Antibacterial and Antitumor Activity of Bogorol B-JX Isolated from Brevibacillus Laterosporus JX-5. World J. Microbiol. Biotechnol. 2017, 33, 177. [Google Scholar] [CrossRef]
- Laubach, C.A. Studies On Aerobic Spore-Bearing Non-Pathogenic Bacteria Part II Spore-Bearing Bacteria In Dust. J. Bacteriol. 1916, 1, 493–533. [Google Scholar] [CrossRef] [PubMed]
- Shida, O.; Takagi, H.; Kadowaki, K.; Komagata, K. Proposal for Two New Genera, Brevibacillus Gen. Nov. and Aneurinibacillus Gen. Nov. Int. J. Syst. Bacteriol. 1996, 46, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Sharma, V.; Patil, P.B.; Korpole, S. Identification, Purification and Characterization of Laterosporulin, a Novel Bacteriocin Produced by Brevibacillus Sp. Strain GI-9. PLoS ONE 2012, 7, e31498. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, P.; Senthilkumar, P.K. On Overview of Saltpan Halophilic Bacterium. Antimicrob. Agents 2017, 3, 1–5. [Google Scholar]
- Vasavada, S.H.; Thumar, J.T.; Singh, S.P. Secretion of a Potent Antibiotic by Salt-Tolerant and Alkaliphilic Actinomycete Streptomyces Sannanensis Strain RJT-1. Curr. Sci. 2006, 91, 1393–1397. [Google Scholar]
- Chamekh, R.; Deniel, F.; Donot, C.; Jany, J.-L.; Nodet, P.; Belabid, L. Isolation, Identification and Enzymatic Activity of Halotolerant and Halophilic Fungi from the Great Sebkha of Oran in Northwestern of Algeria. Mycobiology 2019, 47, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Fariq, A.; Yasmin, A.; Jamil, M. Production, Characterization and Antimicrobial Activities of Bio-Pigments by Aquisalibacillus Elongatus MB592, Salinicoccus Sesuvii MB597, and Halomonas Aquamarina MB598 Isolated from Khewra Salt Range, Pakistan. Extremophiles 2019, 23, 435–449. [Google Scholar] [CrossRef]
- Mainka, T.; Weirathmüller, D.; Herwig, C.; Pflügl, S. Potential Applications of Halophilic Microorganisms for Biological Treatment of Industrial Process Brines Contaminated with Aromatics. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab015. [Google Scholar] [CrossRef]
- Mesbah, N.M.; Hedrick, D.B.; Peacock, A.D.; Rohde, M.; Wiegel, J. Natranaerobius Thermophilus Gen. Nov., Sp. Nov., a Halophilic, Alkalithermophilic Bacterium from Soda Lakes of the Wadi An Natrun, Egypt, and Proposal of Natranaerobiaceae Fam. Nov. and Natranaerobiales Ord. Nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2507–2512. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Krumbein, W.E.; Goldberg, M.; Shilo, M. Solar Lake (Sinai). 1. Physical and Chemical Limnology 1. Limnol. Oceanogr. 1977, 22, 597–608. [Google Scholar] [CrossRef]
- Asker, D.; Ohta, Y. Haloferax Alexandrinus Sp. Nov., an Extremely Halophilic Canthaxanthin-Producing Archaeon from a Solar Saltern in Alexandria (Egypt). Int. J. Syst. Evol. Microbiol. 2002, 52, 729–738. [Google Scholar] [PubMed]
- Eltarahony, M.; Ibrahim, A.; El-shall, H.; Ibrahim, E.; Althobaiti, F.; Fayad, E. Antibacterial, Antifungal and Antibiofilm Activities of Silver Nanoparticles Supported by Crude Bioactive Metabolites of Bionanofactories Isolated from Lake Mariout. Molecules 2021, 26, 3027. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, L.; Hong, D.; Liu, Y.; Han, P.; Li, S.; Jia, Y. Broad-spectrum Cytotoxicity to Cancer Cells of Brevilaterin C from Brevibacillus laterosporus and Its Specific Mechanism on Human Epidermal Cancer Cells. J. Cell. Biochem. 2022, 123, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- El-Garawani, I.M.; El-Sabbagh, S.M.; Abbas, N.H.; Ahmed, H.S.; Eissa, O.A.; Abo-Atya, D.M.; Khalifa, S.A.M.; El-Seedi, H.R. A Newly Isolated Strain of Halomonas Sp.(HA1) Exerts Anticancer Potential via Induction of Apoptosis and G2/M Arrest in Hepatocellular Carcinoma (HepG2) Cell Line. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.-Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the Oceans to the Potential Biotechnological and Biomedical Applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Yosri, N.; Khalifa, S.A.M.; Guo, Z.; Xu, B.; Zou, X.; El-Seedi, H.R. Marine Organisms: Pioneer Natural Sources of Polysaccharides/Proteins for Green Synthesis of Nanoparticles and Their Potential Applications. Int. J. Biol. Macromol. 2021, 193, 1767–1798. [Google Scholar] [CrossRef] [PubMed]
- Holmalahti, J.; von Wright, A.; Raatikainen, O. Variations in the Spectra of Biological Activities of Actinomycetes Isolated from Different Soils. Lett. Appl. Microbiol. 1994, 18, 144–146. [Google Scholar] [CrossRef]
- NCBI Resource Coordinators. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [PubMed]
- Zayed, M.; Badawi, M.A. In-Silico Evaluation of a New Gene from Wheat Reveals the Divergent Evolution of the CAP160 Homologous Genes Into Monocots. J. Mol. Evol. 2020, 88, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Pearson, W.R. An Introduction to Sequence Similarity (“Homology”) Searching. Curr. Protoc. Bioinforma. 2013, 42, 3.1.1–3.1.8. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v3: An Online Tool for the Display and Annotation of Phylogenetic and Other Trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Microbiological Media; CRC Press: Boca Raton, FL, USA, 2004; ISBN 0429129033. [Google Scholar]
- Rogosa, M.; Mitchell, J.A.; Wiseman, R.F. A Selective Medium for the Isolation and Enumeration of Oral and Fecal Lactobacilli. J. Bacteriol. 1951, 62, 132–133. [Google Scholar] [CrossRef] [PubMed]
- El-Garawani, I.M.; Hassab El-Nabi, S.; El Kattan, A.; Sallam, A.; Elballat, S.; Abou-Ghanima, S.; El Azab, I.H.; El-Seedi, H.R.; Khalifa, S.A.M.; El-Shamy, S. The Ameliorative Role of Acacia Senegal Gum against the Oxidative Stress and Genotoxicity Induced by the Radiographic Contrast Medium (Ioxitalamate) in Albino Rats. Antioxidants 2021, 10, 221. [Google Scholar] [CrossRef] [PubMed]
- El-Din, M.I.G.; Fahmy, N.M.; Wu, F.; Salem, M.M.; Khattab, O.M.; El-Seedi, H.R.; Korinek, M.; Hwang, T.-L.; Osman, A.K.; El-Shazly, M. Comparative LC–LTQ–MS–MS Analysis of the Leaf Extracts of Lantana Camara and Lantana Montevidensis Growing in Egypt with Insights into Their Antioxidant, Anti-Inflammatory, and Cytotoxic Activities. Plants 2022, 11, 1699. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- El-Garawani, I.; El-Seedi, H.; Khalifa, S.; El Azab, I.H.; Abouhendia, M.; Mahmoud, S. Enhanced Antioxidant and Cytotoxic Potentials of Lipopolysaccharides-Injected Musca Domestica Larvae. Pharmaceutics 2020, 12, 1111. [Google Scholar] [CrossRef]
- Marche, M.G.; Mura, M.E.; Ruiu, L. Rapid Polymerase Chain Reaction Assays for Brevibacillus Laterosporus Detection. J. Basic Microbiol. 2019, 59, 853–857. [Google Scholar] [CrossRef] [PubMed]
- De Mendiburu, F.; Simon, R. Agricolae-Ten Years of an Open Source Statistical Tool for Experiments in Breeding, Agriculture and Biology. PeerJ Prepr. 2015, 3, e1404v1. [Google Scholar] [CrossRef]
- R Development Core Team R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Deepak, P.; Kim, W.; Paley, M.A.; Yang, M.; Carvidi, A.B.; Demissie, E.G.; El-Qunni, A.A.; Haile, A.; Huang, K.; Kinnett, B. Effect of Immunosuppression on the Immunogenicity of MRNA Vaccines to SARS-CoV-2: A Prospective Cohort Study. Ann. Intern. Med. 2021, 174, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Stamatelopoulou, E.; Agriopoulou, S.; Dourtoglou, E.; Chatzilazarou, A.; Drosou, F.; Marinea, M.; Dourtoglou, V. Diketopiperazines in Wines. J. Wine Res. 2018, 29, 37–48. [Google Scholar] [CrossRef]
- Xing, J.; Yang, Z.; Lv, B.; Xiang, L. Rapid Screening for Cyclo-dopa and Diketopiperazine Alkaloids in Crude Extracts of Portulaca Oleracea L. Using Liquid Chromatography/Tandem Mass Spectrometry. Rapid Commun. Mass Spectrom. Int. J. Devoted Rapid Dissem. Up Minute Res. Mass Spectrom. 2008, 22, 1415–1422. [Google Scholar] [CrossRef]
- Pellegrino, R.M.; Blasi, F.; Angelini, P.; Ianni, F.; Alabed, H.B.R.; Emiliani, C.; Venanzoni, R.; Cossignani, L. LC/MS Q-TOF Metabolomic Investigation of Amino Acids and Dipeptides in Pleurotus Ostreatus Grown on Different Substrates. J. Agric. Food Chem. 2022, 70, 10371–10382. [Google Scholar] [CrossRef] [PubMed]
- Manganyi, M.C.; Ateba, C.N. Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms 2020, 8, 1934. [Google Scholar] [CrossRef] [PubMed]
- AK, A.U.; Demirkan, E.; Cengiz, M.; Sevgi, T.; Zeren, B.; Abdou, M. Optimization of Culture Medium for the Production and Partial Purification and Characterization of an Antibacterial Activity from Brevibacillus Laterosporus Strain EA62. Rom. Biotechnol. Lett. 2019, 24, 705–713. [Google Scholar] [CrossRef]
- Mohapatra, R.K.; Parhi, P.K.; Pandey, S.; Bindhani, B.K.; Thatoi, H.; Panda, C.R. Active and Passive Biosorption of Pb (II) Using Live and Dead Biomass of Marine Bacterium Bacillus Xiamenensis PbRPSD202: Kinetics and Isotherm Studies. J. Environ. Manag. 2019, 247, 121–134. [Google Scholar] [CrossRef]
- Oyedele, A.O.; Ogunbanwo, T.S. Antifungal Activities of Bacillus Subtilis Isolated from Some Condiments and Soil. Afr. J. Microbiol. Res. 2014, 8, 1841–1849. [Google Scholar] [CrossRef]
- Al-Ghazali, L.H.; Omran, R. Optimization of Medium Composition for Antibacterial Metabolite Production from Streptomyces sp. Asian J. Pharm. Clin. Res. 2017, 10, 381–385. [Google Scholar] [CrossRef]
- P de Carvalho, M.; Abraham, W.-R. Antimicrobial and Biofilm Inhibiting Diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Zhang, X.; Lai, D.; Zhou, L. Structural Diversity and Biological Activities of the Cyclodipeptides from Fungi. Molecules 2017, 22, 2026. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Salgado, R.; Tamayo-Galván, V.; Perraud-Gaime, I.; Rodríguez-Serrano, G.M.; González-Robles, R.O.; Durand, N.; Champion-Martínez, E.I.; Saucedo-Castañeda, G. Polyols Induce the Production of Antifungal Compounds by Lactobacillus Plantarum. Curr. Microbiol. 2022, 79, 99. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.-H.; Xu, Y.; Gao, J.; Qian, P.-Y.; Zhang, S. Antibacterial and Antilarval Compounds from Marine BacteriumPseudomonas Rhizosphaerae. Ann. Microbiol. 2009, 59, 229–233. [Google Scholar] [CrossRef]
- Brauns, S.C.; Milne, P.; Naudé, R.; Van de Venter, M. Selected Cyclic Dipeptides Inhibit Cancer Cell Growth and Induce Apoptosis in HT-29 Colon Cancer Cells. Anticancer Res. 2004, 24, 1713–1719. [Google Scholar] [PubMed]
- Brauns, S.C.; Dealtry, G.; Milne, P.; Naudé, R.; Van de Venter, M. Caspase-3 Activation and Induction of PARP Cleavage by Cyclic Dipeptide Cyclo(Phe-Pro) in HT-29 Cells. Anticancer Res. 2005, 25, 4197–4202. [Google Scholar]
- Graz, M.; Hunt, A.; Jamie, H.; Grant, G.; Milne, P. Antimicrobial Activity of Selected Cyclic Dipeptides. Pharmazie 1999, 54, 772–775. [Google Scholar]
- Hentzer, M.; Givskov, M. Pharmacological Inhibition of Quorum Sensing for the Treatment of Chronic Bacterial Infections. J. Clin. Investig. 2003, 112, 1300–1307. [Google Scholar] [CrossRef]
- Ström, K.; Sjögren, J.; Broberg, A.; Schnürer, J. Lactobacillus Plantarum MiLAB 393 Produces the Antifungal Cyclic Dipeptides Cyclo (l-Phe-l-Pro) and Cyclo (l-Phe-Trans-4-OH-l-Pro) and 3-Phenyllactic Acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef]
- Balaure, P.C.; Grumezescu, A.M. Recent Advances in Surface Nanoengineering for Biofilm Prevention and Control. Part II: Active, Combined Active and Passive, and Smart Bacteria-Responsive Antibiofilm Nanocoatings. Nanomaterials 2020, 10, 1527. [Google Scholar] [CrossRef]
- Desjardine, K.; Pereira, A.; Wright, H.; Matainaho, T.; Kelly, M.; Andersen, R.J. Tauramamide, a Lipopeptide Antibiotic Produced in Culture by Brevibacillus Laterosporus Isolated from a Marine Habitat: Structure Elucidation and Synthesis. J. Nat. Prod. 2007, 70, 1850–1853. [Google Scholar] [CrossRef]
- Liu, Z.; Jiao, D. Necroptosis, Tumor Necrosis and Tumorigenesis. Cell Stress 2019, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.N.; Mohandas, C.; Nambisan, B.; Sreerag, R.S.; Jayaprakas, C.A. Cyclo(l-Pro-d-Arg): A New Antibacterial and Antitumour Diketopiperazine from Bacillus Cereus Associated with a Rhabditid Entomopathogenic. Folia Microbiol. 2014, 59, 197–202. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Isolates | Inhibition Zone Diameters | |||
|---|---|---|---|---|
| E. coli | S. typhi | S. aureus | K. pneumoniae | |
| 1 | 9 ± 2.64 c | 11 ± 4.58 bc | 7 ± 1.73 d | 10 ± 3.46 bc |
| 2 | - | 16 ± 2.64 ab | 12 ± 3.00 bcd | 17 ± 2.00 ab |
| 3 | - | 7 ± 1.00 cd | - | 9 ± 1.73 c |
| 4 | 11 ± 1.00 bc | 15 ± 2.64 abc | - | 12 ± 2.00 abc |
| 5 | 21 ± 2.64 a | 20 ± 7.00 a | 20 ± 4.35 a | 19 ± 2.64 a |
| 6 | 15 ± 2.64 abc | 11 ± 2.64 bc | - | - |
| 7 | - | - | - | 12 ± 2.64 abc |
| 8 | 9 ± 1.00 c | 12 ± 1.73 abc | - | - |
| 9 | - | - | 11 ± 1.30 cd | 14 ± 3.30 abc |
| 10 | - | - | - | - |
| 11 | 11 ± 1.73 bc | 13 ± 3.00 abc | 10 ± 1.00 cd | 9 ± 1.73 c |
| 12 | - | - | 14 ± 4.35 abc | 12 ± 1.00 abc |
| 13 | 10 ± 4.35 bc | - | - | 12 ± 3.46 abc |
| 14 | - | 14 ± 2.00 abc | 10 ± 2.64 cd | 11 ± 2.64 bc |
| 15 | 15 ± 4.35 abc | 13 ± 4.35 abc | - | - |
| 16 | 16 ± 4.35 ab | 13 ± 3.60 abc | 15 ± 2.64 abc | 16 ± 4.35 abc |
| 17 | - | - | 15 ± 2.64 abc | 10 ± 1.00 bc |
| 18 | 11 ± 1.73 bc | 15 ± 2.64 abc | - | - |
| 19 | - | - | 18 ± 4.35 ab | 12 ± 3.46 abc |
| 20 | 19 ± 1.00 a | 17 ± 2.64 ab | - | 14 ± 2.64 abc |
| Pathogens | Inhibition Zone Diameters (mm) | |
|---|---|---|
| Bacterial Filtrate | Levofloxacin | |
| E. coli | 23 ± 1.00 b | 33 ± 2.64 a |
| S. typhi | 22 ± 2.64 b | 34 ± 3.60 a |
| S. aureus | 20 ± 2.64 b | 31 ± 2.64 a |
| K. pneumoniae | 19 ± 2.64 b | 32 ± 1.00 a |
| Compound Name | Parent Mass m/z (g/mol) | Adduct | Raw Material Fragments | Literature Fragment | Structure | Source |
|---|---|---|---|---|---|---|
| Cyclo (Leu-Pro) 1 | 211.257 | [M + H]+ | 183.37, 155.17, 138.19, 86.14 and 70.11 | 154.1, 138.1, 125.1, 110.1, 98.1, 84.1, and 70.1 [53] |  | Brevibacillus laterosporus- Bacillus sp. |
| Cyclo-(Val-Phe) 2 | 245.24 | [M-H]− | 219.16, 202.13 and 120.08 | 219, 202, 174, 157, and 120 [54] |  | Endophytic Streptomyces |
| Cyclo (Tyr-Pro) 3 | 261.188 | [M + H] + | 233.13, 155.08, 147.04 and 136.08 | NA |  | Bacillus sp. |
| Cyclo (Ile-Tyr) 4 | 277.29 | [M + H] + | 249.16, 232.13, 171.11 and 136.08 | 249, 232, 204,171, and 136 [54] |  | Marine-Derived Bacteria Harizani |
| Cyclo (Phe-Pro) 5 | 245.131 | [M + H] + | 120.09 and 70.07 | 217, 200, 172, 154, 120, and 70 [54] |  | Endophytic Streptomyces |
| Tyr-Pro 6 | 279.255 | [M + H] + | 136.08 and 116.07 | 147.04, 142.95, 137.07, 136.07, 121.05, 119.04, 118.06, 116.06, 109.06, 92.05, 71.06, and 70.06 [55] |  | Marine-Derived Bacteria |
| Cholic acid 7 | 374.454 | [M + H-2H2O] + | 355.21 and 337.19 | N/A |  | Fecal Bacteria from Cholesterol Gallstone Patients Marine Ascidian-Associated Bacterium Hasllibacter halocynthiae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zayed, M.; El-Garawani, I.M.; El-Sabbagh, S.M.; Amr, B.; Alsharif, S.M.; Tayel, A.A.; AlAjmi, M.F.; Ibrahim, H.M.S.; Shou, Q.; Khalifa, S.A.M.; et al. Structural Diversity, LC-MS-MS Analysis and Potential Biological Activities of Brevibacillus laterosporus Extract. Metabolites 2022, 12, 1102. https://doi.org/10.3390/metabo12111102
Zayed M, El-Garawani IM, El-Sabbagh SM, Amr B, Alsharif SM, Tayel AA, AlAjmi MF, Ibrahim HMS, Shou Q, Khalifa SAM, et al. Structural Diversity, LC-MS-MS Analysis and Potential Biological Activities of Brevibacillus laterosporus Extract. Metabolites. 2022; 12(11):1102. https://doi.org/10.3390/metabo12111102
Chicago/Turabian StyleZayed, Muhammad, Islam M. El-Garawani, Sabha M. El-Sabbagh, Bassem Amr, Sultan M. Alsharif, Ahmed A. Tayel, Mohamed F. AlAjmi, Hasnaa M. S. Ibrahim, Qiyang Shou, Shaden A. M. Khalifa, and et al. 2022. "Structural Diversity, LC-MS-MS Analysis and Potential Biological Activities of Brevibacillus laterosporus Extract" Metabolites 12, no. 11: 1102. https://doi.org/10.3390/metabo12111102
APA StyleZayed, M., El-Garawani, I. M., El-Sabbagh, S. M., Amr, B., Alsharif, S. M., Tayel, A. A., AlAjmi, M. F., Ibrahim, H. M. S., Shou, Q., Khalifa, S. A. M., El-Seedi, H. R., & Elfeky, N. (2022). Structural Diversity, LC-MS-MS Analysis and Potential Biological Activities of Brevibacillus laterosporus Extract. Metabolites, 12(11), 1102. https://doi.org/10.3390/metabo12111102