
Metabolism of Imidazole Dipeptides, Taurine, Branched-Chain Amino Acids, and Polyamines of the Breast Muscle Are Affected by Post-Hatch Development in Chickens

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Body and Tissue Weights
2.2. Targeted and Non-Targeted Metabolomic Analysis
2.2.1. Quantitative Analyses of Imidazole Dipeptides and Taurine in Breast Muscle and Plasma
2.2.2. Non-Targeted and Semi-Quantified Analyses of Metabolites in the Breast Muscle and Plasma
2.2.3. Pathway Analyses in the Breast Muscle and Plasma
2.2.4. Focusing on the Metabolites Based on the Pathway Analyses
2.2.5. mRNA Levels of Metabolic Enzymes and Insulin/IGF-1 Signaling-Related Factors
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Sample Preparation for Gas Chromatography/Mass Spectrometry Analysis
4.3. GC/MS Analysis
4.4. Data Processing for GC/MS Data
4.5. UPLC-MS/MS Analysis
4.6. Quantification of mRNA Levels by Real-Time PCR
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Cong, J.; Zhang, L.; Li, J.; Wang, S.; Gao, F.; Zhou, G. Effects of dietary supplementation with carnosine on growth performance, meat quality, antioxidant capacity and muscle fiber characteristics in broiler chickens. J. Sci. Food Agric. 2017, 97, 3733–3741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Jin, C.; Wang, X.; Yan, H.; Tan, H.; Gao, C. Dietary supplementation with pioglitazone hydrochloride and L-carnosine improves the growth performance, muscle fatty acid profiles and shelf life of yellow-feathered broiler chickens. Anim. Nutr. 2021, 7, 168–175. [Google Scholar] [CrossRef]
- Qi, B.; Wang, J.; Ma, Y.B.; Wu, S.G.; Qi, G.H.; Zhang, H.J. Effect of dietary β-alanine supplementation on growth performance, meat quality, carnosine content, and gene expression of carnosine-related enzymes in broilers. Poult. Sci. 2018, 97, 1220–1228. [Google Scholar] [CrossRef]
- Tomonaga, S.; Kaneko, K.; Yuji, K.; Kido, Y.; Denbow, D.M.; Furuse, M. Dietary β-alanine enhances brain, but not muscle, carnosine and anserine concentrations in broilers. Anim. Sci. J. 2006, 77, 79–86. [Google Scholar] [CrossRef]
- Han, H.L.; Zhang, J.F.; Yan, E.F.; Shen, M.M.; Wu, J.M.; Gan, Z.D.; Wei, C.H.; Zhang, L.L.; Wang, T. Effects of taurine on growth performance, antioxidant capacity, and lipid metabolism in broiler chickens. Poult. Sci. 2020, 99, 5707–5717. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, L.; Li, J.; Xing, T.; Jiang, Y.; Gao, F. Dietary taurine supplementation ameliorates muscle loss in chronic heat stressed broilers via suppressing the perk signaling and reversing endoplasmic reticulum-stress-induced apoptosis. J. Sci. Food Agric. 2021, 101, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Zhang, H.J.; Wu, S.G.; Yue, H.Y.; Zuo, J.J.; Feng, D.Y.; Qi, G.H. Effect of β-hydroxy-β-methylbutyrate calcium on growth, blood parameters, and carcass qualities of broiler chickens. Poult. Sci. 2013, 92, 753–759. [Google Scholar] [CrossRef]
- Smith, T.K. Effect of dietary putrescine on whole body growth and polyamine metabolism. Proc. Soc. Exp. Biol. Med. 1990, 194, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Sousadias, M.G.; Smith, T.K. Toxicity and growth-promoting potential of spermine when fed to chicks. J. Anim. Sci. 1995, 73, 2375–2381. [Google Scholar] [CrossRef]
- Jacob, J.P.; Blair, R.; Hart, L.E.; Gardiner, E.E. The effect of taurine transport antagonists on cardiac taurine concentration and the incidence of sudden death syndrome in male broiler chickens. Poult. Sci. 1991, 70, 561–567. [Google Scholar] [CrossRef]
- Scanes, C.G. Perspectives on the endocrinology of poultry growth and metabolism. Gen. Comp. Endocrinol. 2009, 163, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Tamemoto, H.; Kadowaki, T.; Tobe, K.; Yagi, T.; Sakura, H.; Hayakawa, T.; Terauchi, Y.; Ueki, K.; Kaburagi, Y.; Satoh, S.; et al. Insulin resistance and growth retardation in mice lacking insulin receptor substrate-1. Nature 1994, 372, 182–186. [Google Scholar] [CrossRef]
- Toyoshima, Y.; Nakamura, K.; Tokita, R.; Teramoto, N.; Sugihara, H.; Kato, H.; Yamanouchi, K.; Minami, S. Disruption of insulin receptor substrate-2 impairs growth but not insulin function in rats. J. Biol. Chem. 2020, 295, 11914–11927. [Google Scholar] [CrossRef]
- Nakashima, K.; Ishida, A.; Shimamoto, S.; Ijiri, D.; Ohtsuka, A. Insulin Stimulation of Protein Synthesis and mTOR Signaling in Chick Myotube Cultures. J. Poult. Sci. 2020, 57, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Ishida, A.; Shimamoto, S.; Ijiri, D.; Ohtsuka, A. Effects of Insulin-Like Growth Factor-I on the Expression of Atrogin-1/MAFbx in Chick Myotube Cultures. J. Poult. Sci. 2017, 54, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kobayashi, H.; Mawatari, K.; Sato, J.; Bajotto, G.; Kitaura, Y.; Shimomura, Y. Effects of branched-chain amino acid supplementation on plasma concentrations of free amino acids, insulin, and energy substrates in young men. J. Nutr. Sci. Vitaminol. 2011, 57, 114–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.M.; Rho, J.Y.; Suzuki, M.; Nishihara, M.; Takahashi, M. Effect of taurine in rat milk on the growth of offspring. J. Vet. Med. Sci. 2000, 62, 693–698. [Google Scholar] [CrossRef] [Green Version]
- Tomonaga, S.; Okuyama, H.; Tachibana, T.; Makino, R. Effects of high ambient temperature on plasma metabolomic profiles in chicks. Anim. Sci. J. 2018, 89, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying metabolites by integrating metabolome databases with mass spectrometry cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Salazar, C.; Armenta, J.M.; Shulaev, V. An UPLC-ESI-MS/MS Assay Using 6-Aminoquinolyl-N-Hydroxysuccinimidyl Carbamate Derivatization for Targeted Amino Acid Analysis: Application to Screening of Arabidopsis thaliana Mutants. Metabolites 2012, 2, 398–428. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. Metabo Analyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| P7 | P28 | P42 | KW p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | |||||
| Initial body weight (g) | 63.4 | 67.8 | 83.9 | 66.2 | 67.2 | 70.0 | 65.1 | 69.6 | 73.7 | 0.756 | |||
| Final body weight (g) | C | 63.4 | 67.8 | 83.9 | B | 335.3 | 337.0 | 341.8 | A | 605.0 | 640.0 | 660.0 | <0.005 |
| Breast muscle weight (g) | C | 0.98 | 1.15 | 1.35 | B | 2.02 | 2.08 | 2.13 | A | 2.46 | 2.50 | 2.59 | <0.005 |
| Liver weight (g) | C | 2.62 | 2.91 | 3.03 | B | 10.38 | 11.20 | 12.03 | A | 19.39 | 20.67 | 21.95 | <0.005 |
| Brain weight (g) | C | 1.14 | 1.15 | 1.18 | B | 8.65 | 9.25 | 10.30 | A | 14.66 | 15.46 | 16.76 | <0.005 |
| Metab. No. | Pathway Name | Breast Muscle | Plasma |
|---|---|---|---|
| 1 | Valine, leucine, and isoleucine biosynthesis | * | ** |
| 2 | Alanine, aspartate, and glutamate metabolism | * | * |
| 3 | Glyoxylate and dicarboxylate metabolism | * | * |
| 4 | Aminoacyl-tRNA biosynthesis | ** | |
| 5 | Arginine and proline metabolism | * | |
| 6 | Arginine biosynthesis | * | |
| 7 | β-Alanine metabolism | * | |
| 8 | Cysteine and methionine metabolism | * | |
| 9 | D-Glutamine and D-glutamate metabolism | * | |
| 10 | Glutathione metabolism | * | |
| 11 | Glycine, serine, and threonine metabolism | * | |
| 12 | Histidine metabolism | * | |
| 13 | Nitrogen metabolism | * | |
| 14 | Phenylalanine metabolism | * | |
| 15 | Phenylalanine, tyrosine, and tryptophan biosynthesis | * | |
| 16 | Taurine and hypotaurine metabolism | * | |
| 17 | Ascorbate and aldarate metabolism | * | |
| 18 | Citrate cycle | * | |
| 19 | Glycerolipid metabolism | * | |
| 20 | Pantothenate and CoA biosynthesis | * | |
| 21 | Galactose metabolism | * | |
| 22 | Valine, leucine, and isoleucine degradation | * |
| P7 | P28 | P42 | KW p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | |||||
| Breast muscle | |||||||||||||
| Carnosine | B | 76.6 | 104.8 | 121.0 | A | 188.9 | 222.7 | 308.1 | A | 205.9 | 280.2 | 354.4 | <0.05 |
| Anserine | B | 71.9 | 95.0 | 130.6 | A | 212 | 311.7 | 376.4 | A | 232.2 | 273.7 | 338.5 | <0.05 |
| Balenine | 84.9 | 100.6 | 114.8 | 140.8 | 154.1 | 175.5 | 57.5 | 158.2 | 180.2 | 0.055 | |||
| Total imidazole dipeptides | B | 76.8 | 91.2 | 127.6 | A | 204.7 | 318.7 | 337.4 | A | 242.1 | 291.4 | 315.5 | <0.05 |
| β-Alanine | 58.3 | 104.7 | 139.3 | 98.7 | 121.8 | 169.7 | 63.8 | 98.8 | 148.7 | 0.472 | |||
| 3-Hydroxypropionic acid | A | 78.3 | 107.8 | 117.8 | A | 109.3 | 134.3 | 160.0 | A | 127.5 | 173.3 | 187.9 | <0.05 |
| Aspartic acid | 89.5 | 99.4 | 110.8 | 81.8 | 93.5 | 100.7 | 89.2 | 90.4 | 92.4 | 0.310 | |||
| Uracil | 84.9 | 102.6 | 113.8 | 92.6 | 113.9 | 126.3 | 93.1 | 100.9 | 103.3 | 0.527 | |||
| Uridine | 92.0 | 101.2 | 107.4 | 94.7 | 111.3 | 119.3 | 99.5 | 103.7 | 113.5 | 0.472 | |||
| Spermidine | A | 70.7 | 110.8 | 123.9 | B | 30.8 | 37.3 | 38.8 | B | 14.7 | 24.6 | 27.3 | <0.05 |
| Plasma | |||||||||||||
| Carnosine | 45.5 | 85.5 | 161.7 | 41.3 | 61.9 | 91.6 | 83.1 | 118.8 | 259.3 | 0.168 | |||
| Anserine | 38.7 | 111.8 | 155.4 | 77.4 | 88.5 | 144.3 | 112.5 | 141.6 | 321 | 0.298 | |||
| Total imidazole dipeptides | 41.2 | 109.4 | 154.1 | 63.9 | 78.6 | 124.7 | 116.1 | 124.4 | 292.1 | 0.176 | |||
| β-Alanine | 85.7 | 99.8 | 114.4 | 95.6 | 102.2 | 131.1 | 94.7 | 117.4 | 151.5 | 0.432 | |||
| 3-Hydroxypropionic acid | 91.9 | 100.1 | 108.0 | 97.0 | 101.5 | 103.6 | 89.9 | 93.5 | 106.2 | 0.403 | |||
| Uracil | 85.9 | 96.4 | 115.9 | 113.8 | 120.1 | 140.8 | 102.2 | 116.0 | 148.0 | 0.087 | |||
| Uridine | 83.9 | 105.2 | 113.5 | 108.5 | 112.8 | 158.9 | 98.1 | 109.2 | 200.3 | 0.310 | |||
| Histidine | 86.5 | 96.1 | 115.5 | 95.7 | 128 | 144.8 | 81.5 | 135.5 | 177.1 | 0.403 | |||
| P7 | P28 | P42 | KW p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | |||||
| Breast muscle | |||||||||||||
| Taurine | A | 80.0 | 91.1 | 124.4 | B | 18.5 | 22.1 | 26.2 | B | 18.4 | 25.6 | 31.7 | <0.05 |
| Cystathionine | A | 53.9 | 57.8 | 167.2 | B | 0.2 | 0.4 | 0.6 | B | 0.1 | 0.2 | 0.5 | <0.05 |
| Homocysteine | A | 82.6 | 91.0 | 121.9 | B | 3.3 | 4.2 | 6.7 | B | 1.8 | 2.6 | 7.3 | <0.05 |
| Homocystine | A | 53.9 | 69.7 | 161.3 | B | 1.2 | 3.6 | 9.7 | B | 4.7 | 8.1 | 11.5 | <0.05 |
| Hypotaurine | A | 73.5 | 85.6 | 133.7 | B | 20.4 | 25.9 | 34.9 | B | 20.0 | 24.6 | 28.9 | <0.05 |
| Methionine | A | 66.8 | 100.0 | 133.2 | B | 36.9 | 46.2 | 59.0 | AB | 39.1 | 49.6 | 64.1 | <0.05 |
| Plasma | |||||||||||||
| Taurine | 22.0 | 76.5 | 189.8 | 25.6 | 43.0 | 131.3 | 123.5 | 181.7 | 198.2 | 0.131 | |||
| Cystathionine | 78.5 | 110.8 | 116.1 | 52.1 | 59.4 | 90.6 | 39.0 | 47.0 | 70.8 | 0.075 | |||
| Cysteine | 62.0 | 105.4 | 135.3 | 98.9 | 115.3 | 143.9 | 92.7 | 106.4 | 148.7 | 0.827 | |||
| Cystine | A | 86.5 | 100.8 | 113.1 | A | 38.2 | 55.1 | 75.9 | A | 60.2 | 94.6 | 103.3 | <0.05 |
| Homocysteine | 69.0 | 110.2 | 125.9 | 68.6 | 79.5 | 122.8 | 51.0 | 68.6 | 110.5 | 0.566 | |||
| Homocystine | A | 74.4 | 109.5 | 120.8 | B | 15.5 | 27.2 | 31.5 | C | 4.9 | 6.3 | 7.9 | <0.05 |
| Hypotaurine | 64.7 | 84.1 | 143.3 | 53.6 | 55.7 | 82.3 | 78.9 | 109.9 | 113.3 | 0.112 | |||
| Methionine | 77.7 | 97.7 | 123.4 | 77.8 | 86.7 | 99.8 | 95.8 | 117.8 | 138.6 | 0.141 | |||
| P7 | P28 | P42 | KW p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | |||||
| Breast muscle | |||||||||||||
| Valine | A | 73.8 | 101.6 | 125.4 | B | 31.2 | 38.3 | 41.3 | B | 35.7 | 41.1 | 49.0 | <0.05 |
| Leucine | A | 60.5 | 100.4 | 139.3 | B | 24.5 | 34.0 | 40.2 | B | 25.7 | 38.7 | 47.1 | <0.05 |
| Isoleucine | A | 85.0 | 98.1 | 115.9 | B | 44.4 | 49.7 | 51.4 | B | 41.0 | 44.5 | 54.8 | <0.05 |
| Glutamic acid | A | 83.2 | 103.6 | 115.0 | B | 29.1 | 32.3 | 40.7 | B | 29.8 | 34.4 | 39.0 | <0.05 |
| 2-Keto-isovaleric acid | 48.1 | 78.9 | 162.4 | 62.0 | 87.5 | 146.9 | 64.7 | 99.7 | 133.7 | 0.852 | |||
| 2-Methyl- 3-hydroxybutyric acid | 11.2 | 118.6 | 179.5 | 27.7 | 126.0 | 255.4 | 48.9 | 72.0 | 112.1 | 0.651 | |||
| Plasma | |||||||||||||
| Valine | A | 79.4 | 110.0 | 115.6 | A | 48.8 | 54.2 | 77.3 | A | 64.3 | 81.2 | 83.0 | <0.05 |
| Leucine | 83.3 | 103.5 | 114.9 | 56.9 | 62.1 | 85.5 | 75.3 | 88.3 | 94.4 | 0.061 | |||
| Isoleucine | 87.2 | 95.5 | 115.1 | 65.5 | 70.7 | 92.4 | 70.7 | 85.3 | 95.1 | 0.164 | |||
| 2-Hydroxyisovaleric acid | A | 74.2 | 78.2 | 136.6 | B | 28.3 | 37.4 | 54.0 | B | 19.2 | 21.6 | 27.0 | <0.05 |
| 2-Hydroxy- 3-methylvaleric acid | A | 51.6 | 88.6 | 154.1 | AB | 20.0 | 41.4 | 62.6 | B | 8.7 | 9.9 | 14.3 | <0.05 |
| 2-Keto-isovaleric acid | A | 71.9 | 97.0 | 129.6 | B | 21.9 | 30.2 | 50.0 | B | 16.0 | 27.1 | 48.9 | <0.05 |
| 2-Ketoisocaproic acid | A | 86.0 | 96.6 | 115.7 | AB | 58.3 | 70.6 | 75.0 | B | 48.8 | 58.9 | 78.4 | <0.05 |
| 2-Methyl- 3-hydroxybutyric acid | A | 80.5 | 98.4 | 120.3 | B | 49.7 | 56.2 | 78.7 | AB | 52.7 | 68.7 | 86.6 | <0.05 |
| 3-Methyl- 2-oxovaleric acid | A | 82.8 | 94.8 | 119.8 | B | 41.5 | 54.7 | 65.0 | B | 34.8 | 47.0 | 61.5 | <0.05 |
| HMB | A | 54.9 | 111.1 | 139.6 | B | 19.6 | 21.6 | 25.6 | C | 15.3 | 17.0 | 17.4 | <0.05 |
| P7 | P28 | P42 | KW p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | |||||
| Breast muscle | |||||||||||||
| Arginine | A | 70.2 | 99.0 | 130.3 | B | 16.3 | 18.4 | 25.0 | B | 13.3 | 17.6 | 23.7 | <0.05 |
| Ornithine | A | 75.7 | 94.0 | 127.3 | B | 17.8 | 21.2 | 27.0 | B | 15.2 | 20.2 | 24.2 | <0.05 |
| Proline | A | 86.5 | 95.4 | 115.8 | B | 8.4 | 14.8 | 19.8 | B | 8.6 | 9.9 | 12.3 | <0.05 |
| Putrescine | A | 66.7 | 94.3 | 136.1 | B | 18.9 | 22.6 | 28.0 | B | 11.3 | 15.5 | 17.8 | <0.05 |
| Spermidine | A | 70.7 | 110.8 | 123.9 | B | 30.8 | 37.3 | 38.8 | B | 14.7 | 24.6 | 27.3 | <0.05 |
| Glutamic acid | A | 83.2 | 103.6 | 115.0 | B | 29.1 | 32.3 | 40.7 | B | 29.8 | 34.4 | 39.0 | <0.05 |
| Plasma | |||||||||||||
| Ornithine | 88.7 | 100.4 | 111.1 | 78.4 | 101.3 | 124.8 | 93.8 | 109.5 | 133.9 | 0.619 | |||
| Proline | 90.5 | 99.8 | 109.6 | 50.1 | 76.0 | 88.0 | 64.7 | 77.2 | 101.8 | 0.061 | |||
| P7 | P28 | P42 | KW p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | Q1 | Q2 | Q3 | |||||
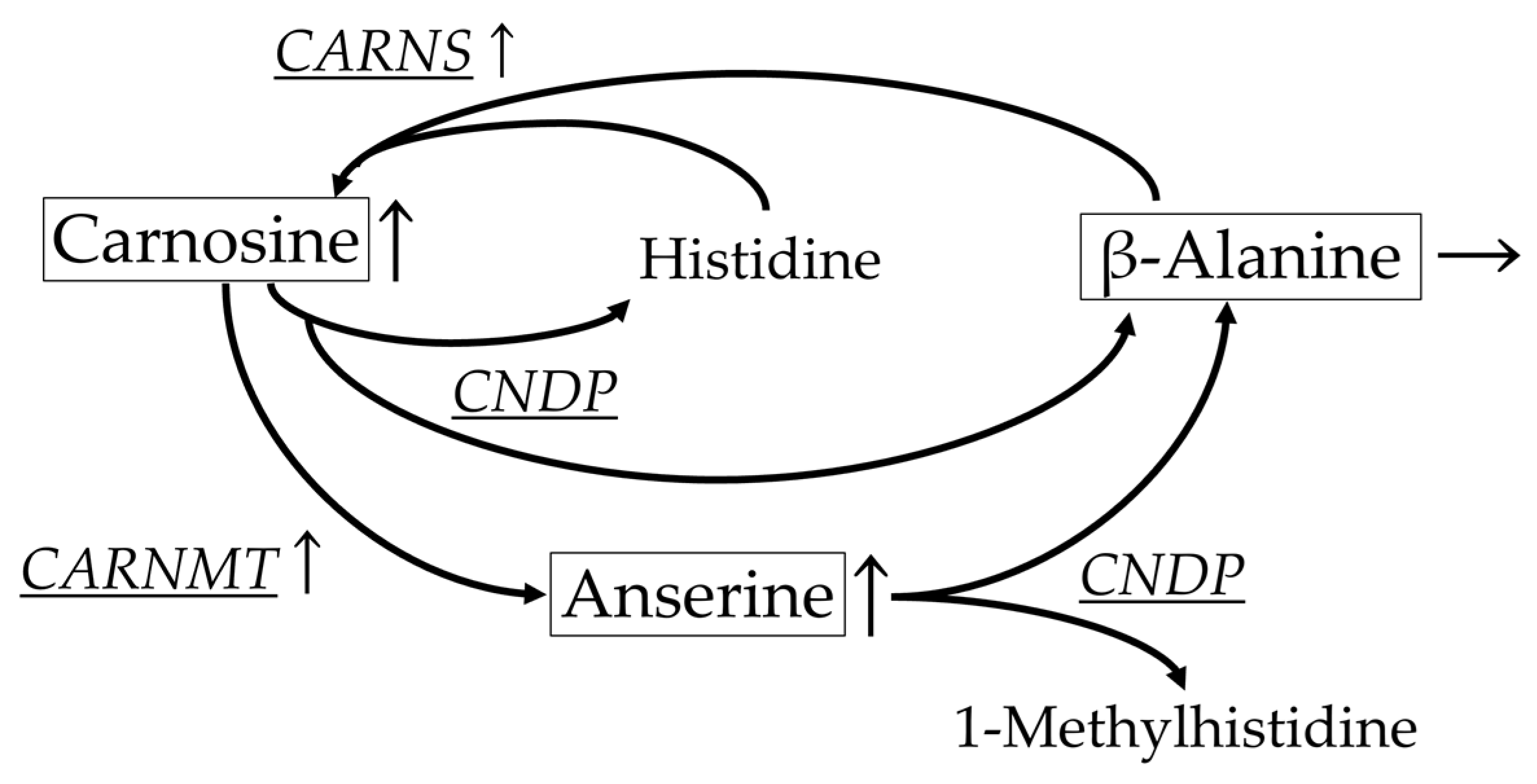
| Carnosine metabolism | |||||||||||||
| CARNS | B | 79.1 | 93.7 | 124.0 | AB | 111.8 | 122.0 | 135.3 | A | 130.8 | 135.4 | 143.1 | <0.05 |
| CNDP | A | 44.0 | 61.1 | 175.5 | A | 24.2 | 37.4 | 48.0 | A | 8.0 | 10.8 | 53.9 | <0.05 |
| CARNMT | B | 83.4 | 101.5 | 115.8 | A | 141.9 | 148.0 | 150.7 | A | 142.9 | 170.0 | 180.8 | <0.05 |
| Sulfur amino acid degradation | |||||||||||||
| CTH | B | 86.3 | 101.2 | 113.1 | A | 136.7 | 154.4 | 205.3 | A | 155.7 | 162.3 | 200.3 | <0.05 |
| Branched-chain amino acid degradation | |||||||||||||
| BCAT | A | 86.1 | 93.2 | 117.3 | A | 79.4 | 83.8 | 103.7 | B | 47.1 | 49.4 | 60.1 | <0.05 |
| BCKDH E1a | A | 87.7 | 94.4 | 115.1 | A | 120.8 | 141.0 | 156.0 | A | 109.1 | 116.4 | 139.7 | <0.05 |
| BCKDH E1b | 84.3 | 92.9 | 119.2 | 68.4 | 82.4 | 98.4 | 75.3 | 78.3 | 95.4 | 0.265 | |||
| Polyamine synthesis | |||||||||||||
| ODC | 78.1 | 101.1 | 121.3 | 75.8 | 83.2 | 94.5 | 82.3 | 90.4 | 102.9 | 0.432 | |||
| AMD | 83.2 | 105.6 | 114.1 | 93.4 | 102.8 | 113.9 | 97.7 | 111.3 | 143.2 | 0.472 | |||
| Insulin/IGF-1 signaling | |||||||||||||
| INSR | B | 87.7 | 98.5 | 113.1 | A | 143.3 | 153.7 | 166.4 | A | 175.5 | 201.7 | 224.8 | <0.05 |
| IGF1R | B | 83.9 | 96.7 | 117.7 | AB | 113.1 | 127.0 | 136.4 | A | 127.3 | 146.5 | 151.2 | <0.05 |
| IRS1 | B | 84.8 | 99.8 | 115.3 | A | 178.8 | 198.2 | 216.5 | A | 203.5 | 210.0 | 225.9 | <0.05 |
| IRS2 | B | 82.7 | 97.3 | 118.7 | AB | 116.0 | 145.5 | 182.7 | A | 157.2 | 184.4 | 256.7 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomonaga, S.; Kawase, T.; Tsukahara, T.; Ohta, Y.; Shiraishi, J.-i. Metabolism of Imidazole Dipeptides, Taurine, Branched-Chain Amino Acids, and Polyamines of the Breast Muscle Are Affected by Post-Hatch Development in Chickens. Metabolites 2022, 12, 86. https://doi.org/10.3390/metabo12010086
Tomonaga S, Kawase T, Tsukahara T, Ohta Y, Shiraishi J-i. Metabolism of Imidazole Dipeptides, Taurine, Branched-Chain Amino Acids, and Polyamines of the Breast Muscle Are Affected by Post-Hatch Development in Chickens. Metabolites. 2022; 12(1):86. https://doi.org/10.3390/metabo12010086
Chicago/Turabian StyleTomonaga, Shozo, Takahiro Kawase, Takamitsu Tsukahara, Yoshiyuki Ohta, and Jun-ichi Shiraishi. 2022. "Metabolism of Imidazole Dipeptides, Taurine, Branched-Chain Amino Acids, and Polyamines of the Breast Muscle Are Affected by Post-Hatch Development in Chickens" Metabolites 12, no. 1: 86. https://doi.org/10.3390/metabo12010086
APA StyleTomonaga, S., Kawase, T., Tsukahara, T., Ohta, Y., & Shiraishi, J.-i. (2022). Metabolism of Imidazole Dipeptides, Taurine, Branched-Chain Amino Acids, and Polyamines of the Breast Muscle Are Affected by Post-Hatch Development in Chickens. Metabolites, 12(1), 86. https://doi.org/10.3390/metabo12010086