
Extraction and Identification of Volatile Organic Compounds Emitted by Fragrant Flowers of Three Tillandsia Species by HS-SPME/GC-MS


 , , and
, , and 
Abstract
:
1. Introduction
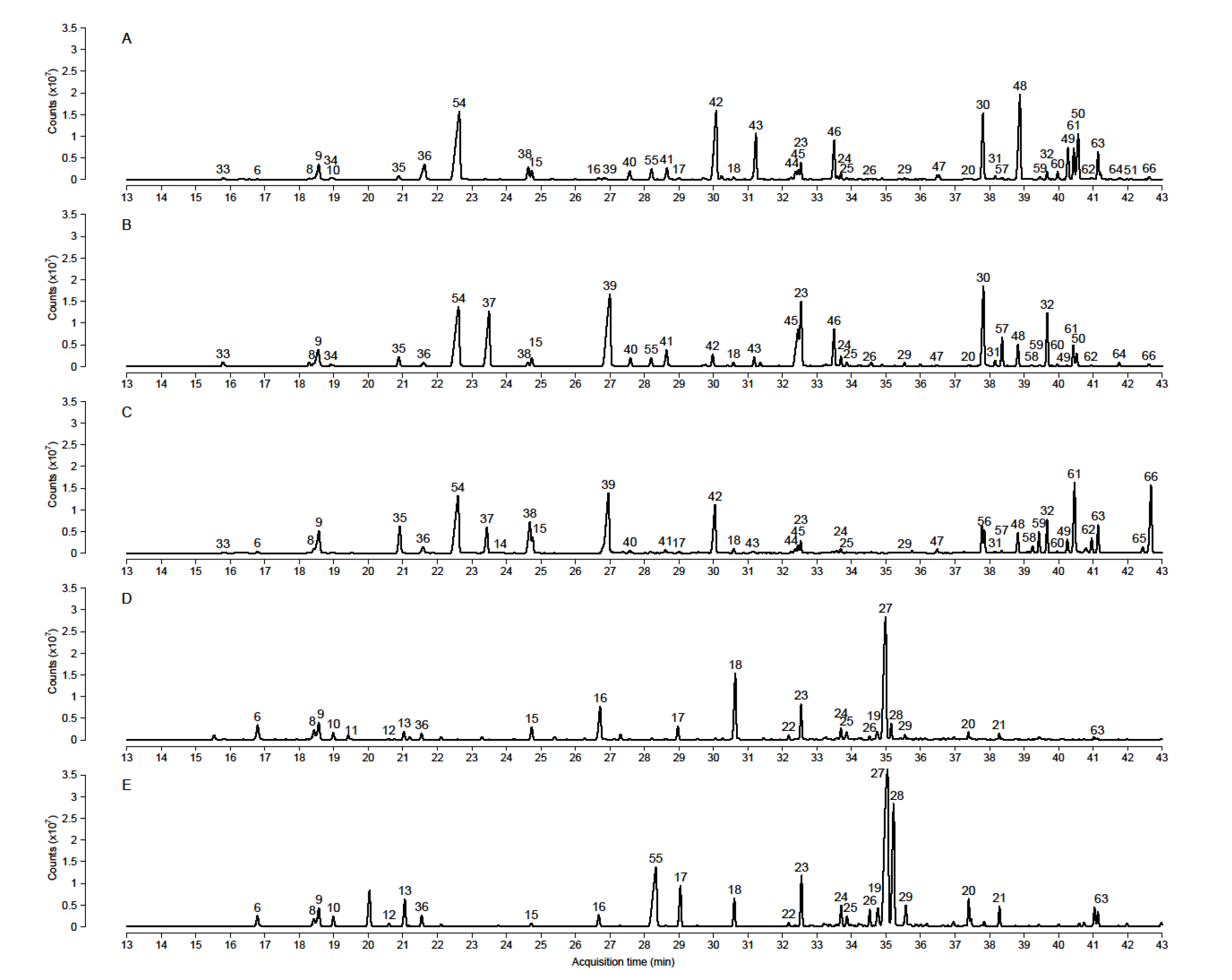
2. Results
2.1. Identification
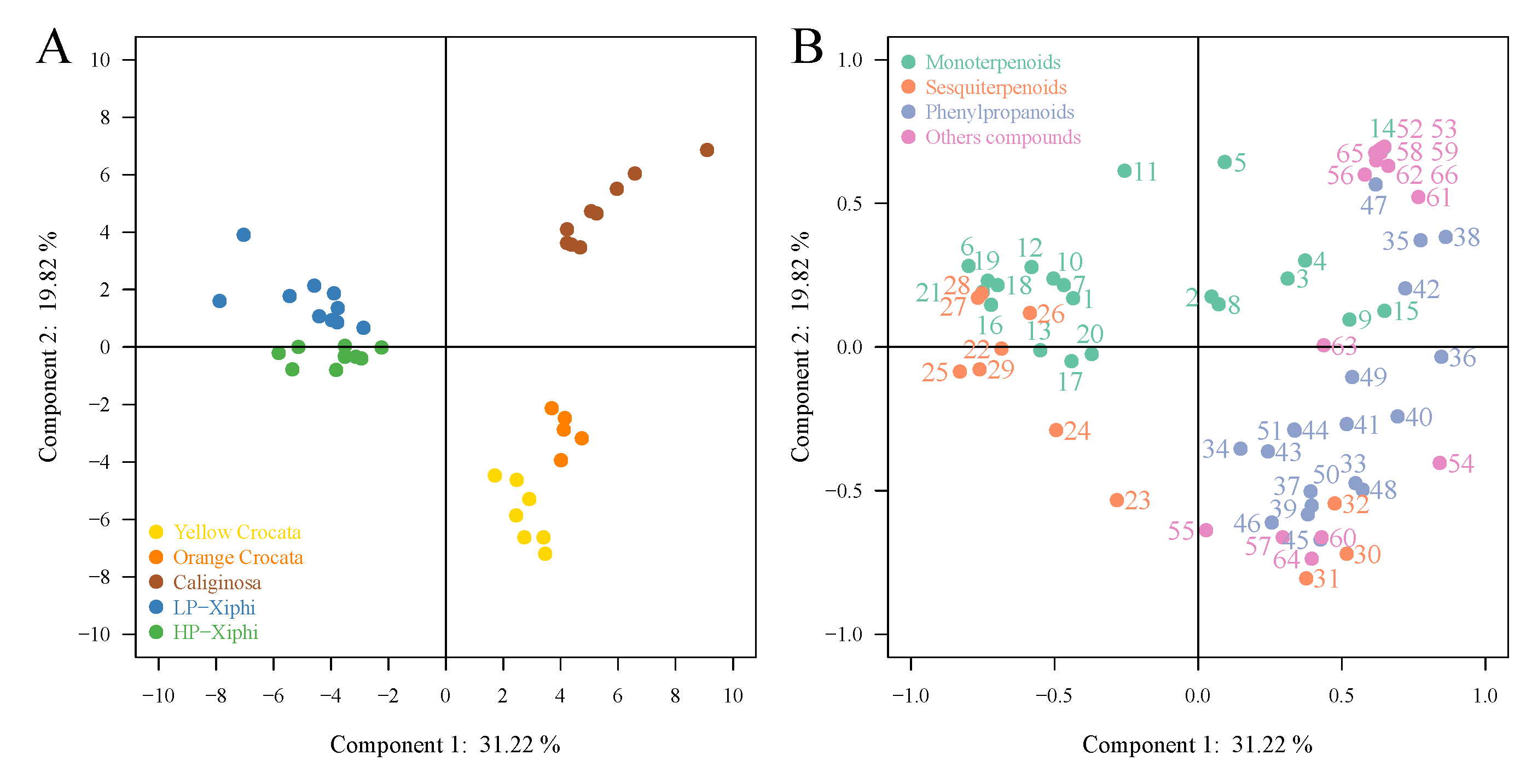
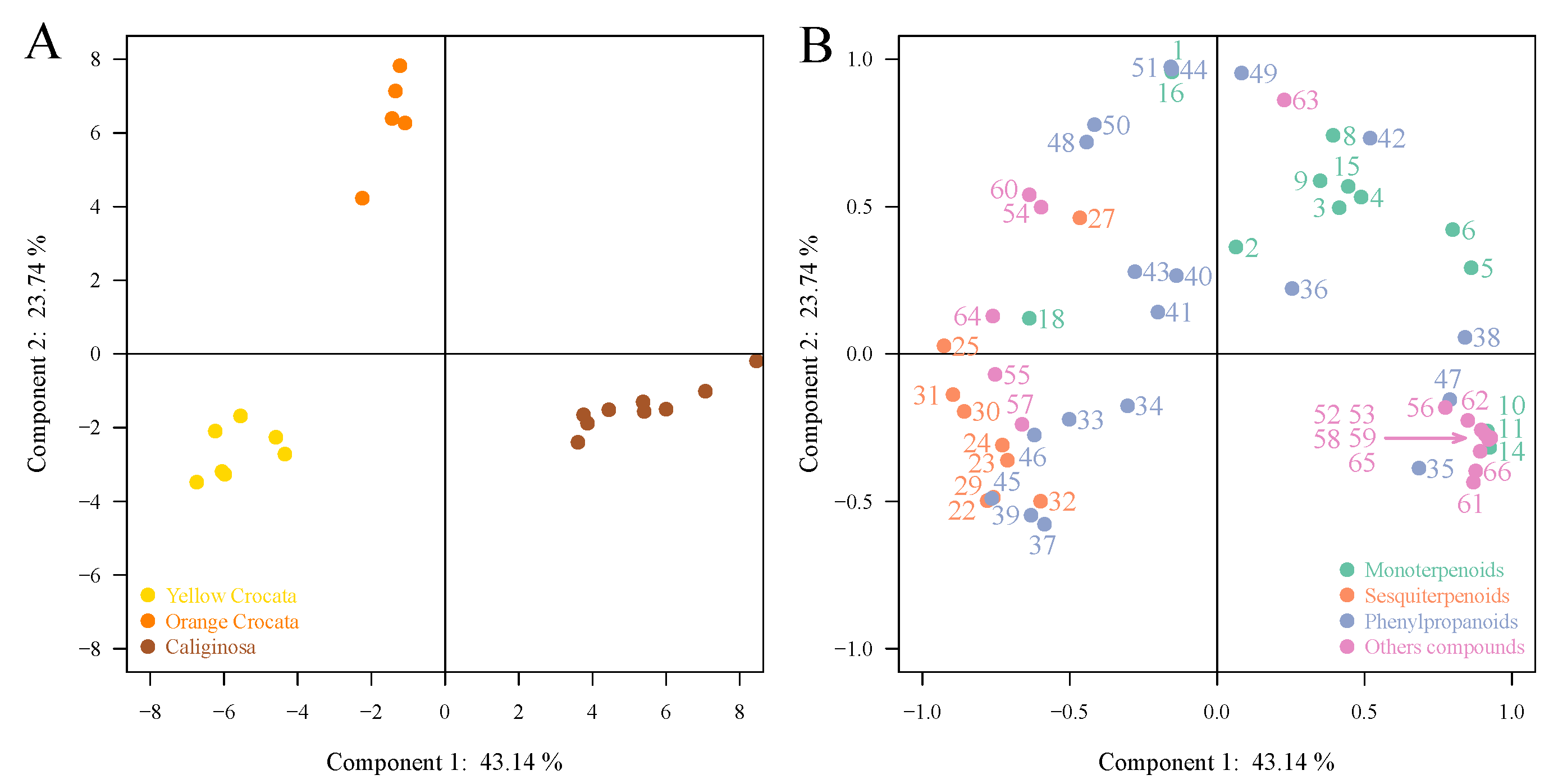
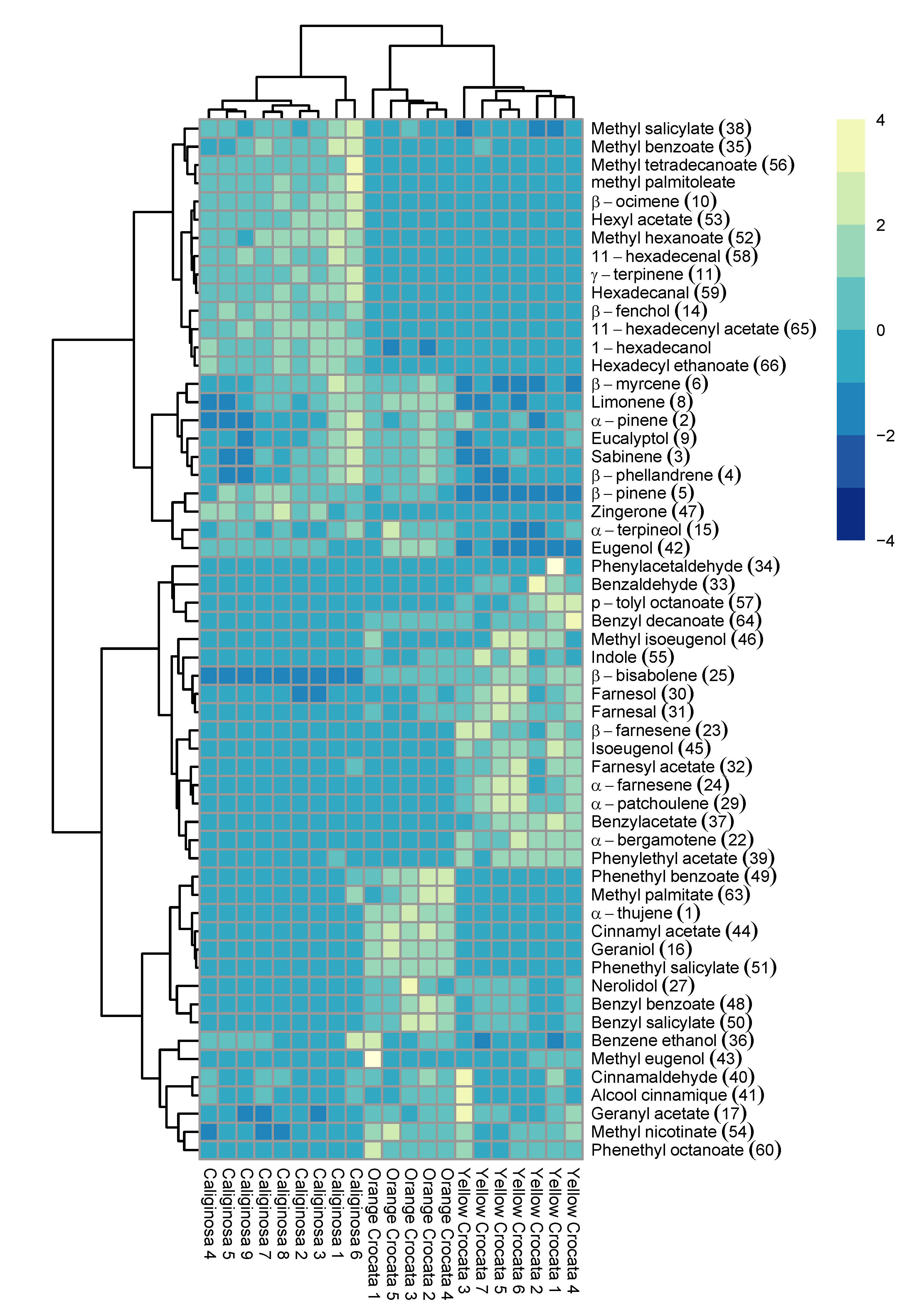
2.2. Chemometrics Analysis of Volatile Organic Compounds
3. Discussion
4. Materials and Methods
4.1. Plant Material and Chemicals
4.2. HS-SPME Conditions
4.3. Instrumentation and GC-MS Conditions
4.4. Identification
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. CRC. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Vivaldo, G.; Masi, E.; Taiti, C.; Caldarelli, G.; Mancuso, S. The network of plants volatile organic compounds. Sci. Rep. 2017, 7, 11050. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Yu, C.; Cheng, B.; Wan, H.; Luo, L.; Pan, H.; Zhang, Q. Volatile compound analysis and aroma evaluation of tea-scented roses in China. Ind. Crops Prod. 2020, 155, 112735. [Google Scholar] [CrossRef]
- Ueda, H.; Kikuta, Y.; Matsuda, K. Plant communication. Plant Signal. Behav. 2012, 7, 222–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F.; Wang, T.; Fan, J.; Liu, Z.; Zong, J.; Fan, W.; Han, Y.; Grierson, D. Volatile composition and classification of Lilium flower aroma types and identification, polymorphisms, and alternative splicing of their monoterpene synthase genes. Hortic. Res. 2019, 6, 110. [Google Scholar] [CrossRef] [Green Version]
- Báez, D.; Pino, J.A.; Morales, D. Floral scent composition in hedychium coronarium J. Koenig Analyzed by SPME. J. Essent. Oil Res. 2011, 23, 64–67. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents—A checklist of volatile compounds isolated by headspace techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, K.; Kim, N.-S.; Lee, D.-S. Determination of floral fragrances of Rosa hybrida using solid-phase trapping-solvent extraction and gas chromatography–mass spectrometry. J. Chromatogr. A 2000, 902, 389–404. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R. Sampling flower scent for chromatographic analysis. J. Sep. Sci. 2008, 31, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wang, J.; Li, D. Microextraction techniques for the determination of volatile and semivolatile organic compounds from plants: A review. Anal. Chim. Acta 2013, 799, 8–22. [Google Scholar] [CrossRef]
- Lo, M.-M.; Benfodda, Z.; Bénimélis, D.; Fontaine, J.-X.; Molinié, R.; Meffre, P. Development of a HS-SPME/GC-MS method for the extraction and identification of the volatile compounds emitted by flowers of tillandsia xiphioides. ACS Omega 2021, 6, 12691–12698. [Google Scholar] [CrossRef]
- Estrella-parra, E.; Flores-cruz, M.; Blancas-flores, G. The Tillandsia genus: History, uses, chemistry, and biological activity. Bol. Latinoam. y del Caribe Plantas Med. y Aromáticas 2019, 18, 239–264. [Google Scholar]
- De Vasconcelos, A.L.; De Vasconcelos, A.L.; Ximenes, E.A.; Randau, K.P. Tillandsia recurvata L. (Bromeliaceae): A pharmacognostic study. Rev. Ciências Farm. Básica Apl. 2015, 34, 151–159. [Google Scholar]
- Gouda, E.J. Tillandsia Bromeliaceae; Eggli, U., Nyffeler, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; ISBN 978-3-662-56484-4. [Google Scholar]
- Gerlach, G.; Schill, R. Composition of orchid scents attracting euglossine bees. Bot. Acta 1991, 104, 379–384. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E. Risk, trust, and consumer acceptance of plant biotechnology: Implications for genetically modified ornamental plants. J. Crop. Improv. 2006, 18, 451–486. [Google Scholar] [CrossRef]
- Roguenant, A. Les Tillandsia et les Racinaea, 1st ed.; Belin: Paris, France, 2001; pp. 221–304. [Google Scholar]
- Burkle, L.A.; Runyon, J.B. The Smell of environmental change: Using floral scent to explain shifts in pollinator attraction. Appl. Plant. Sci. 2017, 5, 1600123. [Google Scholar] [CrossRef]
- Bernardello, L.M.; Galetto, L.; Juliani, H.R. Floral nectar, nectary structure and pollinators in some argentinean bromeliaceae. Ann. Bot. 1991, 67, 401–411. [Google Scholar] [CrossRef]
- Knudsen, J. Trends in floral scent chemistry in pollination syndromes: Floral scent composition in moth-pollinated taxa. Bot. J. Linn. Soc. 1993, 113, 263–284. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.L.; Gómez, M.; Murcia, M.D.; Gómez, E.; Hidalgo, A.M.; Montiel, C.; Martínez, R. Biosynthesis of benzyl acetate: Optimization of experimental conditions, kinetic modelling and application of alternative methods for parameters determination. Bioresour. Technol. Rep. 2020, 11, 100519. [Google Scholar] [CrossRef]
- Kuo, C.H.; Chiang, S.H.; Ju, H.Y.; Chen, Y.M.; Liao, M.Y.; Liu, Y.C.; Shieh, C.J. Enzymatic synthesis of rose aromatic ester (2-phenylethyl acetate) by lipase. J. Sci. Food Agric. 2012, 92, 2141–2147. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Park, C. Enzymatic synthesis of phenethyl ester from phenethyl alcohol with acyl donors. Enzym. Microb. Technol. 2017, 100, 37–44. [Google Scholar] [CrossRef]
- Li, P.; Peng, Y.; Mei, J.; Xie, J. Effects of microencapsulated eugenol emulsions on microbiological, chemical and organoleptic qualities of farmed Japanese sea bass (Lateolabrax japonicus) during cold storage. LWT 2020, 118, 108831. [Google Scholar] [CrossRef]
- Issa, M.Y.; Mohsen, E.; Younis, I.Y.; Nofal, E.S.; Farag, M.A. Volatiles distribution in jasmine flowers taxa grown in Egypt and its commercial products as analyzed via solid-phase microextraction (SPME) coupled to chemometrics. Ind. Crops Prod. 2020, 144, 112002. [Google Scholar] [CrossRef]
- Smith, R.; Adams, T.; Doull, J.; Feron, V.; Goodman, J.; Marnett, L.; Portoghese, P.; Waddell, W.; Wagner, B.; Rogers, A.; et al. Safety assessment of allylalkoxybenzene derivatives used as flavouring substances—methyl eugenol and estragole. Food Chem. Toxicol. 2002, 40, 851–870. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Chen, F.; Pichersky, E. Characterization of an acyltransferase capable of synthesizing benzylbenzoate and other volatile esters in flowers and damaged leaves of clarkia breweri. Plant. Physiol. 2002, 130, 466–476. [Google Scholar] [CrossRef] [Green Version]
- Boatright, J.; Negre, F.; Chen, X.; Kish, C.M.; Wood, B.; Peel, G.; Orlova, I.; Gang, D.; Rhodes, D.; Dudareva, N. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 2004, 135, 1993–2011. [Google Scholar] [CrossRef] [Green Version]
- Lapczynski, A.; McGinty, D.; Jones, L.; Bhatia, S.; Letizia, C.S.; Api, A.M. Fragrance material review on benzyl salicylate. Food Chem. Toxicol. 2007, 45, 362–380. [Google Scholar] [CrossRef]
- Zhu, L.-X.; Zhang, M.-M.; Xiang, X.-F.; Lan, Y.-B.; Shi, Y.; Duan, C.; Zhang, R.-L. Aromatic characterization of traditional Chinese wine Msalais by partial least-square regression analysis based on sensory quantitative descriptive and odor active values, aroma extract dilution analysis, and aroma recombination and omission tests. Food Chem. 2021, 361, 129781. [Google Scholar] [CrossRef]
- Dormont, L.; Delle-Vedove, R.; Bessière, J.M.; Schatz, B. Floral scent emitted by white and coloured morphs in orchids. Phytochemistry 2014, 100, 51–59. [Google Scholar] [CrossRef]
- Majetic, C.J.; Raguso, R.A.; Ashman, T.L. The impact of biochemistry vs. population membership on floral scent profiles in colour polymorphic Hesperis matronalis. Ann. Bot. 2008, 102, 911–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Conchou, L.; Bessière, J.-M.; Cazals, G.; Schatz, B.; Imbert, E. Flower color polymorphism in Iris lutescens (Iridaceae): Biochemical analyses in light of plant–insect interactions. Phytochemistry 2013, 94, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Elawa, S.; Mirdell, R.; Tesselaar, E.; Farnebo, S. The microvascular response in the skin to topical application of methyl nicotinate: Effect of concentration and variation between skin sites. Microvasc. Res. 2019, 124, 54–60. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | LP-Xiphi | HP-Xiphi | Yellow crocata | Orange crocata | T. caliginosa |
|---|---|---|---|---|---|
 |  |  |  |  | |
| Height * (mm) | 50 | 50 | 20 | 15 | 15 |
| Breadth * (mm) | 20 | 20 | 5 | 5 | 5 |
| Odor level | Scented | Scented | Scented | Scented | Scented |
| # | Family | Compound | RT (min) | Composition (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Yellow crocata | Orange crocata | T. Caliginosa | LP-Xiphi | HP-Xiphi | ||||
| 1 | M | α-thujene b, d | 14.15 | nd | 0.2 | nd | 0.3 | nd |
| 2 | M | α-pinene c, d | 14.46 | 0.3 | 0.3 | 0.3 | 0.5 | nd |
| 3 | M | Sabinene c, d | 16.05 | 0.2 | 0.3 | 0.4 | 0.3 | nd |
| 4 | M | β-phellandrene b, d | 16.23 | 0.3 | 0.5 | 0.6 | 0.5 | nd |
| 5 | M | β-pinene c, d | 16.46 | nd | 0.2 | 0.4 | 0.3 | nd |
| 6 | M | β-myrcene c, d | 16.78 | 0.5 | 1.2 | 1.6 | 4.7 | 6.6 |
| 7 | M | α-phellandrene b, d | 17.51 | nd | nd | nd | 0.7 | nd |
| 8 | M | Limonene c, d | 18.52 | 6.4 | 11.4 | 10.8 | 10.1 | 11.5 |
| 9 | M | Eucalyptol c, d | 18.66 | 8.1 | 11.5 | 12.1 | 7.2 | 7.9 |
| 10 | M | β-ocimene c, d | 18.98 | nd | nd | 0.4 | 18.8 | 9.3 |
| 11 | M | γ-terpinene c, d | 19.5 | nd | nd | 0.2 | 0.3 | nd |
| 12 | M | Terpinolene b, d | 20.57 | nd | nd | nd | 0.3 | nd |
| 13 | M | β-linalool c, d | 21.02 | nd | nd | nd | 1.6 | 16.1 |
| 14 | M | β-fenchol c, d | 23.8 | nd | nd | 0.3 | nd | nd |
| 15 | M | α-terpineol c, d | 24.71 | 1.2 | 1.9 | 2.1 | 0.8 | 0.9 |
| 16 | M | Geraniol c, d | 26.64 | nd | 0.3 | nd | 1.2 | 1.1 |
| 17 | M | Methyl geranate b, e | 29.02 | nd | nd | nd | nd | 1.9 |
| 18 | M | Geranyl acetate c, e | 30.59 | 0.3 | 0.3 | 0.2 | 19.6 | 9.4 |
| 19 | M | Geranyl butyrate b, e | 34.75 | nd | nd | nd | 1.5 | 1.3 |
| 20 | M | Geranyl tiglate b, e | 37.39 | nd | nd | nd | nd | 1.4 |
| 21 | M | Geranyl hexanoate b, e | 38.27 | nd | nd | nd | 0.9 | 1.2 |
| 22 | S | α-bergamotene b, e | 32.16 | 0.2 | nd | nd | 0.5 | 0.8 |
| 23 | S | β-farnesene b, e | 32.55 | 5.5 | 1.2 | 1.0 | 3.3 | 3.5 |
| 24 | S | α-farnesene b, e | 33.69 | 1.2 | 0.4 | 0.4 | 1.2 | 1.2 |
| 25 | S | β-bisabolene b, e | 33.86 | 0.3 | 0.2 | nd | 0.8 | 1.7 |
| 26 | S | α-bisabolene b, e | 34.52 | nd | nd | nd | 0.5 | 0.9 |
| 27 | S | Nerolidol c, e | 34.98 | 0.3 | 0.4 | nd | 18.7 | 14.6 |
| 28 | S | Denderalasin b, e | 35.17 | nd | nd | nd | 4.7 | 5.0 |
| 29 | S | α-patchoulene b, e | 35.55 | 0.4 | nd | nd | 0.7 | 0.8 |
| 30 | S | Farnesol c, e | 37.81 | 9.3 | 4.3 | 1.8 | nd | nd |
| 31 | S | Farnesal b, e | 38.16 | 0.6 | 0.3 | nd | nd | nd |
| 32 | S | Farnesyl acetate b, e | 39.67 | 4.5 | 0.5 | 1.3 | nd | nd |
| 33 | P | Benzaldehyde c, d | 15.8 | 0.5 | 0.2 | 0.2 | nd | nd |
| 34 | P | Phenylacetaldehyde c, d | 18.92 | 2.3 | 0.3 | nd | nd | nd |
| 35 | P | Methyl benzoate c, d | 20.89 | 0.8 | 0.3 | 2.2 | nd | nd |
| 36 | P | Benzene ethanol b, d | 21.57 | 0.9 | 1.3 | 1.5 | nd | nd |
| 37 | P | Benzylacetate c, d | 23.43 | 5.5 | nd | 1.3 | nd | nd |
| 38 | P | Methyl salicylate c, d | 24.63 | 0.6 | 1.1 | 2.4 | nd | nd |
| 39 | P | Phenylethyl acetate c, d | 26.87 | 14.2 | nd | 2.5 | nd | nd |
| 40 | P | Cinnamaldehyde b, e | 27.5 | 0.5 | 0.6 | 0.5 | nd | nd |
| 41 | P | Alcool cinnamique c, e | 28.61 | 1.2 | 1.2 | 0.8 | nd | nd |
| 42 | P | Eugenol c, e | 29.96 | 0.9 | 14.5 | 12.3 | nd | nd |
| 43 | P | Methyl eugenol c, e | 31.18 | 0.4 | 1.0 | nd | nd | nd |
| 44 | P | Cinnamyl acetate b, e | 32.37 | nd | 0.7 | nd | nd | nd |
| 45 | P | Isoeugenol c, e | 32.45 | 5.1 | 0.6 | 0.9 | nd | nd |
| 46 | P | Methyl isoeugenol b, e | 33.49 | 2.8 | 0.8 | nd | nd | nd |
| 47 | P | Zingerone b, e | 36.5 | nd | 0.5 | 2.7 | nd | nd |
| 48 | P | Benzyl benzoate c, e | 38.83 | 4.5 | 8.9 | 1.8 | nd | nd |
| 49 | P | Phenethyl benzoate b, e | 40.24 | 0.2 | 5.1 | 1.4 | nd | nd |
| 50 | P | Benzyl salicylate b, e | 40.51 | 1.7 | 5.1 | nd | nd | nd |
| 51 | P | Phenethyl salicylate a, e | 42.09 | nd | 0.2 | nd | nd | nd |
| 52 | O | Methyl hexanoate b, d | 14.24 | nd | nd | 0.3 | nd | nd |
| 53 | O | Hexyl acetate c, d | 17.58 | nd | nd | 0.2 | nd | nd |
| 54 | O | Methyl nicotinate c, d | 22.59 | 13.3 | 15.2 | 10.8 | nd | nd |
| 55 | O | Indole b, e | 28.22 | 1.2 | 0.6 | nd | nd | 1.1 |
| 56 | O | Methyl tetradecanoate b, e | 37.85 | nd | nd | 1.7 | nd | nd |
| 57 | O | p-tolyl octanoate b, e | 38.36 | 1.1 | 0.2 | nd | nd | nd |
| 58 | O | 11-hexadecenal b, e | 39.24 | nd | nd | 0.5 | nd | nd |
| 59 | O | Hexadecanal b, e | 39.43 | nd | nd | 1.3 | nd | nd |
| 60 | O | Phenethyl octanoate b, e | 39.97 | 0.3 | 0.4 | nd | nd | nd |
| 61 | O | 1-hexadecanol b, e | 40.42 | 2.0 | 1.0 | 10.2 | nd | nd |
| 62 | O | Methyl palmitoleate a, e | 40.96 | nd | nd | 1.2 | nd | nd |
| 63 | O | Methyl palmitate a, e | 41.14 | nd | 4.3 | 1.7 | nd | 1.7 |
| 64 | O | Benzyl decanoate a, e | 41.76 | 0.3 | 0.2 | nd | nd | nd |
| 65 | O | 11-hexadecenyl acetate a, e | 42.43 | nd | nd | 0.7 | nd | nd |
| 66 | O | Hexadecyl ethanoate a, e | 42.66 | 0.5 | nd | 6.9 | nd | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, M.-M.; Benfodda, Z.; Bénimélis, D.; Fontaine, J.-X.; Molinié, R.; Meffre, P. Extraction and Identification of Volatile Organic Compounds Emitted by Fragrant Flowers of Three Tillandsia Species by HS-SPME/GC-MS. Metabolites 2021, 11, 594. https://doi.org/10.3390/metabo11090594
Lo M-M, Benfodda Z, Bénimélis D, Fontaine J-X, Molinié R, Meffre P. Extraction and Identification of Volatile Organic Compounds Emitted by Fragrant Flowers of Three Tillandsia Species by HS-SPME/GC-MS. Metabolites. 2021; 11(9):594. https://doi.org/10.3390/metabo11090594
Chicago/Turabian StyleLo, Mame-Marietou, Zohra Benfodda, David Bénimélis, Jean-Xavier Fontaine, Roland Molinié, and Patrick Meffre. 2021. "Extraction and Identification of Volatile Organic Compounds Emitted by Fragrant Flowers of Three Tillandsia Species by HS-SPME/GC-MS" Metabolites 11, no. 9: 594. https://doi.org/10.3390/metabo11090594
APA StyleLo, M.-M., Benfodda, Z., Bénimélis, D., Fontaine, J.-X., Molinié, R., & Meffre, P. (2021). Extraction and Identification of Volatile Organic Compounds Emitted by Fragrant Flowers of Three Tillandsia Species by HS-SPME/GC-MS. Metabolites, 11(9), 594. https://doi.org/10.3390/metabo11090594