
Anti-Obesity Effect of Hot Water Extract of Barley Sprout through the Inhibition of Adipocyte Differentiation and Growth

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
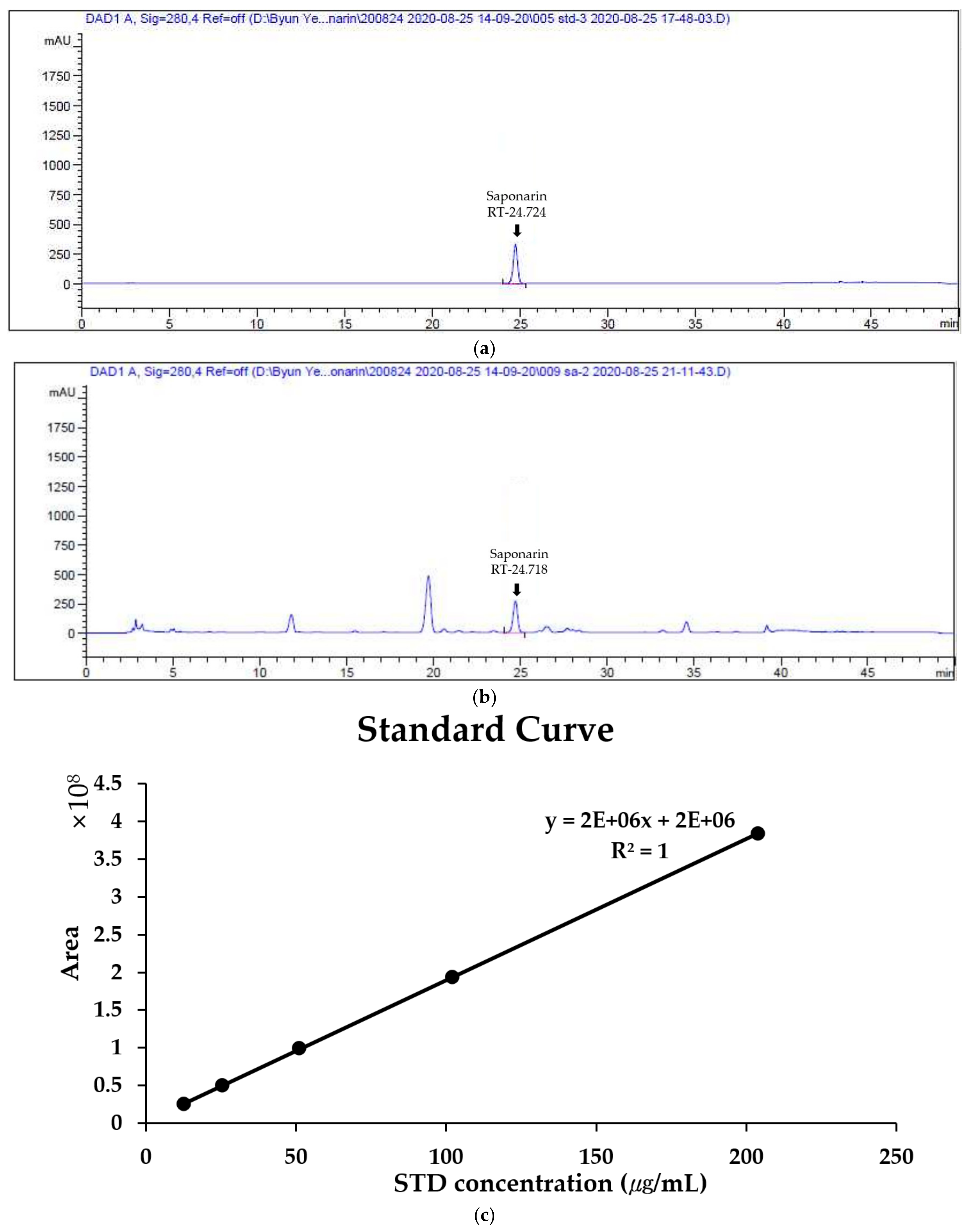
2.1. Quantitative Analysis of Saponarin in BSE
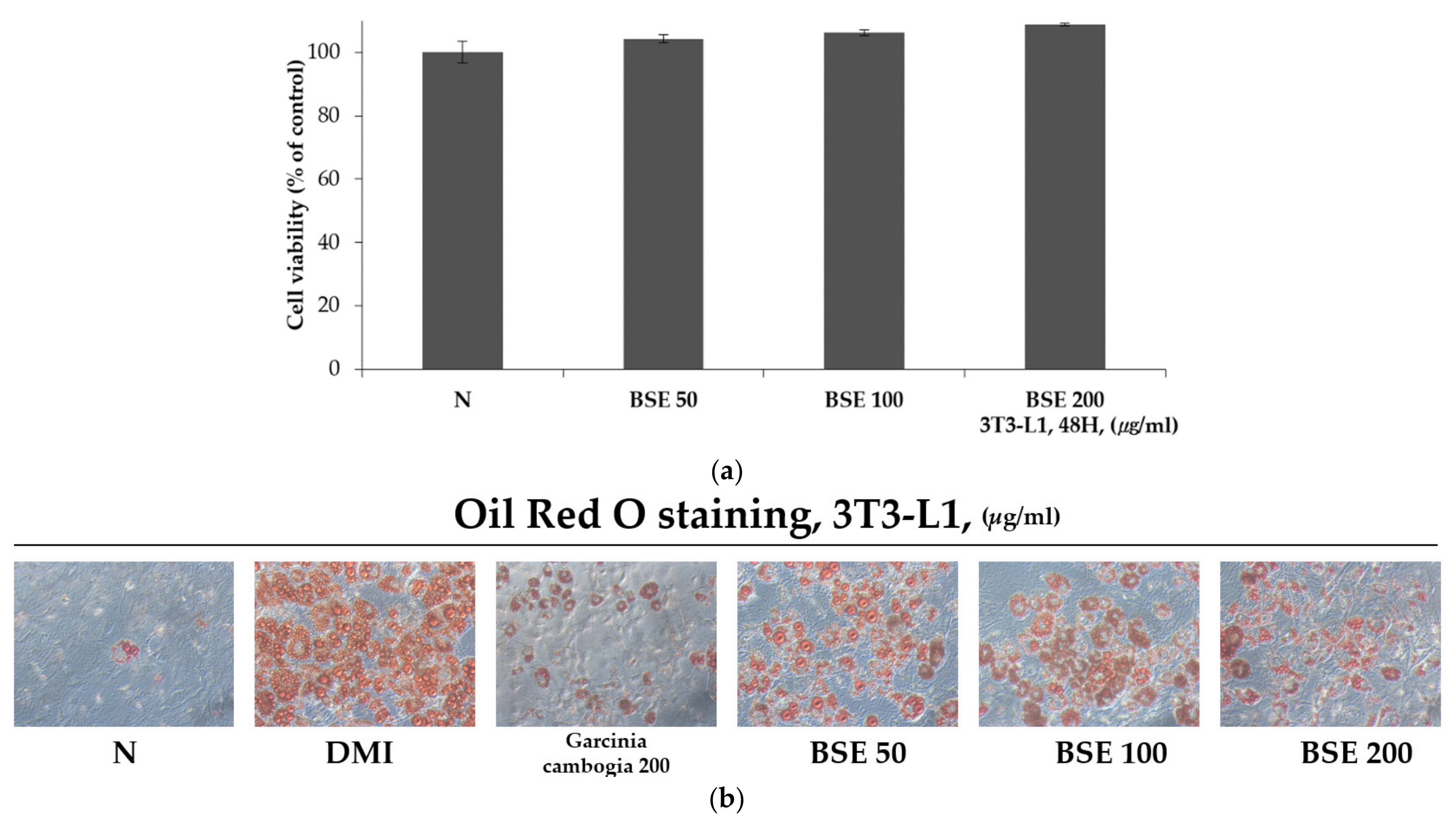
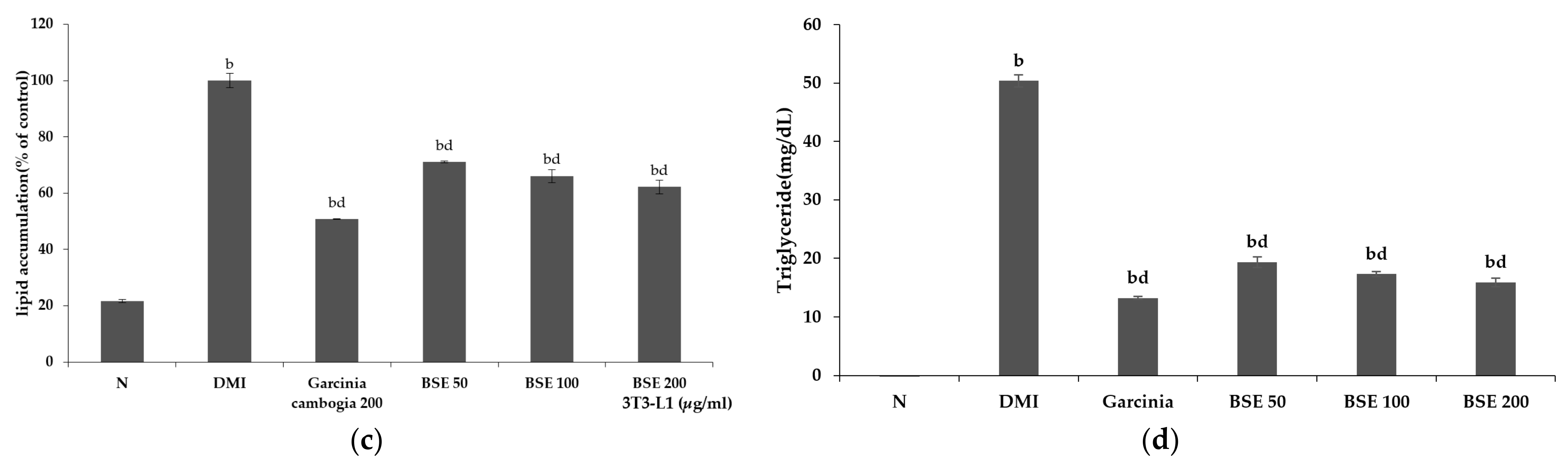
2.2. Inhibitory Effect of BSE Differentiation into Adipocytes without Cytotoxicity
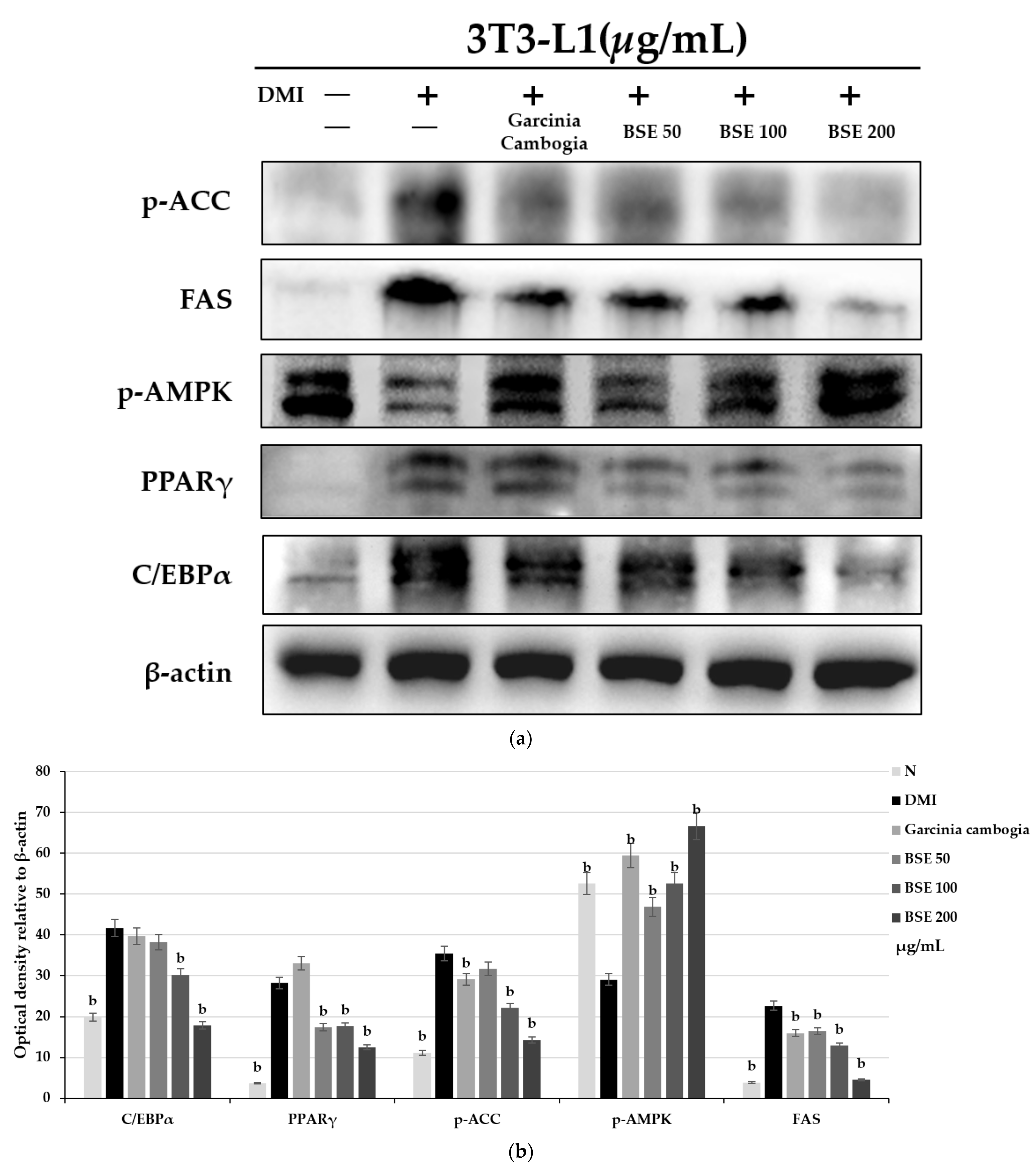
2.3. Effect of BSE on Adipocyte-Differentiation-Related Protein Expression Change
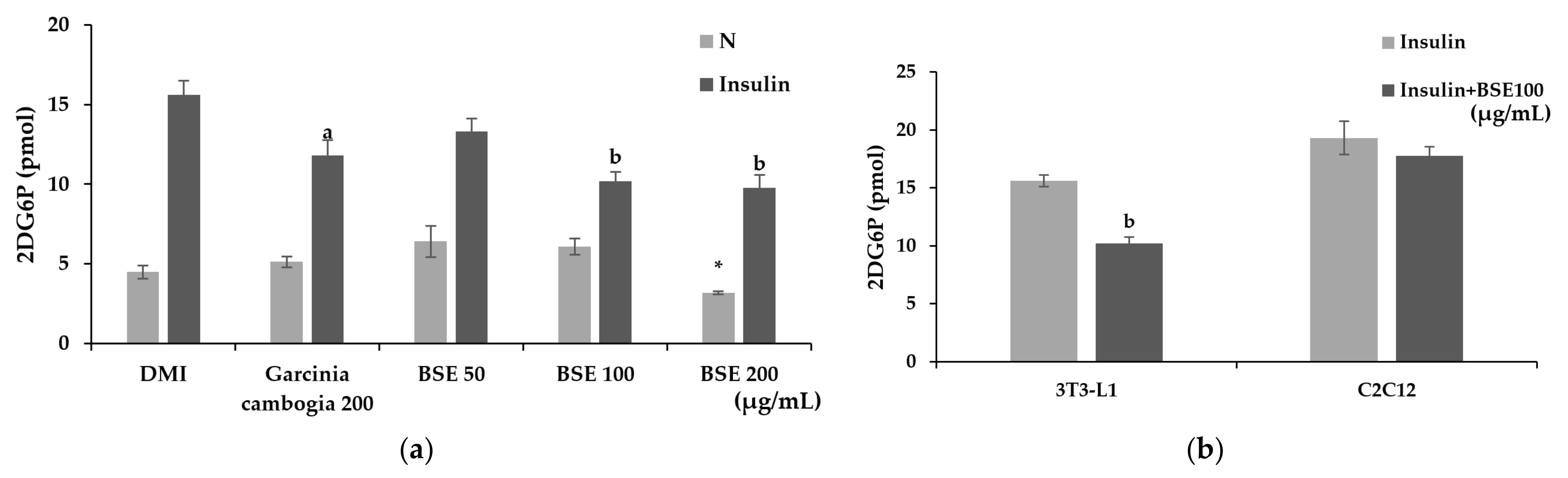
2.4. Inhibitory Effect of BSE on Glucose Uptake in Adipocytes
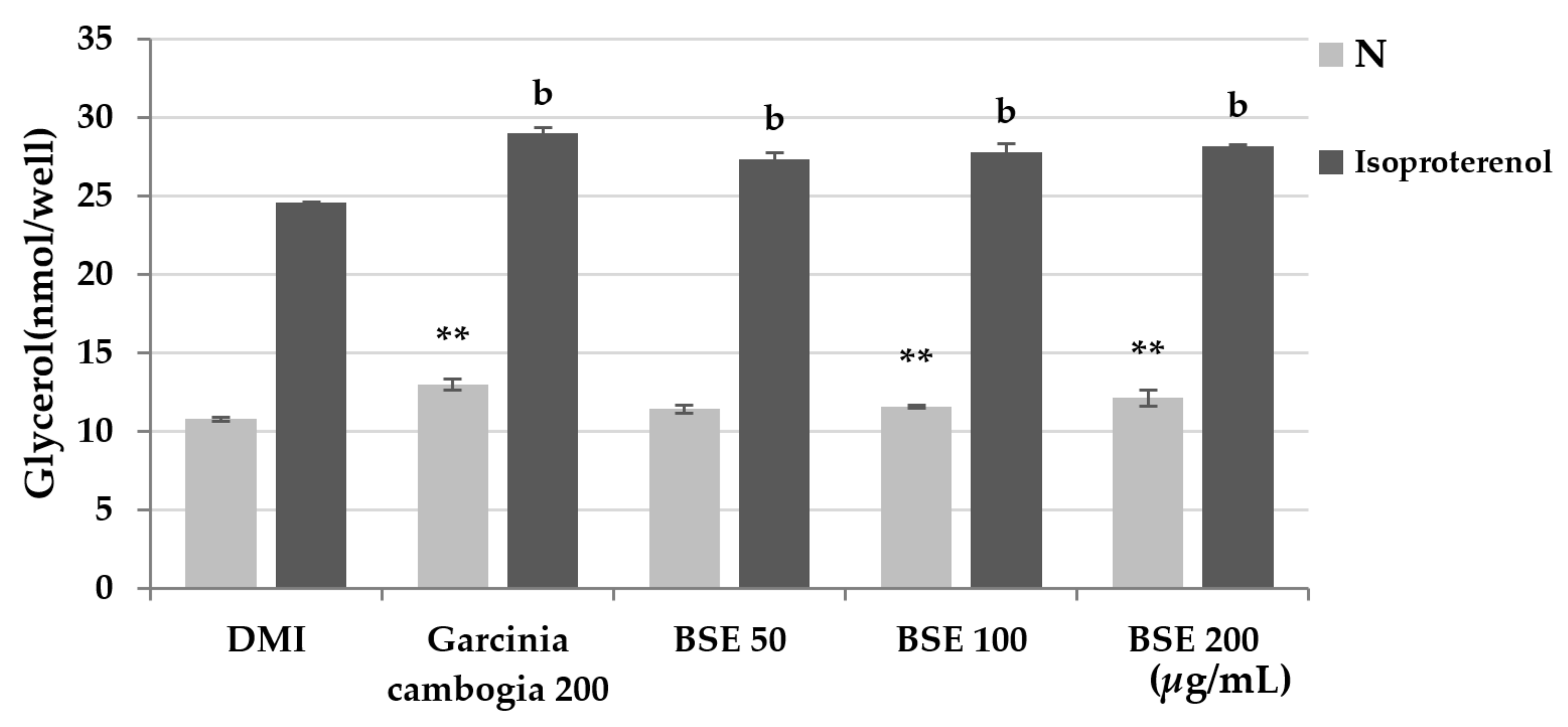
2.5. Confirmation of Lipolysis Effect by BSE
2.6. Confirmation of Changes in Body and Tissue Weight and Food Intake of Experimental Animals by BSE
2.7. Confirmation of Blood Biochemical Changes by BSE
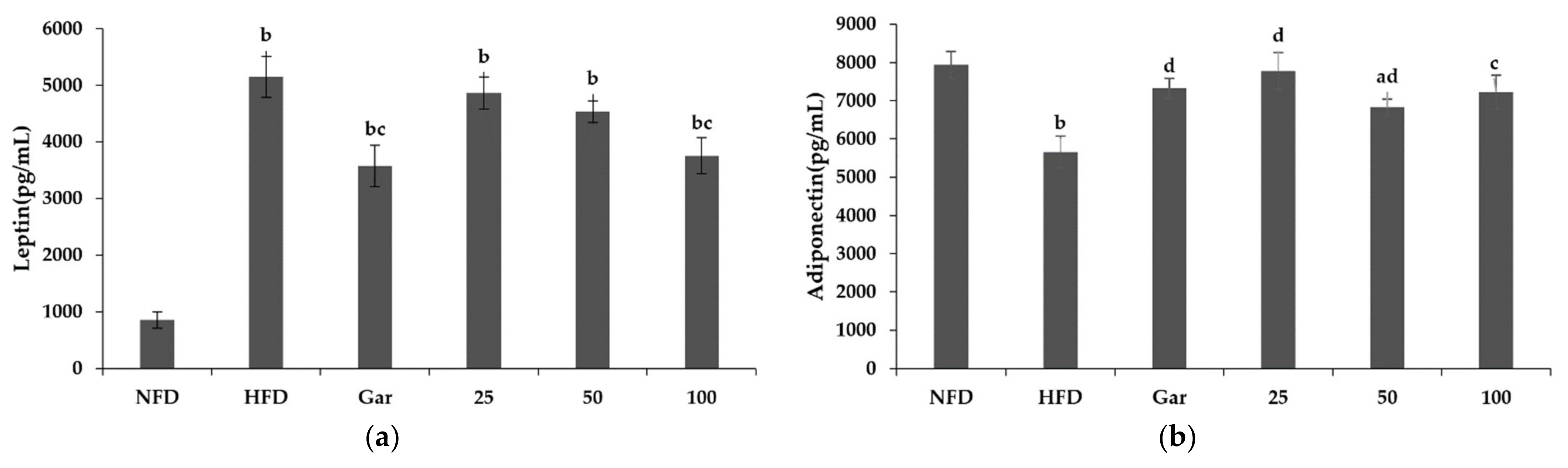
2.8. Confirmation of Changes in Leptin and Adiponectin Levels by BSE
2.9. Effect of BSE on Adipose Tissue Histology and Adipocyte-Differentiation-Related Factors
2.10. Effect of BSE on Liver Histology and Adipocyte-Differentiation-Related Factors
3. Discussion
4. Materials and Methods
4.1. Reagent
4.2. Quantitative Analysis of Saponarin in BSE Using HPLC
4.3. Cell Culture
4.4. WST-1 Assay
4.5. Differentiation Induction
4.6. Oil Red O Staining
4.7. TG Assay
4.8. Western Blotting
4.9. Glucose Uptake Assay
4.10. Lipolysis Assay
4.11. Breeding and Diet of Experimental Animals
4.12. Body Weight and Food Efficiency Measurements
4.13. Blood and Tissue Collection and Analysis
4.14. Significance Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, L.; Aronne, L.J.; Beilin, L.J.; Burke, V.; Igel, L.I.; Lloyd-Jones, D.; Sowers, J. Obesity-related hypertension: Pathogenesis, cardiovascular risk, and treatment: A position paper of The Obesity Society and the American Society of Hypertension. J. Clin. Hypertens. 2013, 15, 14–33. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.Y. Obesity-related digestive diseases and their pathophysiology. Gut Liver 2017, 11, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Boles, A.; Kandimalla, R.; Reddy, P.H. Dynamics of diabetes and obesity: Epidemiological perspective. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. Obesity and diabetes: The increased risk of cancer and cancer-related mortality. Physiol. Rev. 2015, 95, 727–748. [Google Scholar] [CrossRef] [Green Version]
- De Pergola, G.; Silvestris, F. Obesity as a major risk factor for cancer. J. Obes. 2013, 2013, 291546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallone, F.; Deudon, R.; Muller, C.; Vaysse, C. Breast cancer, obesity and adipose tissue: A high-risk combination. Med. Sci. 2018, 34, 1079–1086. [Google Scholar] [CrossRef]
- Tomiyama, A.J. Stress and obesity. Ann. Rev. Psychol. 2019, 70, 703–718. [Google Scholar] [CrossRef] [Green Version]
- Fock, K.M.; Khoo, J. Diet and exercise in management of obesity and overweight. J. Gastroenterol. Hepatol. 2013, 28, 59–63. [Google Scholar] [CrossRef]
- Saris, W.H.M.; Foster, G.D. Simple carbohydrates and obesity: Fact, fiction and future. Int. J. Obes. 2006, 30, S1–S3. [Google Scholar] [CrossRef]
- Tappy, L.; Le, K.A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [Green Version]
- Christou, G.A.; Katsiki, N.; Blundell, J.; Fruhbeck, G.; Kiortsis, D.N. Semaglutide as a promising antiobesity drug. Obes. Rev. 2019, 20, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Xu, L.; Liu, H.; Lv, Y.; Zheng, Q.; Li, L. Comparative efficacy of five long-term weight loss drugs: Quantitative information for medication guidelines. Obes. Rev. 2017, 18, 1377–1385. [Google Scholar] [CrossRef]
- Bessesen, D.H.; Van Gaal, L.F. Progress and challenges in anti-obesity pharmacotherapy. Lancet Diabetes Endocrinol. 2018, 6, 237–248. [Google Scholar] [CrossRef]
- Krentz, A.J.; Fujioka, K.; Hompesch, M. Evolution of pharmacological obesity treatments: Focus on adverse side-effect profiles. Diabetes Obes. Metab. 2016, 18, 558–570. [Google Scholar] [CrossRef]
- Bersoux, S.; Byun, T.H.; Chaliki, S.S.; Poole, K.G. Pharmacotherapy for obesity: What you need to know. Cleve. Clin. J. Med. 2017, 84, 951–958. [Google Scholar] [CrossRef]
- Costa, A.G.V.; Garcia-Diaz, D.F.; Jimenez, P.; Silva, P.I. Bioactive compounds and health benefits of exotic tropical red–black berries. J. Funct. Foods 2013, 5, 539–549. [Google Scholar] [CrossRef]
- Kang, J.G.; Park, C.Y. Anti-obesity drugs: A review about their effects and safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Mayer, M.A.; Hocht, C.; Puyó, A.; Taira, C.A. Recent advances in obesity pharmacotherapy. Curr. Clin. Pharmacol. 2009, 4, 53–61. [Google Scholar] [CrossRef]
- Kawk, H.W.; Nam, G.H.; Kim, M.J.; Kim, S.Y.; Kim, G.N.; Kim, Y.M. Anti-obesity effect of an ethanol extract of cheongchunchal in vitro and in vivo. Nutrients 2020, 12, 3453. [Google Scholar] [CrossRef] [PubMed]
- Im, J.Y.; Ki, H.H.; Xin, M.; Kwon, S.U.; Kim, Y.H.; Kim, D.K.; Hong, S.P.; Jin, J.S.; Lee, Y.M. Anti-obesity effect of Triticum aestivum sprout extract in high-fat-diet-induced obese mice. Biosci. Biotechnol. Biochem. 2015, 79, 1133–1140. [Google Scholar] [CrossRef] [Green Version]
- Kang, N.E.; Ha, A.W.; Woo, H.W.; Kim, W.K. Peanut sprouts extract (Arachis hypogaea L.) has anti-obesity effects by controlling the protein expressions of PPARgamma and adiponectin of adipose tissue in rats fed high-fat diet. Nutr. Res. Pract. 2014, 8, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Marton, M.; Mandoki, Z.; Csapo-Kiss, Z.; Csapo, J. The role of sprouts in human nutrition. A review. Acta Univ. Sapientiae 2010, 3, 81–117. [Google Scholar]
- Lee, Y.H.; Kim, J.H.; Kim, S.H.; Oh, J.Y.; Seo, W.D.; Kim, K.M.; Jung, J.C.; Jung, Y.S. Barley sprouts extract attenuates alcoholic fatty liver injury in mice by reducing inflammatory response. Nutrients 2016, 8, 440. [Google Scholar] [CrossRef] [Green Version]
- Byun, A.R.; Chun, H.; Lee, J.; Lee, S.W.; Lee, H.S.; Shim, K.W. Effects of a dietary supplement with barley sprout extract on blood cholesterol metabolism. Evid.-Based Complement. Alternat. Med. 2015, 2015, 473056. [Google Scholar] [CrossRef] [Green Version]
- Seo, W.D.; Yuk, H.J.; Curtis-Long, M.J.; Jang, K.C.; Lee, J.H.; Han, S.I.; Kang, H.W.; Nam, M.H.; Lee, S.J.; Lee, J.H.; et al. Effect of the growth stage and cultivar on policosanol profiles of barley sprouts and their adenosine 5′-monophosphate-activated protein kinase activation. J. Agric. Food Chem. 2013, 61, 1117–1123. [Google Scholar] [CrossRef]
- Lee, J.H.; Jia, Y.; Thach, T.T.; Han, Y.; Kim, B.; Wu, C.; Kim, Y.; Seo, W.D.; Lee, S.J. Hexacosanol reduces plasma and hepatic cholesterol by activation of AMP-activated protein kinase and suppression of sterol regulatory element-binding protein-2 in HepG2 and C57BL/6J mice. Nutr. Res. 2017, 43, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Poti, F.; Santi, D.; Spaggiari, G.; Zimetti, F.; Zanotti, I. Polyphenol health effects on cardiovascular and neurodegenerative disorders: A review and meta-analysis. Int. J. Mol. Sci. 2019, 20, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arranz, S.; Chiva-Blanch, G.; Valderas-Martinez, P.; Medina-Remon, A.; Lamuela-Raventos, R.M.; Estruch, R. Wine, beer, alcohol and polyphenols on cardiovascular disease and cancer. Nutrients 2012, 4, 759–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, K.H.; Park, M.J.; Ra, J.E.; Han, S.I.; Nam, M.H.; Kim, J.H.; Lee, J.H.; Seo, W.D. Saponarin from barley sprouts inhibits NF-kappaB and MAPK on LPS-induced RAW 264.7 cells. Food Funct. 2014, 5, 3005–3013. [Google Scholar] [CrossRef] [PubMed]
- Vitcheva, V.; Simeonova, R.; Krasteva, I.; Yotova, M.; Nikolov, S.; Mitcheva, M. Hepatoprotective effects of saponarin, isolated from Gypsophila trichotoma Wend. on cocaine-induced oxidative stress in rats. Redox Rep. 2011, 16, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Simeonova, R.; Vitcheva, V.; Krasteva, I.; Zdraveva, P.; Konstantinov, S.; Ionkova, I. Antidiabetic and antioxidant effects of saponarin from Gypsophila trichotoma on streptozotocin-induced diabetic normotensive and hypertensive rats. Phytomedicine 2016, 23, 483–490. [Google Scholar] [CrossRef]
- Simeonova, R.; Vitcheva, V.; Kondeva-Burdina, M.; Krasteva, I.; Manov, V.; Mitcheva, M. Hepatoprotective and antioxidant effects of saponarin, isolated from Gypsophila trichotoma Wend. on paracetamol-induced liver damage in rats. Biomed. Res. Int. 2013, 2013, 757126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basile, A.; Giordano, S.; López-Sáez, J.A.; Cobianchi, R.C. Antibacterial activity of pure flavonoids isolated from mosses. Phytochemistry 1999, 52, 1479–1482. [Google Scholar] [CrossRef]
- Kim, J.S.; Jeong, E.; Jo, S.M.; Park, J.; Kim, J.Y. Comparative study of the effects of light controlled germination conditions on saponarin content in barley sprouts and lipid accumulation suppression in HepG2 hepatocyte and 3T3-L1 adipocyte cells using barley sprout extracts. Molecules 2020, 25, 5349. [Google Scholar] [CrossRef]
- Engin, A. Fat Cell and Fatty Acid Turnover in Obesity. Adv. Exp. Med. Biol. 2017, 960, 135–160. [Google Scholar] [CrossRef]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Cristancho, A.G.; Lazar, M.A. Forming functional fat: A growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 2011, 12, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.K.; Marcinko, K.; Desjardins, E.M.; Lally, J.S.; Ford, R.J.; Steinberg, G.R. Treatment of nonalcoholic fatty liver disease: Role of AMPK. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E730–E740. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, V.L.; MacDonald, P.E.; Klip, A. The cell biology of systemic insulin function. J. Cell Biol. 2018, 217, 2273–2289. [Google Scholar] [CrossRef] [Green Version]
- Carpene, C.; Les, F.; Casedas, G.; Peiro, C.; Fontaine, J.; Chaplin, A.; Mercader, J.; Lopez, V. Resveratrol anti-obesity effects: Rapid inhibition of adipocyte glucose utilization. Antioxidants 2019, 8, 74. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashida, H.; Furuyashiki, T.; Nagayasu, H.; Bessho, H.; Sakakibara, H.; Hashimoto, T.; Kanazawa, K. Anti-obesity actions of green tea: Possible involvements in modulation of the glucose uptake system and suppression of the adipogenesis-related transcription factors. Biofactors 2004, 22, 135–140. [Google Scholar] [CrossRef]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Ann. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.H.; Jang, K.W.; Park, M.H.; Myung, C.S. Garcinia cambogia suppresses adipogenesis in 3T3-L1 cells by inhibiting p90RSK and Stat3 activation during mitotic clonal expansion. J. Cell Physiol. 2021, 236, 1822–1839. [Google Scholar] [CrossRef]
- Corey, R.; Werner, K.T.; Singer, A.; Moss, A.; Smith, M.; Noelting, J.; Rakela, J. Acute liver failure associated with Garcinia cambogia use. Ann. Hepatol. 2016, 15, 123–126. [Google Scholar] [CrossRef]
- Ferreira, V.; Mathieu, A.; Soucy, G.; Giard, J.M.; Erard-Poinsot, D. Acute severe liver injury related to long-term garcinia cambogia intake. ACG Case Rep. J. 2020, 7, e00429. [Google Scholar] [CrossRef] [PubMed]
- Kothadia, J.P.; Kaminski, M.; Samant, H.; Olivera-Martinez, M. Hepatotoxicity associated with use of the weight loss supplement garcinia cambogia: A case report and review of the literature. Case Rep. Hepatol. 2018, 2018, 6483605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemieux, I.; Lamarche, B.; Couillard, C.; Pascot, A.; Cantin, B.; Bergeron, J.; Dagenais, G.R.; Després, J.P. Total cholesterol/HDL cholesterol ratio vs LDL cholesterol/HDL cholesterol ratio as indices of ischemic heart disease risk in men: The Quebec Cardiovascular Study. Arch. Intern. Med. 2001, 161, 2685–2692. [Google Scholar] [CrossRef]
- Derosa, G.; Maffioli, P. Anti-obesity drugs: A review about their effects and their safety. Expert Opin. Drug Saf. 2012, 11, 459–471. [Google Scholar] [CrossRef]
- Son, J.W.; Kim, S. Comprehensive review of current and upcoming anti-obesity drugs. Diabetes Metab. J. 2020, 44, 802–818. [Google Scholar] [CrossRef]
- Park, B.; Lee, S.; Lee, B.; Kim, I.; Baek, N.; Lee, T.H.; Lee, S.Y.; Son, M.; Park, H. New ethanol extraction improves the anti-obesity effects of black tea. Arch. Pharm. Res. 2016, 39, 310–320. [Google Scholar] [CrossRef]
- Onyebuchi, C.; Kavaz, D. Effect of extraction temperature and solvent type on the bioactive potential of Ocimum gratissimum L. extracts. Sci. Rep. 2020, 10, 21760. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kim, D.S. The extraction solvent influences the anti-inflammatory effects of jakyakgamcho-tang in lipopolysaccharide-stimulated macrophages and mice with gouty arthritis. Int J. Mol. Sci. 2020, 21, 9748. [Google Scholar] [CrossRef]
- Kolb, H.; Kempf, K.; Rohling, M.; Martin, S. Insulin: Too much of a good thing is bad. BMC Med. 2020, 18, 224. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, D.S.; Ebbeling, C.B. The carbohydrate-insulin model of obesity: Beyond “calories in, calories out”. JAMA Intern. Med. 2018, 178, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Seidell, J.C. Carbohydrate intake and obesity. Eur. J. Clin. Nutr. 2007, 61, S75–S99. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Thakur, G.; Kumar, B.D.; Mitra, A.; Chakraborty, C. Long-term effects of a carbohydrate-rich diet on fasting blood sugar, lipid profile, and serum insulin values in rural Bengalis. J. Diabetes 2009, 1, 288–295. [Google Scholar] [CrossRef]
- Borer, K.T. Understanding human physiological limitations and societal pressures in favor of overeating helps to avoid obesity. Nutrients 2019, 11, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.C.; Lindeman, A.K.; Wallace, J.; Niederpruem, M. Diet composition, energy intake, and exercise in relation to body fat in men and women. Am. J. Clin. Nutr. 1990, 52, 426–430. [Google Scholar] [CrossRef]
- Koh, Y.M.; Jang, S.W.; Ahn, T.W. Anti-obesity effect of Yangkyuksanwha-tang in high-fat diet-induced obese mice. BMC Complement. Altern. Med. 2019, 19, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandeya, P.R.; Lamichhane, R.; Lee, K.H.; Lamichhane, G.; Kim, S.G.; Jung, H.J. Efficacy of a novel herbal formulation (F2) on the management of obesity: In vitro and in vivo study. Evid.-Based Complement. Alternat. Med. 2021, 2021, 8854915. [Google Scholar] [CrossRef]
- Lee, C.L.; Wen, J.Y.; Hsu, Y.W.; Pan, T.M. Monascus-fermented yellow pigments monascin and ankaflavin showed antiobesity effect via the suppression of differentiation and lipogenesis in obese rats fed a high-fat diet. J. Agric. Food Chem. 2013, 61, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Achari, A.E.; Jain, S.K. Adiponectin, a therapeutic target for obesity, diabetes, and endothelial dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, G.J.; Azzara, A.V.; Heaner, M.K. Roles for central leptin receptors in the control of meal size. Appetite 2013, 71, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Tryon, R.R.; Horn, W.F.; Welch, L.; Keim, N.L. Estradiol, SHBG and leptin interplay with food craving and intake across the menstrual cycle. Physiol. Behav. 2016, 165, 304–312. [Google Scholar] [CrossRef]
- Pan, H.; Guo, J.; Su, Z. Advances in understanding the interrelations between leptin resistance and obesity. Physiol. Behav. 2014, 130, 157–169. [Google Scholar] [CrossRef]
- Benetolo, P.O.; Fernandes, M.I.M.; Ciampo, I.; Elias-Junior, J.; Sawamura, R. Evaluation of nonalcoholic fatty liver disease using magnetic resonance in obese children and adolescents. J. Pediatr. 2019, 95, 34–40. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | NFD 1 | HFD 2 | Gar 3 | 25 4 | 50 5 | 100 6 |
|---|---|---|---|---|---|---|
| Weight (g) | ||||||
| 1 week (start supplement) | 22.86 ± 0.63 | 21.52 ± 0.29 | 21.48 ± 0.23 | 22.3 ± 0.74 | 22.17 ± 0.24 | 20.90 ± 0.45 |
| 9 weeks (end supplement) | 26.77 ± 0.30 | 36.87 ± 1.71 b | 32.78 ± 1.09 b | 34.48 ± 0.95 b | 33.60 ± 0.59 b | 31.37 ± 0.42 bc |
| Weight gain (g) | 3.90 ± 0.72 | 15.35 ± 1.48 b | 11.30 ± 1.01 bc | 12.18 ± 1.07 b | 11.43 ± 0.72 bc | 10.47 ± 0.70 bc |
| Intake | ||||||
| Food intake (g/day) | 2.57 ± 0.62 | 2.11 ± 0.47 b | 1.96 ± 0.7 b | 1.97 ± 0.44 bc | 1.88 ± 0.63 bc | 1.91 ± 0.91 b |
| Energy intake (kcal/day) | 10.3 ± 0.25 | 10.8 ± 0.24 | 10.01 ± 0.36 | 10.05 ± 0.22 c | 9.59 ± 0.32 c | 9.77 ± 0.46 |
| FER 7 | 0.027 ± 0.002 | 0.129 ± 0.012 b | 0.102 ± 0.019 b | 0.110 ± 0.019 b | 0.108 ± 0.016 b | 0.097 ± 0.023 bc |
| Tissue weight (g) | ||||||
| Liver | 1.10 ± 0.04 | 0.97 ± 0.05 b | 0.86 ± 0.02 d | 0.95 ± 0.04 | 0.91 ± 0.02 c | 0.88 ± 0.03 d |
| Abdominal fat tissue | 0.50 ± 0.03 | 1.94 ± 0.14 b | 1.72 ± 0.12 b | 1.85 ± 0.12 b | 1.57 ± 0.06 b | 1.40 ± 0.06 bc |
| Epididymal fat tissue | 0.11 ± 0.01 b | 0.37 ± 0.03 b | 0.31 ± 0.02 b | 0.28 ± 0.01 bc | 0.29 ± 0.02 b | 0.24 ± 0.01 bd |
| Visceral fat tissue | 0.71 ± 0.05 b | 1.88 ± 0.18 b | 1.42 ± 0.08 bc | 1.60 ± 0.11 b | 1.52 ± 0.05 b | 1.33 ± 0.04 bc |
| Subcutaneous fat tissue | 0.71 ± 0.07 b | 2.96 ± 0.36 b | 2.17 ± 0.15 b | 2.21 ± 0.24 b | 1.97 ± 0.08 bc | 1.58 ± 0.09 bd |
| Measurements | NFD 1 | HFD 2 | Gar 3 | 25 4 | 50 5 | 100 6 |
|---|---|---|---|---|---|---|
| ALT (U/L) | 38.75 ± 1.25 | 84.25 ± 3.54 b | 55 ± 3.11 b d | 75 ± 7.55 b | 68.5 ± 9.42 a | 57.5 ± 7.03 ac |
| AST (U/L) | 25 ± 0.58 | 73.5 ± 9.51 b | 32.5 ± 4.17 d | 49.5 ± 8.97 a | 56.75 ± 15.6 | 34.25 ± 6.3 c |
| Glucose (mg/dL) | 194 ± 13.68 | 304 ± 18.06 b | 259.5 ± 7.23 b | 223.5 ± 11.72 c | 244.75 ± 18.29 | 242 ± 14.16 c |
| Total-Cholesterol (mg/dL) | 85.5 ± 1.04 | 153.5 ± 6.22 b | 138.5 ± 4.91 b | 133.25 ± 9.72 b | 141 ± 3.76 b | 134.25 ± 2.66 bc |
| Triglyceride (mg/dL) | 98.25 ± 4.87 | 117.75 ± 5.89 a | 98.25 ± 3.84 c | 108.5 ± 12.87 | 100.25 ± 10.49 | 89.5 ± 5.42 c |
| HDL (mg/dL) | 67.25 ± 3.38 | 89.25 ± 3.28 b | 88 ± 1.41 b | 83.5 ± 4.41 a | 95.25 ± 2.69 b | 85.5 ± 1.19 b |
| LDL (mg/dL) | 8.75 ± 0.25 | 11.5 ± 0.5 b | 9.75 ± 0.25 ac | 11 ± 0.82 a | 12.75 ± 0.25 b | 10.25 ± 0.48 a |
| Atherodenic index 7 | 1.28 ± 0.03 | 1.73 ± 0.1 b | 1.58 ± 0.05 a | 1.59 ± 0.04 b | 1.48 ± 0.04 a | 1.57 ± 0.05 a |
| Time (min) | Mobile Phase A (%) 1 | Mobile Phase B (%) 2 |
|---|---|---|
| 0 | 80 | 20 |
| 35 | 60 | 40 |
| 40 | 5 | 95 |
| 45 | 5 | 95 |
| 46 | 80 | 20 |
| 50 | 80 | 20 |
| Ingredient (g/kg) | Normal-Fat Diet | High-Fat Diet |
|---|---|---|
| Casein | 200 | 265.0 |
| L-cysteine | 3 | 4 |
| Corn starch | 150 | - |
| Maltodextrin | - | 160 |
| Sucrose | 500 | 90 |
| Cellulose | 50 | 65.5 |
| Soybean oil | 50 | 30 |
| Lard | - | 310 |
| Mineral mixture | 35 | 48 |
| Vitamin mixture | 10 | 21 |
| Choline bitartrate | 2 | 3 |
| Energy (kcal/g) | 4 | 5.1 |
| Blue food color | - | 0.1 |
| Protein (% kcal) | 20 | 18.3 |
| Carbohydrate (% kcal) | 64 | 21.4 |
| Fat (% kcal) | 16 | 60.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-J.; Kawk, H.-W.; Kim, S.-H.; Lee, H.-J.; Seo, J.-W.; Kim, J.-T.; Jang, S.-H.; Kim, M.-J.; Kim, Y.-M. Anti-Obesity Effect of Hot Water Extract of Barley Sprout through the Inhibition of Adipocyte Differentiation and Growth. Metabolites 2021, 11, 610. https://doi.org/10.3390/metabo11090610
Kim M-J, Kawk H-W, Kim S-H, Lee H-J, Seo J-W, Kim J-T, Jang S-H, Kim M-J, Kim Y-M. Anti-Obesity Effect of Hot Water Extract of Barley Sprout through the Inhibition of Adipocyte Differentiation and Growth. Metabolites. 2021; 11(9):610. https://doi.org/10.3390/metabo11090610
Chicago/Turabian StyleKim, Myeong-Jin, Hye-Won Kawk, Sang-Hyeon Kim, Hyo-Jae Lee, Ji-Won Seo, Jong-Tae Kim, Seung-Hee Jang, Min-Jeong Kim, and Young-Min Kim. 2021. "Anti-Obesity Effect of Hot Water Extract of Barley Sprout through the Inhibition of Adipocyte Differentiation and Growth" Metabolites 11, no. 9: 610. https://doi.org/10.3390/metabo11090610
APA StyleKim, M.-J., Kawk, H.-W., Kim, S.-H., Lee, H.-J., Seo, J.-W., Kim, J.-T., Jang, S.-H., Kim, M.-J., & Kim, Y.-M. (2021). Anti-Obesity Effect of Hot Water Extract of Barley Sprout through the Inhibition of Adipocyte Differentiation and Growth. Metabolites, 11(9), 610. https://doi.org/10.3390/metabo11090610