
Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Fetal Carcass Traits
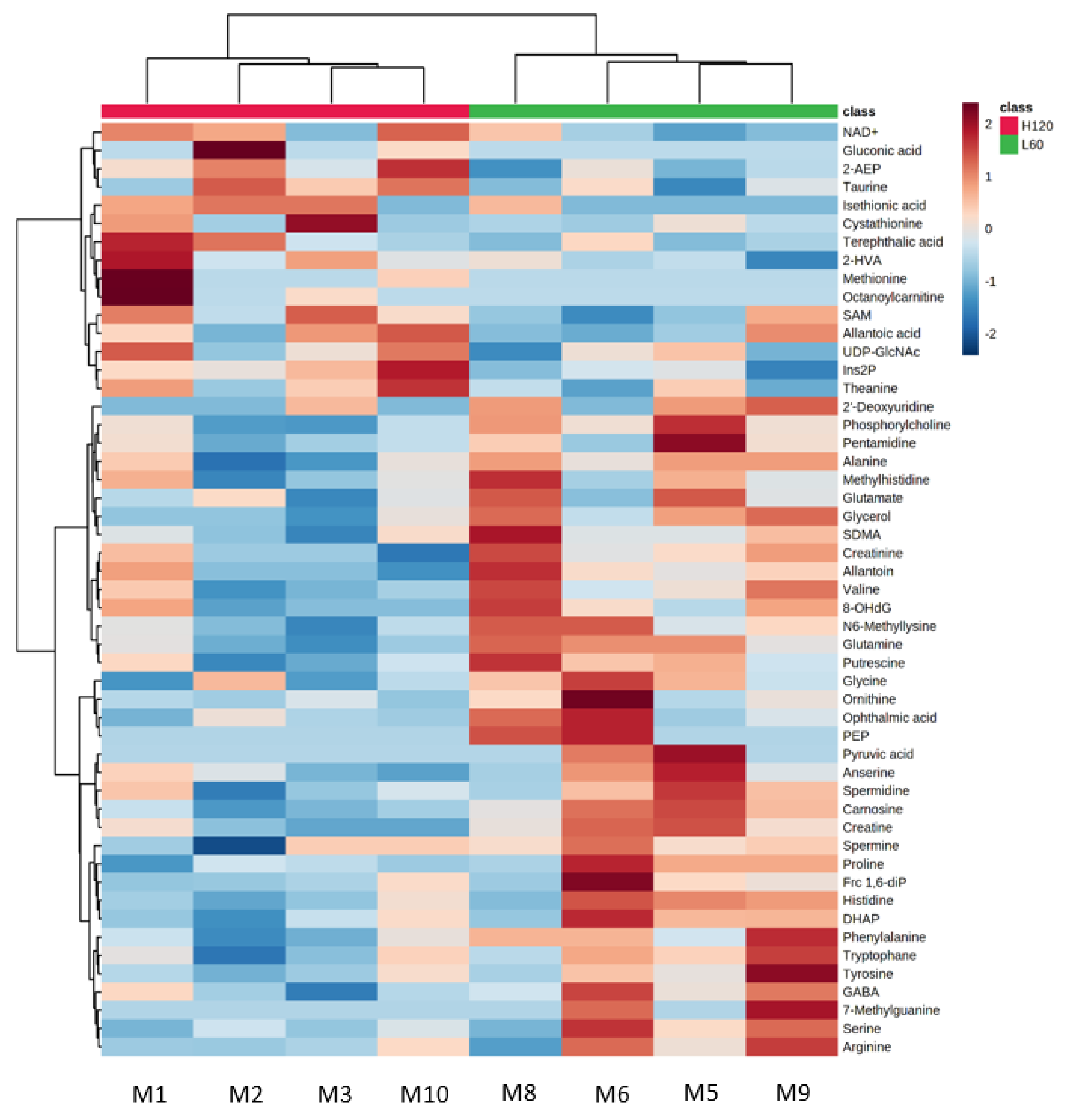
2.2. Metabolomic Profile of Fetal Skeletal Muscle
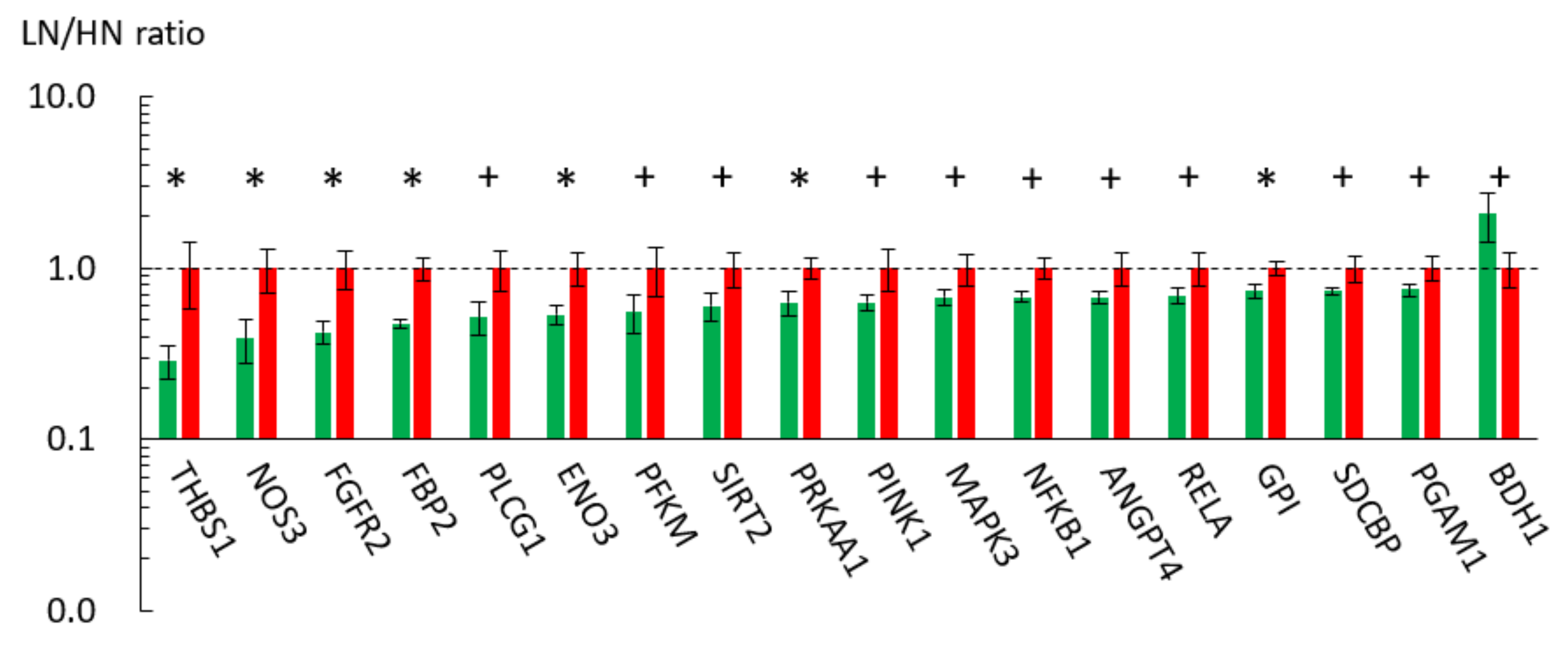
2.3. Effect of MUN on the Gene Expression Profile of the Fetal Skeletal Muscle
2.4. GO Analysis of MUN-Associated Metabolic Pathways
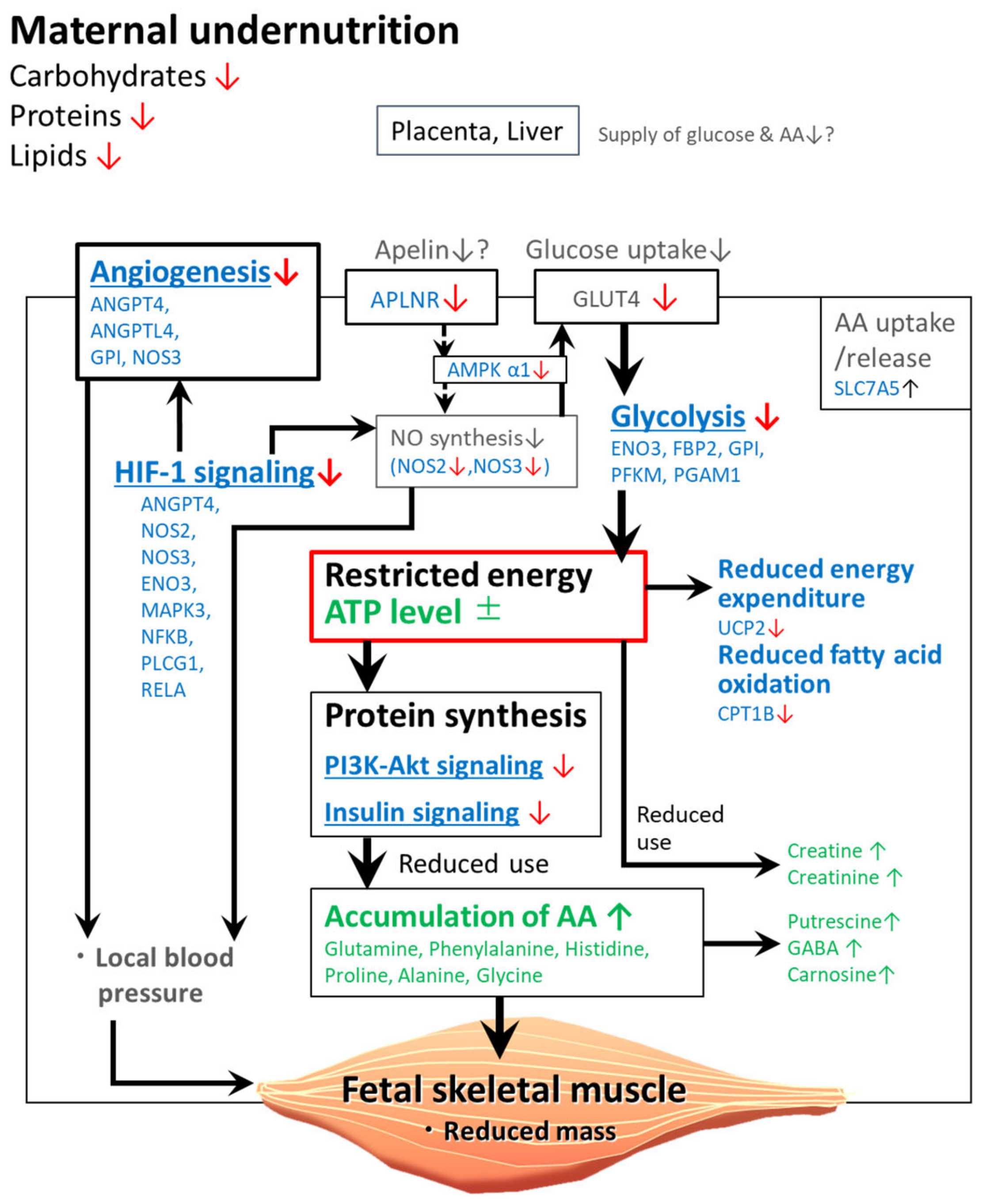
3. Discussion
3.1. Altered Amino Acid Metabolism Was the Major Response to IUGR
3.2. Association of Altered AAs with Metabolism of Nitrogen, β-Alanine, and Glycerophospholipids
3.3. Other Metabolites Altered in the Fetal Muscles of the LN Group
3.4. Potential Mechanisms of NOS Downregulation and Its Impact in the Fetuses of the LN Group
3.5. Downregulated Genes in the Fetuses of the LN Group Are Associated with Energy Metabolism or Angiogenesis
3.6. Altered Expression of Genes Associated with One-Carbon Cycle and Uptake of Large Neutral AAs
3.7. Levels of Nutrients for Dams to Be Compared in the Present Study
4. Materials and Methods
4.1. Animals and Dietary Treatments
4.2. Sample Collection
4.3. Sample Preparation for CE-TOFMS
4.4. Instrumentation and Conditions of CE-TOFMS
4.5. Data Analysis of CE-TOFMS Results
4.6. RNA Preparation and Complementary DNA (cDNA) Synthesis
4.7. Microarray Analysis
4.8. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.9. Functional Annotation of Target Genes
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sandoval, C.; Lambo, C.A.; Beason, K.; Dunlap, K.A.; Satterfield, M.C. Effect of maternal nutrient restriction on skeletal muscle mass and associated molecular pathways in SGA and Non-SGA sheep fetuses. Domest. Anim. Endocrinol. 2020, 72, 106443. [Google Scholar] [CrossRef]
- Yates, D.T.; Macko, A.R.; Nearing, M.; Chen, X.; Rhoads, R.P.; Limesand, S.W. Developmental programming in response to intrauterine growth restriction impairs myoblast function and skeletal muscle metabolism. J. Pregnancy 2012, 2012, 631038. [Google Scholar] [CrossRef]
- Osgerby, J.C.; Wathes, D.C.; Howard, D.; Gadd, T.S. The effect of maternal undernutrition on ovine fetal growth. J. Endocrinol. 2002, 173, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Vonnahme, K.A.; Hess, B.W.; Hansen, T.R.; McCormick, R.J.; Rule, D.C.; Moss, G.E.; Murdoch, W.J.; Nijland, M.J.; Skinner, D.C.; Nathanielsz, P.W.; et al. Maternal undernutrition from early- to mid-gestation leads to growth retardation, cardiac ventricular hypertrophy, and increased liver weight in the fetal sheep. Biol. Reprod. 2003, 69, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, L.A.; Zhang, L.; Tuersunjiang, N.; Ma, Y.; Long, N.M.; Uthlaut, A.B.; Smith, D.T.; Nathanielsz, P.W.; Ford, S.P. Early maternal undernutrition programs increased feed intake, altered glucose metabolism and insulin secretion, and liver function in aged female offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R795–R804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, L.J.; Foster, T.; Rhodes, P.; Rhind, S.M.; Gardner, D.S. Protein-energy malnutrition during early gestation in sheep blunts fetal renal vascular and nephron development and compromises adult renal function. J. Physiol. 2012, 590, 377–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, P.; Ghatta, S.; Dubey, N.; Lemley, C.O.; Johnson, M.L.; Modgil, A.; Vonnahme, K.; Caton, J.S.; Reynolds, L.P.; Sun, C.; et al. Maternal nutrient restriction during pregnancy impairs an endothelium-derived hyperpolarizing factor-like pathway in sheep fetal coronary arteries. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H134–H142. [Google Scholar] [CrossRef] [Green Version]
- Devaskar, S.U.; Chu, A. Intrauterine Growth Restriction: Hungry for an Answer. Physiology 2016, 31, 131–146. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.D. Endocrine regulation of fetal skeletal muscle growth: Impact on future metabolic health. J. Endocrinol. 2014, 221, R13–R29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Otomaru, K.; Oshima, K.; Goto, Y.; Oshima, I.; Muroya, S.; Sano, M.; Saneshima, R.; Nagao, Y.; Kinoshita, A.; et al. Effects of low and high levels of maternal nutrition consumed for the entirety of gestation on the development of muscle, adipose tissue, bone, and the organs of Wagyu cattle fetuses. Anim. Sci. J. 2021, 92, e13600. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ford, S.P.; Nathanielsz, P.W.; Du, M. Effect of maternal nutrient restriction in sheep on the development of fetal skeletal muscle. Biol. Reprod. 2004, 71, 1968–1973. [Google Scholar] [CrossRef] [Green Version]
- Quigley, S.P.; Kleemann, D.O.; Kakar, M.A.; Owens, J.A.; Nattrass, G.S.; Maddocks, S.; Walker, S.K. Myogenesis in sheep is altered by maternal feed intake during the peri-conception period. Anim. Reprod. Sci. 2005, 87, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Widdowson, E.M.; Crabb, D.E.; Milner, R.D. Cellular development of some human organs before birth. Arch. Dis. Child. 1972, 47, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Yates, D.T.; Clarke, D.S.; Macko, A.R.; Anderson, M.J.; Shelton, L.A.; Nearing, M.; Allen, R.E.; Rhoads, R.P.; Limesand, S.W. Myoblasts from intrauterine growth-restricted sheep fetuses exhibit intrinsic deficiencies in proliferation that contribute to smaller semitendinosus myofibres. J. Physiol. 2014, 592, 3113–3125. [Google Scholar] [CrossRef]
- Maier, A.; McEwan, J.C.; Dodds, K.G.; Fischman, D.A.; Fitzsimons, R.B.; Harris, A.J. Myosin heavy chain composition of single fibres and their origins and distribution in developing fascicles of sheep tibialis cranialis muscles. J. Muscle Res. Cell Motil. 1992, 13, 551–572. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ford, S.P.; Means, W.J.; Hess, B.W.; Nathanielsz, P.W.; Du, M. Maternal nutrient restriction affects properties of skeletal muscle in offspring. J. Physiol. 2006, 575, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.T.; Cadaret, C.N.; Beede, K.A.; Riley, H.E.; Macko, A.R.; Anderson, M.J.; Camacho, L.E.; Limesand, S.W. Intrauterine growth-restricted sheep fetuses exhibit smaller hindlimb muscle fibers and lower proportions of insulin-sensitive Type I fibers near term. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R1020–R1029. [Google Scholar] [CrossRef] [Green Version]
- Maresca, S.; Valiente, S.L.; Rodriguez, A.M.; Testa, L.M.; Long, N.M.; Quintans, G.I.; Pavan, E. The influence of protein restriction during mid- to late gestation on beef offspring growth, carcass characteristic and meat quality. Meat Sci. 2019, 153, 103–108. [Google Scholar] [CrossRef]
- Ramírez, M.; Testa, L.M.; López Valiente, S.; Latorre, M.E.; Long, N.M.; Rodriguez, A.M.; Pavan, E.; Maresca, S. Maternal energy status during late gestation: Effects on growth performance, carcass characteristics and meat quality of steers progeny. Meat Sci. 2020, 164, 108095. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Burghardt, R.C.; Johnson, G.A.; Kim, S.W.; Li, X.L.; Satterfield, M.C.; Spencer, T.E. Impacts of amino acid nutrition on pregnancy outcome in pigs: Mechanisms and implications for swine production. J. Anim. Sci. 2010, 88, E195–E204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.E.; Jones, A.K.; Pillai, S.M.; Hoffman, M.L.; McFadden, K.K.; Zinn, S.A.; Govoni, K.E.; Reed, S.A. Maternal Restricted- and Over-Feeding During Gestation Result in Distinct Lipid and Amino Acid Metabolite Profiles in the Longissimus Muscle of the Offspring. Front. Physiol. 2019, 10, 515. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.I.; Wesolowski, S.R.; Gilje, E.A.; Baker, P.R., 2nd; Reisz, J.A.; D’Alessandro, A.; Hay, W.W., Jr.; Rozance, P.J.; Brown, L.D. Skeletal muscle amino acid uptake is lower and alanine production is greater in late gestation intrauterine growth-restricted fetal sheep hindlimb. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 317, R615–R629. [Google Scholar] [CrossRef]
- Dunlop, K.; Cedrone, M.; Staples, J.F.; Regnault, T.R. Altered fetal skeletal muscle nutrient metabolism following an adverse in utero environment and the modulation of later life insulin sensitivity. Nutrients 2015, 7, 1202–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauvin, M.C.; Pillai, S.M.; Reed, S.A.; Stevens, J.R.; Hoffman, M.L.; Jones, A.K.; Zinn, S.A.; Govoni, K.E. Poor maternal nutrition during gestation in sheep alters prenatal muscle growth and development in offspring. J. Anim. Sci. 2020, 98, skz388. [Google Scholar] [CrossRef] [PubMed]
- He, J.; He, Y.; Yu, B.; Wang, X.; Chen, D. Transcriptome Characterization of Repressed Embryonic Myogenesis Due to Maternal Calorie Restriction. Front. Cell Dev. Biol. 2020, 8, 527. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, L.P.; Nascimento, M.; Moriel, P.; Sommer, J.; Ashwell, M.; Poore, M.H.; Duarte, M.S.; Serão, N.V.L. Impact of energy restriction during late gestation on the muscle and blood transcriptome of beef calves after preconditioning. BMC Genom. 2018, 19, 702. [Google Scholar] [CrossRef]
- Sohel, M.M.H.; Akyuz, B.; Konca, Y.; Arslan, K.; Gurbulak, K.; Abay, M.; Kaliber, M.; Cinar, M.U. Differential protein input in the maternal diet alters the skeletal muscle transcriptome in fetal sheep. Mamm. Genome 2020, 31, 309–324. [Google Scholar] [CrossRef]
- Diniz, W.J.S.; Crouse, M.S.; Cushman, R.A.; McLean, K.J.; Caton, J.S.; Dahlen, C.R.; Reynolds, L.P.; Ward, A.K. Cerebrum, liver, and muscle regulatory networks uncover maternal nutrition effects in developmental programming of beef cattle during early pregnancy. Sci. Rep. 2021, 11, 2771. [Google Scholar] [CrossRef]
- Sandoval, C.; Wu, G.; Smith, S.B.; Dunlap, K.A.; Satterfield, M.C. Maternal Nutrient Restriction and Skeletal Muscle Development: Consequences for Postnatal Health. Adv. Exp. Med. Biol. 2020, 1265, 153–165. [Google Scholar] [CrossRef]
- Paradis, F.; Wood, K.M.; Swanson, K.C.; Miller, S.P.; McBride, B.W.; Fitzsimmons, C. Maternal nutrient restriction in mid-to-late gestation influences fetal mRNA expression in muscle tissues in beef cattle. BMC Genom. 2017, 18, 632. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Zhu, M.J.; Means, W.J.; Hess, B.W.; Ford, S.P. Nutrient restriction differentially modulates the mammalian target of rapamycin signaling and the ubiquitin-proteasome system in skeletal muscle of cows and their fetuses. J. Anim. Sci. 2005, 83, 117–123. [Google Scholar] [CrossRef]
- National Agriculture and Food Research Organization. Japanese Feeding Standard for Beef Cattle 2008 Edn; Japan Livestock Industry Association: Tokyo, Japan, 2009. (In Japanese)
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Rozance, P.J.; Zastoupil, L.; Wesolowski, S.R.; Goldstrohm, D.A.; Strahan, B.; Cree-Green, M.; Sheffield-Moore, M.; Meschia, G.; Hay, W.W., Jr.; Wilkening, R.B.; et al. Skeletal muscle protein accretion rates and hindlimb growth are reduced in late gestation intrauterine growth-restricted fetal sheep. J. Physiol. 2018, 596, 67–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, M.; Zhu, M.J.; Means, W.J.; Hess, B.W.; Ford, S.P. Effect of nutrient restriction on calpain and calpastatin content of skeletal muscle from cows and fetuses1. J. Anim. Sci. 2004, 82, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T. Interorgan amino acid transport and its regulation. J. Nutr. 2003, 133, 2068s–2072s. [Google Scholar] [CrossRef] [Green Version]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- Morris, C.R.; Hamilton-Reeves, J.; Martindale, R.G.; Sarav, M.; Ochoa Gautier, J.B. Acquired Amino Acid Deficiencies: A Focus on Arginine and Glutamine. Nutr. Clin. Pract. 2017, 32, 30S–47S. [Google Scholar] [CrossRef]
- Yoo, H.C.; Yu, Y.C.; Sung, Y.; Han, J.M. Glutamine reliance in cell metabolism. Exp. Mol. Med. 2020, 52, 1496–1516. [Google Scholar] [CrossRef] [PubMed]
- Leury, B.J.; Bird, A.R.; Chandler, K.D.; Bell, A.W. Glucose partitioning in the pregnant ewe: Effects of undernutrition and exercise. Br. J. Nutr. 1990, 64, 449–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemons, J.A.; Schreiner, R.L. Amino acid metabolism in the ovine fetus. Am. J. Physiol. 1983, 244, E459–E466. [Google Scholar] [CrossRef] [PubMed]
- Faichney, G.J.; White, G.A. Effects of maternal nutritional status on fetal and placental growth and on fetal urea synthesis in sheep. Aust. J. Biol. Sci. 1987, 40, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, S.M., Jr. Arginine Metabolism Revisited. J. Nutr. 2016, 146, 2579S–2586S. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, M.; Leonetti, A.; Duranti, G.; Sabatini, S.; Ceci, R.; Mariottini, P. Skeletal Muscle Pathophysiology: The Emerging Role of Spermine Oxidase and Spermidine. Med. Sci. 2018, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosten, A.O. BUN and Creatinine. In Clinical Methods: The History, Physical, and Laboratory Examinations; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworths: Boston, MA, USA, 1990; pp. 874–878. [Google Scholar]
- Bessman, S.P.; Geiger, P.J. Transport of energy in muscle: The phosphorylcreatine shuttle. Science 1981, 211, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Dessì, A.; Atzori, L.; Noto, A.; Visser, G.H.; Gazzolo, D.; Zanardo, V.; Barberini, L.; Puddu, M.; Ottonello, G.; Atzei, A.; et al. Metabolomics in newborns with intrauterine growth retardation (IUGR): Urine reveals markers of metabolic syndrome. J. Matern. Fetal Neonatal Med. 2011, 24 (Suppl. 2), 35–39. [Google Scholar] [CrossRef]
- Zorzano, A.; Lasunción, M.A.; Herrera, E. Role of the availability of substrates on hepatic and renal gluconeogenesis in the fasted late pregnant rat. Metabolism 1986, 35, 297–303. [Google Scholar] [CrossRef]
- Zeng, Z.; Liu, F.; Li, S. Metabolic Adaptations in Pregnancy: A Review. Ann. Nutr. Metab. 2017, 70, 59–65. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Bizzarri, M. Inositols in Insulin Signaling and Glucose Metabolism. Int. J. Endocrinol. 2018, 2018, 1968450. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.C.; Fonteles, M.C.; Houston, D.B.; Zhang, C.; Larner, J. Chiroinositol deficiency and insulin resistance. III. Acute glycogenic and hypoglycemic effects of two inositol phosphoglycan insulin mediators in normal and streptozotocin-diabetic rats in vivo. Endocrinology 1993, 132, 652–657. [Google Scholar] [CrossRef]
- Larner, J. D-chiro-inositol-its functional role in insulin action and its deficit in insulin resistance. Int. J. Exp. Diabetes Res. 2002, 3, 47–60. [Google Scholar] [CrossRef]
- Regnault, T.R.; Teng, C.; de Vrijer, B.; Galan, H.L.; Wilkening, R.B.; Battaglia, F.C. The tissue and plasma concentration of polyols and sugars in sheep intrauterine growth retardation. Exp. Biol. Med. 2010, 235, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.M.; Nebel, C.; Oksbjerg, N.; Bertram, H.C. Metabolomics reveals relationship between plasma inositols and birth weight: Possible markers for fetal programming of type 2 diabetes. J. Biomed. Biotechnol. 2011, 2011, 378268. [Google Scholar] [CrossRef] [PubMed]
- Metges, C.C.; Görs, S.; Lang, I.S.; Hammon, H.M.; Brüssow, K.P.; Weitzel, J.M.; Nürnberg, G.; Rehfeldt, C.; Otten, W. Low and high dietary protein: Carbohydrate ratios during pregnancy affect materno-fetal glucose metabolism in pigs. J. Nutr. 2014, 144, 155–163. [Google Scholar] [CrossRef]
- Priante, E.; Verlato, G.; Giordano, G.; Stocchero, M.; Visentin, S.; Mardegan, V.; Baraldi, E. Intrauterine Growth Restriction: New Insight from the Metabolomic Approach. Metabolites 2019, 9, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, H.; Kakimoto, Y.; Nakajima, T.; Kanazawa, A.; Sano, I. Isolation and identification of 2-aminoethylphosphonic acid from bovine brain. Nature 1965, 207, 1197–1198. [Google Scholar] [CrossRef] [PubMed]
- Alhadeff, J.A.; Van Bruggen, J.T.; Doyle Daves, G. Biosynthetic studies on 2-aminoethylphosphonic acid in a mammalian (rat) system. Biochim. Biophys. Acta (BBA)—Gen. Subj. 1972, 286, 103–106. [Google Scholar] [CrossRef]
- Hasegawa, S.; Tamari, M.; Kametaka, M. Isolation of diacylglyceryl-2-aminoethylphosphonate from bovine liver. J. Biochem. 1976, 80, 531–535. [Google Scholar] [CrossRef]
- Tan, S.A.; Tan, L.G. Distribution of ciliatine (2-aminoethylphosphonic acid) and phosphonoalanine (2-amino-3-phosphonopropionic acid) in human tissues. Clin. Physiol. Biochem. 1989, 7, 303–309. [Google Scholar] [PubMed]
- Joseph, J.C.; Henderson, T.O. 2-Aminoethylphosphonic acid metabolism in the rat. Lipids 1977, 12, 75–84. [Google Scholar] [CrossRef]
- Nagata, Y.; Partridge, T.A.; Matsuda, R.; Zammit, P.S. Entry of muscle satellite cells into the cell cycle requires sphingolipid signaling. J. Cell Biol. 2006, 174, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Stamler, J.S.; Meissner, G. Physiology of nitric oxide in skeletal muscle. Physiol. Rev. 2001, 81, 209–237. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.L.; Cartland, D.; Hussain, A.; Egginton, S. A differential role for nitric oxide in two forms of physiological angiogenesis in mouse. J. Physiol. 2006, 570, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal nutrition and fetal development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef]
- Hellwig-Bürgel, T.; Stiehl, D.P.; Wagner, A.E.; Metzen, E.; Jelkmann, W. Review: Hypoxia-inducible factor-1 (HIF-1): A novel transcription factor in immune reactions. J. Interferon Cytokine Res. 2005, 25, 297–310. [Google Scholar] [CrossRef]
- Lira, V.A.; Soltow, Q.A.; Long, J.H.; Betters, J.L.; Sellman, J.E.; Criswell, D.S. Nitric oxide increases GLUT4 expression and regulates AMPK signaling in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1062–E1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef] [PubMed]
- Kjøbsted, R.; Hingst, J.R.; Fentz, J.; Foretz, M.; Sanz, M.-N.; Pehmøller, C.; Shum, M.; Marette, A.; Mounier, R.; Treebak, J.T.; et al. AMPK in skeletal muscle function and metabolism. FASEB J. 2018, 32, 1741–1777. [Google Scholar] [CrossRef] [Green Version]
- Bahadoran, Z.; Mirmiran, P.; Ghasemi, A. Role of Nitric Oxide in Insulin Secretion and Glucose Metabolism. Trends Endocrinol. Metab. TEM 2020, 31, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Satterfield, M.C.; Li, X.; Wang, X.; Johnson, G.A.; Burghardt, R.C.; Dai, Z.; Wang, J.; Wu, Z. Impacts of arginine nutrition on embryonic and fetal development in mammals. Amino Acids 2013, 45, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Hernando, C.; Suárez, Y. ANGPTL4: A multifunctional protein involved in metabolism and vascular homeostasis. Curr. Opin. Hematol. 2020, 27, 206–213. [Google Scholar] [CrossRef]
- Chong, H.C.; Chan, J.S.; Goh, C.Q.; Gounko, N.V.; Luo, B.; Wang, X.; Foo, S.; Wong, M.T.; Choong, C.; Kersten, S.; et al. Angiopoietin-like 4 stimulates STAT3-mediated iNOS expression and enhances angiogenesis to accelerate wound healing in diabetic mice. Mol. Ther. 2014, 22, 1593–1604. [Google Scholar] [CrossRef] [Green Version]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Aryal, B.; Price, N.L.; Suarez, Y.; Fernández-Hernando, C. ANGPTL4 in Metabolic and Cardiovascular Disease. Trends Mol. Med. 2019, 25, 723–734. [Google Scholar] [CrossRef]
- Muroya, S.; Ogasawara, H.; Nohara, K.; Oe, M.; Ojima, K.; Hojito, M. Coordinated alteration of mRNA-microRNA transcriptomes associated with exosomes and fatty acid metabolism in adipose tissue and skeletal muscle in grazing cattle. Asian-Australas. J. Anim. Sci. 2020, 33, 1824–1836. [Google Scholar] [CrossRef] [PubMed]
- La Paglia, L.; Listì, A.; Caruso, S.; Amodeo, V.; Passiglia, F.; Bazan, V.; Fanale, D. Potential Role of ANGPTL4 in the Cross Talk between Metabolism and Cancer through PPAR Signaling Pathway. PPAR Res. 2017, 2017, 8187235. [Google Scholar] [CrossRef]
- Joseph, S.; Alava, B.; Antolic, A.; Richards, E.M.; Wood, C.E.; Keller-Wood, M. Fetal ovine skeletal and cardiac muscle transcriptomics are differentially altered by increased maternal cortisol during gestation. Physiol. Genom. 2020, 52, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hulver, M.W.; McMillan, R.P.; Cline, M.A.; Gilbert, E.R. The pivotal role of pyruvate dehydrogenase kinases in metabolic flexibility. Nutr. Metab. 2014, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayeur, S.; Wattez, J.S.; Lukaszewski, M.A.; Lecoutre, S.; Butruille, L.; Drougard, A.; Eberlé, D.; Bastide, B.; Laborie, C.; Storme, L.; et al. Apelin Controls Fetal and Neonatal Glucose Homeostasis and Is Altered by Maternal Undernutrition. Diabetes 2016, 65, 554–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecoutre, S.; Marousez, L.; Drougard, A.; Knauf, C.; Guinez, C.; Eberlé, D.; Laborie, C.; Vieau, D.; Lesage, J.; Breton, C. Maternal undernutrition programs the apelinergic system of adipose tissue in adult male rat offspring. J. Dev. Orig. Health Dis. 2017, 8, 3–7. [Google Scholar] [CrossRef]
- Shin, K.; Kenward, C.; Rainey, J.K. Apelinergic System Structure and Function. Compr. Physiol. 2017, 8, 407–450. [Google Scholar] [CrossRef]
- Dray, C.; Knauf, C.; Daviaud, D.; Waget, A.; Boucher, J.; Buléon, M.; Cani, P.D.; Attané, C.; Guigné, C.; Carpéné, C.; et al. Apelin stimulates glucose utilization in normal and obese insulin-resistant mice. Cell Metab. 2008, 8, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.; Jin, H.; Aillaud, M.; Deng, A.C.; Azuma, J.; Asagami, T.; Kundu, R.K.; Reaven, G.M.; Quertermous, T.; Tsao, P.S. Apelin is necessary for the maintenance of insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E59–E67. [Google Scholar] [CrossRef] [Green Version]
- Pratt, J.; Boreham, C.; Ennis, S.; Ryan, A.W.; De Vito, G. Genetic Associations with Aging Muscle: A Systematic Review. Cells 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinitz, S.; Piaggi, P.; Yang, S.; Bonfiglio, S.; Steel, J.; Krakoff, J.; Votruba, S.B. Response of skeletal muscle UCP2-expression during metabolic adaptation to caloric restriction. Int. J. Obes. 2018, 42, 974–984. [Google Scholar] [CrossRef]
- Bonnefont, J.P.; Djouadi, F.; Prip-Buus, C.; Gobin, S.; Munnich, A.; Bastin, J. Carnitine palmitoyltransferases 1 and 2: Biochemical, molecular and medical aspects. Mol. Asp. Med. 2004, 25, 495–520. [Google Scholar] [CrossRef] [PubMed]
- Nowacka-Woszuk, J.; Pruszynska-Oszmalek, E.; Szydlowski, M.; Sadkowski, S.; Szczerbal, I. Diet-induced variability of the resistin gene (Retn) transcript level and methylation profile in rats. BMC Genet. 2015, 16, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grempler, R.; Wolff, M.; Simon, E.; Schmid, R.; Eisele, C.; Rieber, K.; Fischer, E.; Mettel, S.; Gabrielyan, O.; Delic, D.; et al. Discovery and translation of a target engagement marker for AMP-activated protein kinase (AMPK). PLoS ONE 2018, 13, e0197849. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, H.; Zhang, B.; Ling, Y.; Zhang, H. Identification of key HIF-1α target genes that regulate adaptation to hypoxic conditions in Tibetan chicken embryos. Gene 2020, 729, 144321. [Google Scholar] [CrossRef]
- Gong, X.M.; Li, Y.F.; Luo, J.; Wang, J.Q.; Wei, J.; Wang, J.Q.; Xiao, T.; Xie, C.; Hong, J.; Ning, G.; et al. Gpnmb secreted from liver promotes lipogenesis in white adipose tissue and aggravates obesity and insulin resistance. Nat. Metab. 2019, 1, 570–583. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-Carbon Metabolism in Health and Disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Scalise, M.; Galluccio, M.; Console, L.; Pochini, L.; Indiveri, C. The Human SLC7A5 (LAT1): The Intriguing Histidine/Large Neutral Amino Acid Transporter and Its Relevance to Human Health. Front. Chem. 2018, 6, 243. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, R.; Tomi, M.; Akanuma, S.-i.; Tabuchi, A.; Kubo, Y.; Tachikawa, M.; Hosoya, K.-i. Up-regulation of l-Type Amino Acid Transporter 1 (LAT1) in Cultured Rat Retinal Capillary Endothelial Cells in Response to Glucose Deprivation. Drug Metab. Pharmacokinet. 2012, 27, 317–324. [Google Scholar] [CrossRef]
- Walker, D.K.; Drummond, M.J.; Dickinson, J.M.; Borack, M.S.; Jennings, K.; Volpi, E.; Rasmussen, B.B. Insulin increases mRNA abundance of the amino acid transporter SLC7A5/LAT1 via an mTORC1-dependent mechanism in skeletal muscle cells. Physiol. Rep. 2014, 2, e00238. [Google Scholar] [CrossRef]
- Taylor, P.M. Role of amino acid transporters in amino acid sensing. Am. J. Clin. Nutr. 2014, 99, 223s–230s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, E.; Fukushima, N.; Kanetani, T.; Nishimura, S. The Optimum Method for Additional Daily Nutrient Requirements for Pregnant Japanese Black Cattle in Later Pregnancy. Bull. Okayama Prefect. Cent. Anim. Husb. Res. 2019, 9, 11–16. (In Japanese) [Google Scholar]
- Institute of Laboratory Animal Science Research Support Center Kagoshima University. The Guide for the Care and Use of Experimental Animals. (In Japanese). Available online: https://www.kufm.kagoshima-u.ac.jp/~animal/tebiki.htm (accessed on 9 August 2021).
- Muroya, S.; Oe, M.; Nakajima, I.; Ojima, K.; Chikuni, K. CE-TOF MS-based metabolomic profiling revealed characteristic metabolic pathways in postmortem porcine fast and slow type muscles. Meat Sci. 2014, 98, 726–735. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HN (n = 5) | LN (n = 6) | ||||
|---|---|---|---|---|---|
| Mean | SE | Mean * | SE * | p-Value | |
| Age (d) | 261.6 | 1.5 | 260.7 | 1.6 | 0.684 |
| BW (kg) | 32.5 | 0.5 | 23.4 | 2.4 | 0.005 |
| Total muscle (g) | 4889.4 | 128.7 | 3496.8 | 401.3 | 0.009 |
| % of total BW | 15.0 | 0.4 | 14.9 | 0.4 | 0.733 |
| LT muscle (g) | 272.0 | 7.4 | 187.0 | 24.1 | 0.008 |
| % of total muscle | 5.6 | 0.1 | 5.3 | 0.2 | 0.183 |
| Compound | LN | HN | Ratio (LN/HN) | p-Value | ||
|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | |||
| Carnosine | 0.0197 | 0.0011 | 0.0148 | 0.0007 | 1.3 | 0.006 |
| Glutamine | 0.0215 | 0.0009 | 0.0168 | 0.0008 | 1.3 | 0.008 |
| Glycerol | 0.1367 | 0.0105 | 0.1038 | 0.0072 | 1.3 | 0.022 |
| Creatine | 0.0550 | 0.0026 | 0.0447 | 0.0022 | 1.2 | 0.023 |
| N6-Methyllysine | 0.0037 | 0.0003 | 0.0026 | 0.0002 | 1.5 | 0.028 |
| Phosphorylcholine | 0.0125 | 0.0011 | 0.0089 | 0.0007 | 1.4 | 0.034 |
| Phenylalanine | 0.0097 | 0.0009 | 0.0066 | 0.0007 | 1.5 | 0.039 |
| myo-Inositol 2-phosphate | 0.0007 | 0.0000 | 0.0009 | 0.0001 | 0.8 | 0.039 |
| 2-Aminoethylphosphonic acid | 0.0002 | 0.0000 | 0.0003 | 0.0000 | 0.6 | 0.041 |
| Proline | 0.0386 | 0.0033 | 0.0292 | 0.0016 | 1.3 | 0.043 |
| Alanine | 0.1565 | 0.0044 | 0.1276 | 0.0110 | 1.2 | 0.059 |
| Putrescine | 0.0022 | 0.0002 | 0.0015 | 0.0002 | 1.4 | 0.065 |
| Creatinine | 0.0189 | 0.0011 | 0.0151 | 0.0016 | 1.2 | 0.070 |
| Gamma-amino butyric acid | 0.0071 | 0.0005 | 0.0055 | 0.0005 | 1.3 | 0.077 |
| Histidine | 0.0034 | 0.0006 | 0.0019 | 0.0003 | 1.8 | 0.081 |
| Glycine | 0.0701 | 0.0065 | 0.0506 | 0.0073 | 1.4 | 0.090 |
| 2-Hydroxyvaleric acid | 0.0002 | 0.0000 | 0.0002 | 0.0001 | 0.6 | 0.098 |
| Taurine | 0.0249 | 0.0019 | 0.0307 | 0.0026 | 0.8 | 0.111 |
| S-Adenosylmethionine | 0.0006 | 0.0001 | 0.0008 | 0.0001 | 0.7 | 0.113 |
| Dihydroxyacetone phosphate | 0.0082 | 0.0016 | 0.0047 | 0.0011 | 1.7 | 0.118 |
| Metabolism/Pathway | Hits/Total Metabolites | p-Value | Increased in LN | Decreased in LN |
|---|---|---|---|---|
| Pyrimidine metabolism | 2/39 | 0.003 | Glutamine, Deoxyuridine | |
| Aminoacyl-tRNA biosynthesis | 13/48 | 0.007 | Glutamine, Phenylalanine, Proline, Alanine, Valine, Histidine, Glycine, Tryptophan, Serine, Tyrosine, Glutamate, Arginine | Methionine |
| Glycerolipid metabolism | 2/16 | 0.007 | Glycerol, DHAP | |
| Arginine biosynthesis | 4/14 | 0.007 | Glutamine, Ornithine, Glutamate, Arginine | |
| Alanine, aspartate and glutamate metabolism | 5/28 | 0.010 | Glutamine, Alanine, GABA, Pyruvate, Glutamate | |
| Glutathione metabolism | 6/28 | 0.011 | Putrescine, Glycine, Ornithine, Spermidine, Spermine, Glutamate | |
| Histidine metabolism | 3/16 | 0.012 | Carnosine, Histidine, Glutamate | |
| Arginine and proline metabolism | 11/38 | 0.012 | Creatine, Proline, Putrescine, GABA, Ornithine, Spermidine, Pyruvate, Spermine, Glutamate, Arginine | SAM |
| Glyoxylate and dicarboxylate metabolism | 5/32 | 0.013 | Glutamine, Glycine, Serine, Pyruvate, Glutamate | |
| Phosphonate and phosphinate metabolism | 2/6 | 0.017 | Phosphorylcholine | 2-AEP |
| Glycerophospholipid metabolism | 2/36 | 0.018 | Phosphorylcholine, DHAP | |
| Galactose metabolism | 1/27 | 0.022 | Glycerol | |
| Primary bile acid biosynthesis | 2/46 | 0.025 | Glycine | Taurine |
| Purine metabolism | 2/65 | 0.025 | Glutamine, Allantoin | |
| D-Glutamine and D-glutamate metabolism | 4/6 | 0.029 | Glutamate, Glutamine | |
| Nitrogen metabolism | 2/6 | 0.029 | Glutamate, Glutamine | |
| Glycine, serine and threonine metabolism | 5/33 | 0.036 | Creatine, Glycine, Serine, Pyruvate | Cystathionine |
| β-Alanine metabolism | 4/21 | 0.040 | Carnosine, Histidine, Spermidine, Spermine | |
| Glycolysis/Gluconeogenesis | 3/26 | 0.058 | Pyruvate, PEP, DHAP | |
| Selenocompound metabolism | 1/20 | 0.059 | Alanine |
| Category | Term | p-Value | Fold Enrichment | Validated Genes |
|---|---|---|---|---|
| KEGG Pathway | ||||
| bta04940:Type I diabetes mellitus | <0.001 | 3.8050 | ||
| bta00010:Glycolysis/Gluconeogenesis | <0.001 | 3.1407 | GPI, ENO3, PFKM, PGAM1, FBP2 | |
| bta04010:MAPK signaling pathway | <0.001 | 1.9194 | MAPK3, NFKB1, RELA | |
| bta04066:HIF-1 signaling pathway | <0.001 | 2.4950 | ANGPT4, NOS2, NOS3, ENO3, MAPK3, NFKB1, PLCG1, RELA | |
| bta01200:Carbon metabolism | <0.001 | 2.1974 | GPI, PGAM1, ENO3, PFKM, FBP2 | |
| bta04151:PI3K-Akt signaling pathway | <0.001 | 1.5906 | ANGPT4, NOS3, MAPK3, NFKB1, PRKAA1, RELA, THBS1 | |
| bta04919:Thyroid hormone signaling pathway | <0.001 | 2.1196 | MAPK3, PLCG1 | |
| bta00030:Pentose phosphate pathway | 0.003 | 3.4713 | GPI, PFKM, FBP2 | |
| bta04910:Insulin signaling pathway | 0.005 | 1.8377 | FBP2, MAPK3, PRKAA1 | |
| bta04115:p53 signaling pathway | 0.007 | 2.2001 | THBS1 | |
| GO: Biological Process | ||||
| GO:0006096~glycolytic process | <0.001 | 4.3200 | GPI, ENO3, PGAM1 | |
| GO:0034097~response to cytokine | <0.001 | 4.0909 | NFKB1, RELA | |
| GO:0006094~gluconeogenesis | <0.001 | 4.2078 | GPI, FBP2, PGAM1 | |
| GO:0000122~negative regulation of transcription from RNA polymerase II promoter | 0.002 | 1.4727 | FOXP1, NFKB1, RELA | |
| GO:0001525~angiogenesis | 0.002 | 1.9050 | GPI, NOS3, ANGPT4, ANGPTL4 | |
| GO:0042981~regulation of apoptotic process | 0.002 | 1.9474 | PINK1, RELA | |
| GO:0050995~negative regulation of lipid catabolic process | 0.009 | 5.4545 | PRKAA1 | |
| GO:0090200~positive regulation of release of cytochrome c from mitochondria | 0.009 | 3.6172 | PINK1 | |
| GO:0043536~positive regulation of blood vessel endothelial cell migration | 0.010 | 4.2078 | ANGPTL4, PLCG1, THBS1 | |
| GO:0042177~negative regulation of protein catabolic process | 0.010 | 3.1418 | NOS2, SIRT2, RELA | |
| Category | Term | p-Value | Fold Enrichment | Validated Genes |
|---|---|---|---|---|
| KEGG Pathway | ||||
| bta01230:Biosynthesis of amino acids | <0.001 | 2.8335 | ||
| bta00970:Aminoacyl-tRNA biosynthesis | <0.001 | 3.4187 | ||
| bta00260:Glycine, serine and threonine metabolism | <0.001 | 3.4636 | ||
| bta00072:Synthesis and degradation of ketone bodies | 0.001 | 6.4548 | BDH1 | |
| bta04512:ECM-receptor interaction | 0.002 | 2.3124 | ||
| bta00670:One carbon pool by folate | 0.003 | 4.6021 | MTHFD2, ALDH1L2 | |
| bta04550:Signaling pathways regulating pluripotency of stem cells | 0.003 | 1.9581 | ||
| bta04974:Protein digestion and absorption | 0.004 | 2.2541 | ||
| GO: Biological Process | ||||
| GO:0006730~one-carbon metabolic process | <0.001 | 4.4172 | MTHFD2, ALDH1L2 | |
| GO:0046653~tetrahydrofolate metabolic process | <0.001 | 10.6749 | MTHFD2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muroya, S.; Zhang, Y.; Kinoshita, A.; Otomaru, K.; Oshima, K.; Gotoh, Y.; Oshima, I.; Sano, M.; Roh, S.; Oe, M.; et al. Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle. Metabolites 2021, 11, 582. https://doi.org/10.3390/metabo11090582
Muroya S, Zhang Y, Kinoshita A, Otomaru K, Oshima K, Gotoh Y, Oshima I, Sano M, Roh S, Oe M, et al. Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle. Metabolites. 2021; 11(9):582. https://doi.org/10.3390/metabo11090582
Chicago/Turabian StyleMuroya, Susumu, Yi Zhang, Aoi Kinoshita, Kounosuke Otomaru, Kazunaga Oshima, Yuji Gotoh, Ichiro Oshima, Mitsue Sano, Sanggun Roh, Mika Oe, and et al. 2021. "Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle" Metabolites 11, no. 9: 582. https://doi.org/10.3390/metabo11090582
APA StyleMuroya, S., Zhang, Y., Kinoshita, A., Otomaru, K., Oshima, K., Gotoh, Y., Oshima, I., Sano, M., Roh, S., Oe, M., Ojima, K., & Gotoh, T. (2021). Maternal Undernutrition during Pregnancy Alters Amino Acid Metabolism and Gene Expression Associated with Energy Metabolism and Angiogenesis in Fetal Calf Muscle. Metabolites, 11(9), 582. https://doi.org/10.3390/metabo11090582