
Dysregulated Alanine as a Potential Predictive Marker of Glioma—An Insight from Untargeted HRMAS-NMR and Machine Learning Data

Abstract
:1. Introduction
2. Results
2.1. Clinicopathological Characteristics of Study Group
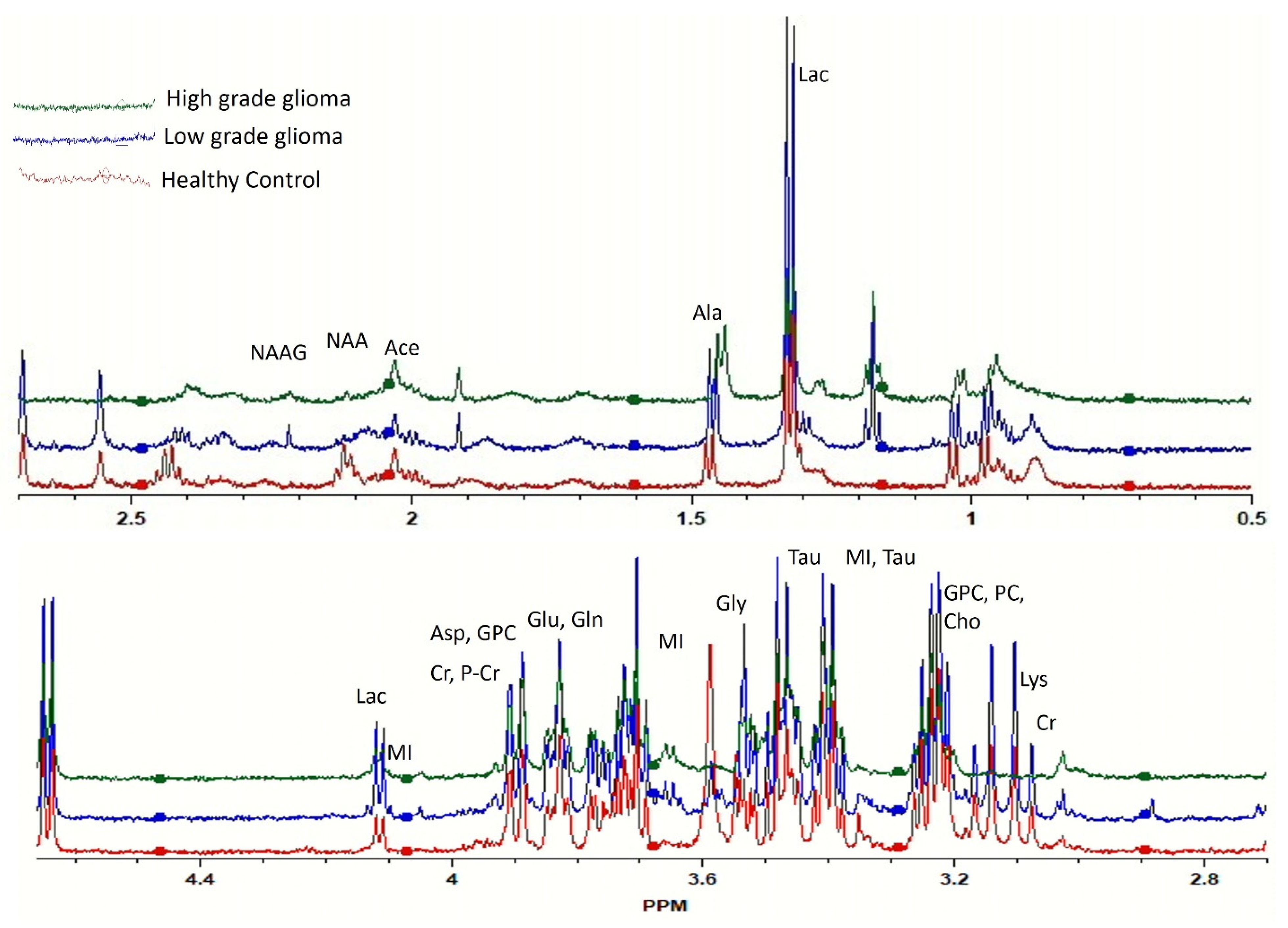
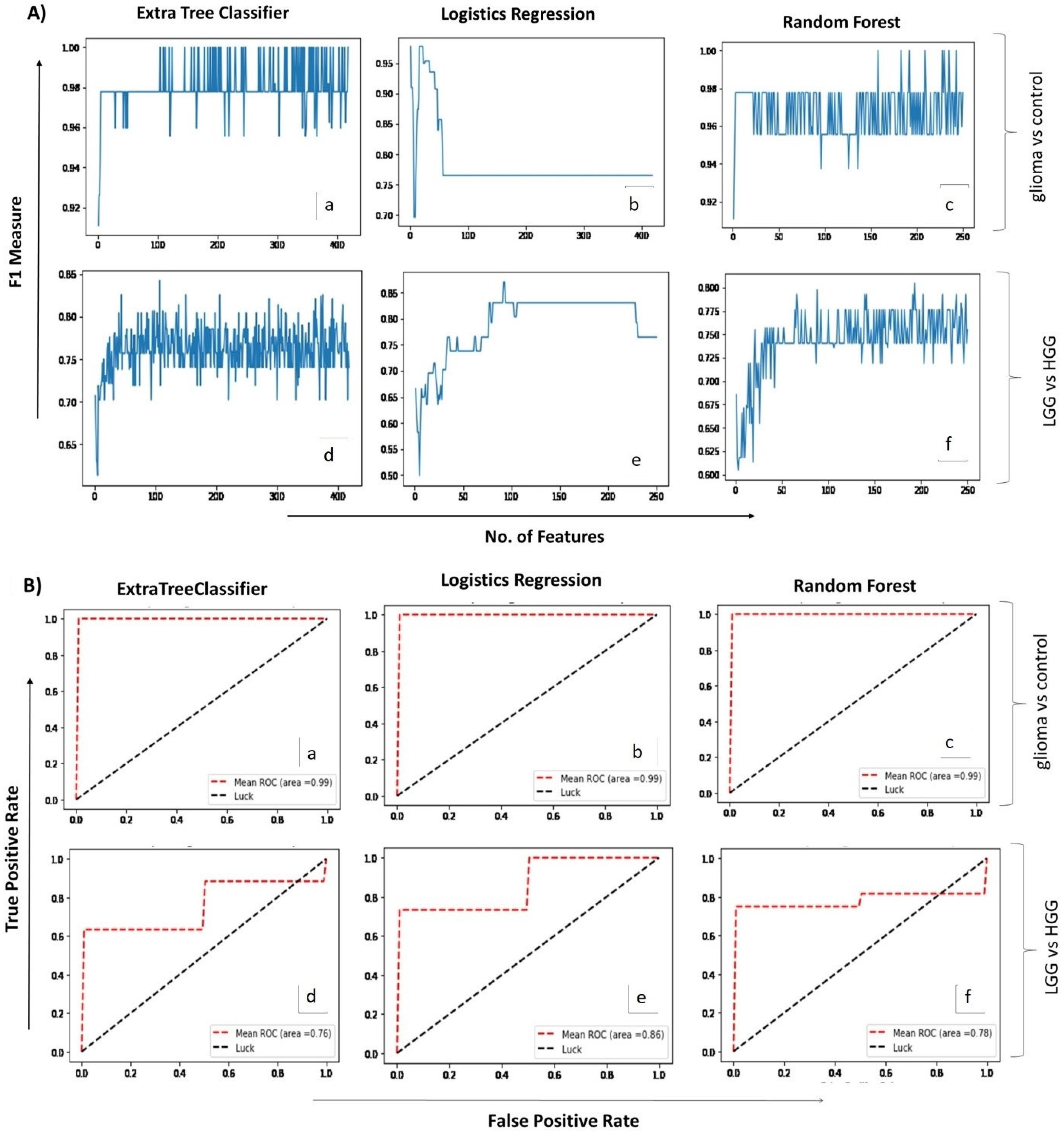
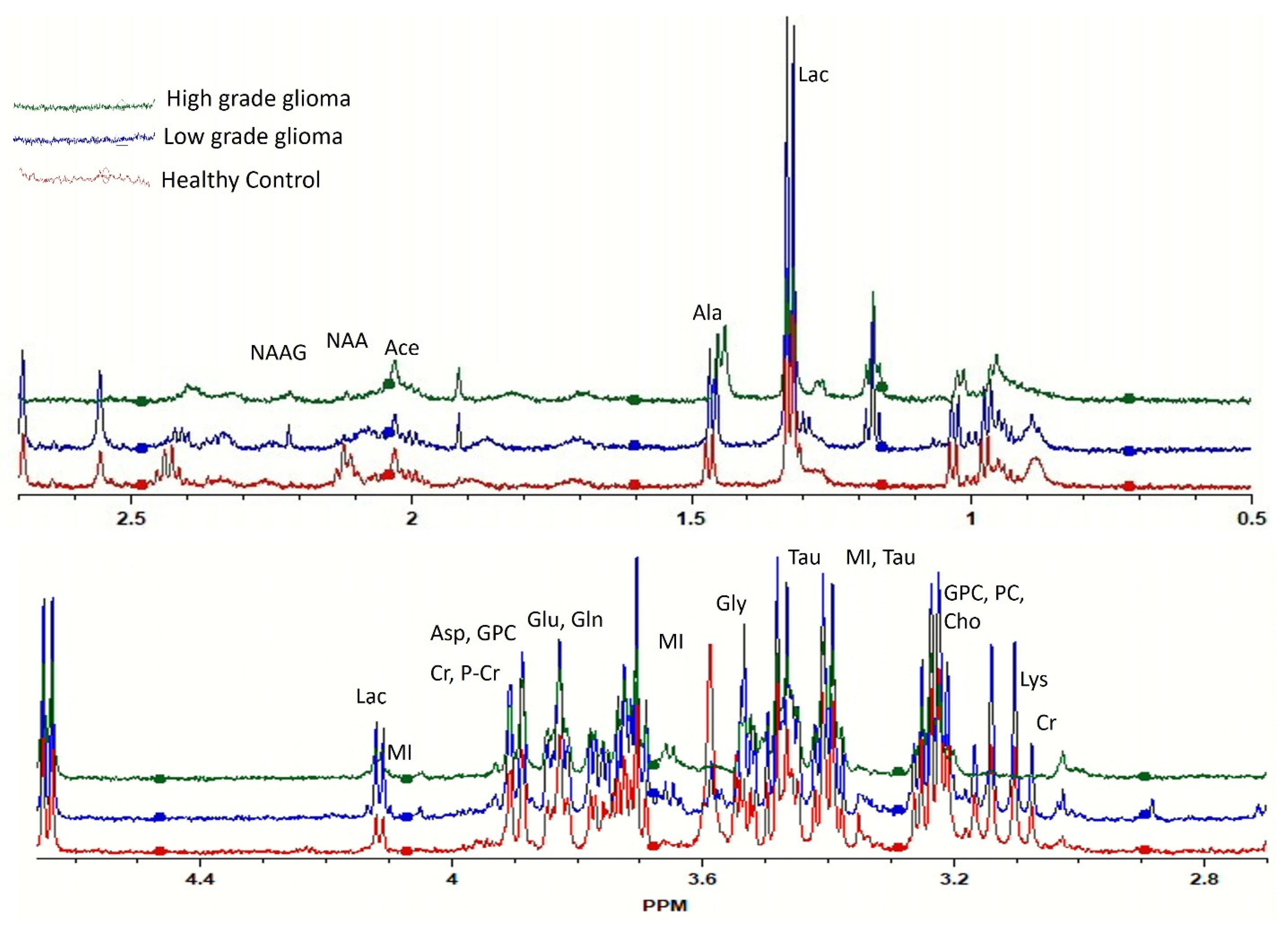
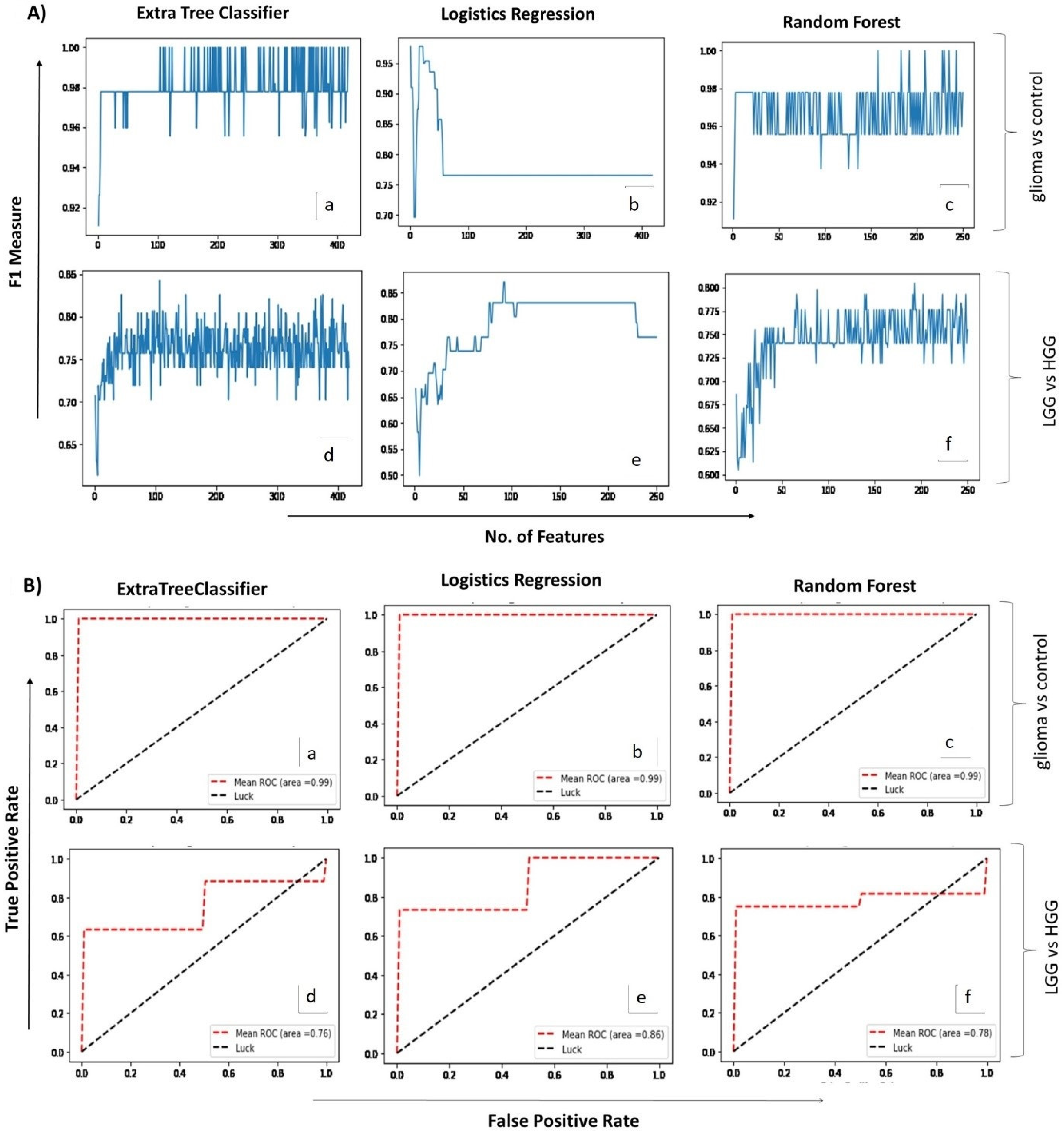
2.2. Correlating Circulating Metabolomic Profiles for Glioma Detection
2.3. Correlating Circulating Metabolomic Profiles for Glioma Classification (LGG/HGG)
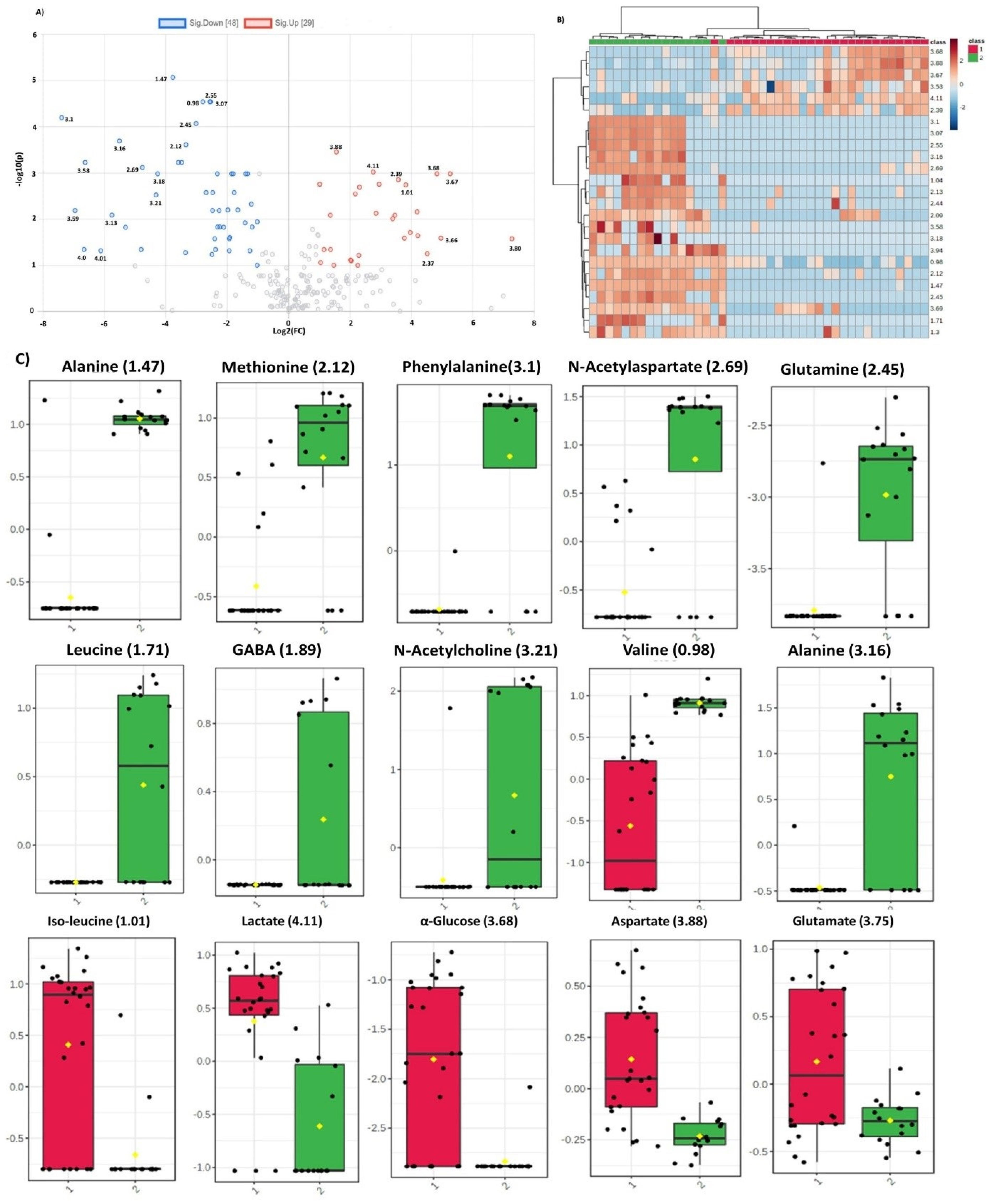
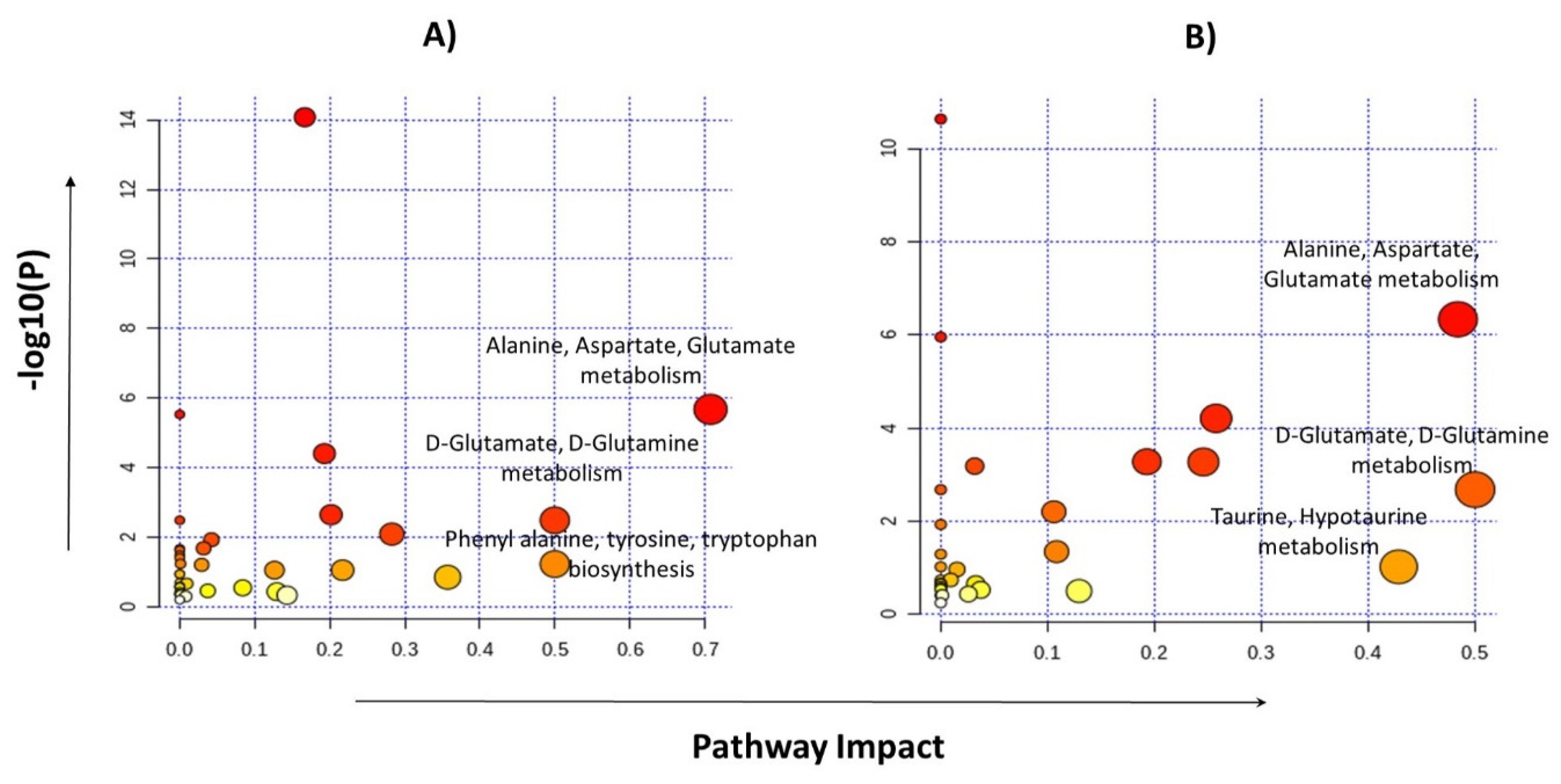
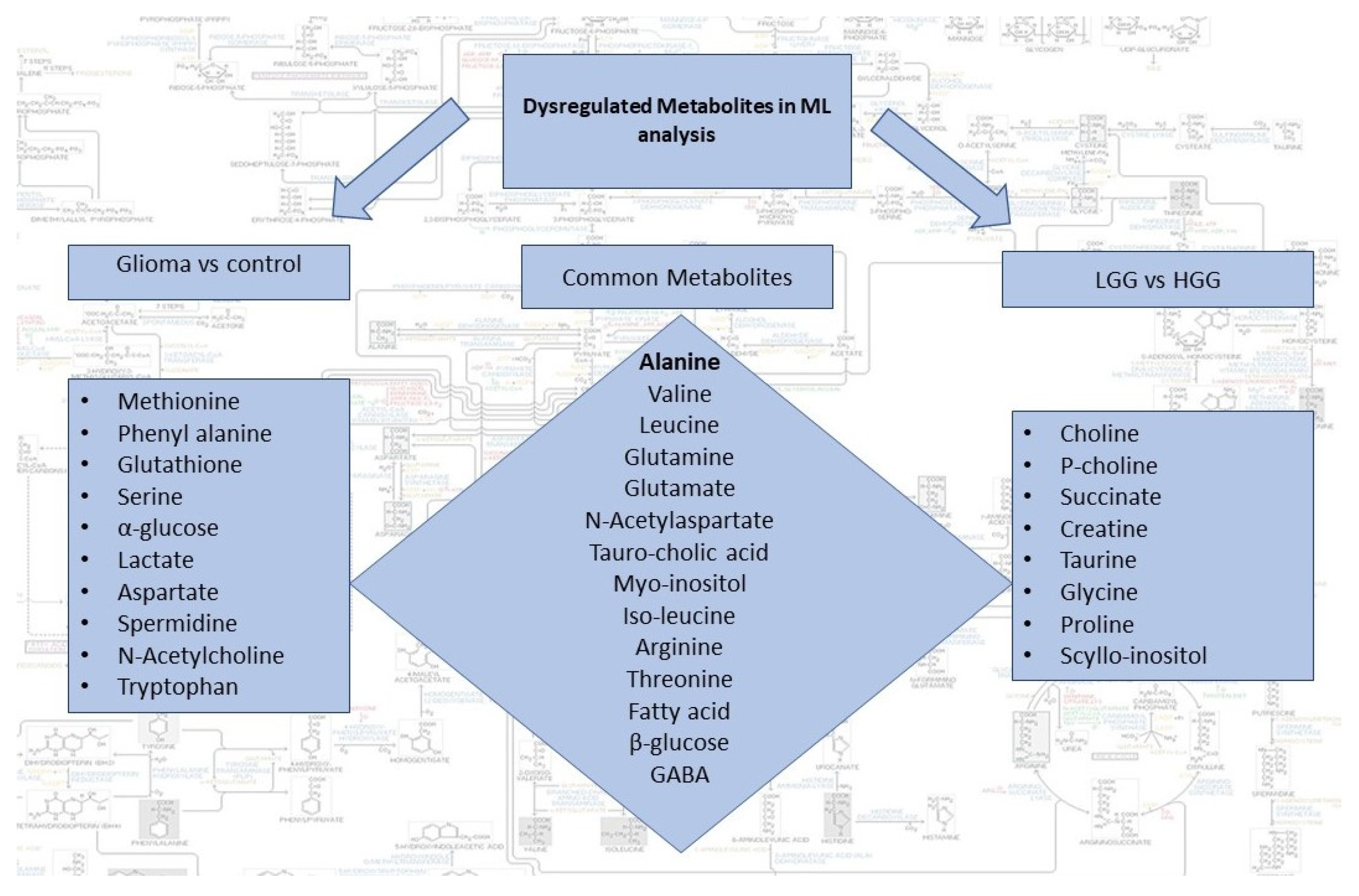
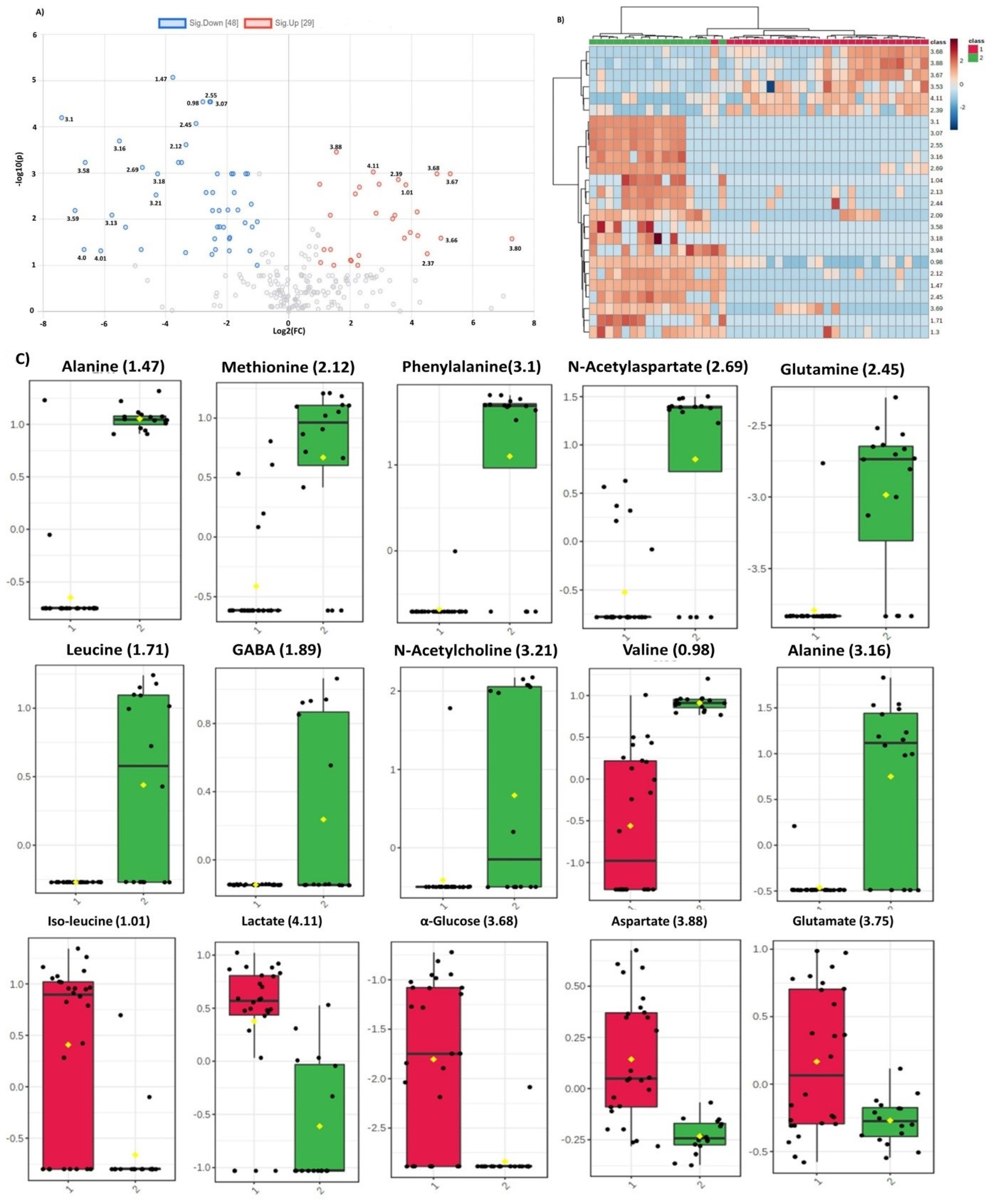
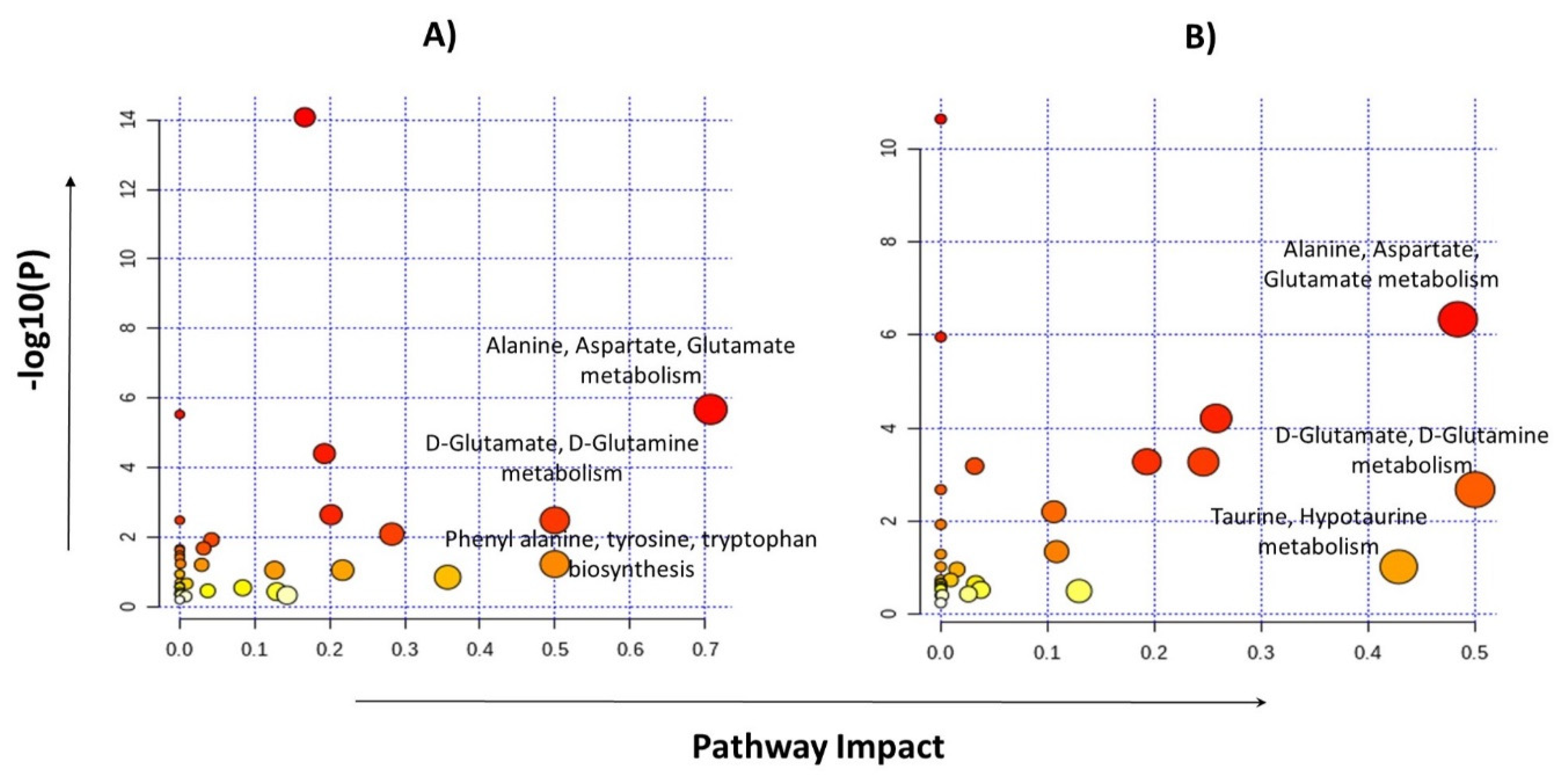
2.4. Identification of Statistically Significant Dysregulated Metabolites and Glioma-Associated Pathways
3. Discussion
4. Materials and Methods
4.1. Study Population and Sample Collection
4.2. Ex Vivo HRMAS-NMR
4.3. Pre-Processing of the Spectral Data
4.4. ML-Assisted Data Analysis
Recall = True positive/True positive + False negative
F1-measure= 2 × Precision × Recall/Precision + Recall
4.5. Statistical Analysis
4.6. Metabolite’s Identification and Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohgaki, H.; Kleihues, P. Epidemiology and etiology of gliomas. Acta Neuropathol. 2005, 109, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, B.K.; Hansen, S.; Laursen, R.J.; Kosteljanetz, M.; Schultz, H.; Nørgård, B.M.; Guldberg, R.; Gradel, K.O. Epidemiology of glioma: Clinical characteristics, symptoms, and predictors of glioma patients grade I–IV in the the Danish Neuro-Oncology Registry. J. Neurooncol. 2017, 135, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Diwanji, T.P.; Engelman, A.; Snider, J.W.; Mohindra, P. Epidemiology, diagnosis, and optimal management of glioma in adolescents and young adults. Adolesc. Health. Med. Ther. 2017, 8, 99. [Google Scholar] [CrossRef] [Green Version]
- Davis, M. GBM treatment overview. Clin. J. Oncol. Nurs. 2016, 20, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, C.; Costa, A.; Osório, L.; Lago, R.C.; Linhares, P.; Carvalho, B.; Caeiro, C. Current standards of care in glioblastoma therapy. Exon Publ. 2017, 197–241. [Google Scholar]
- Stupp, R.; Taillibert, S.; Kanner, A.A.; Kesari, S.; Steinberg, D.M.; Toms, S.A.; Taylor, L.P.; Lieberman, F.; Silvani, A.; Fink, K.L. Maintenance therapy with tumor-treating fields plus temozolomide vs temozolomide alone for glioblastoma: A randomized clinical trial. JAMA 2015, 314, 2535–2543. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zuo, W.; Yang, P.; Zhang, Y. Anti-PD-1, anti-VEGF, and temozolomide therapy in a patient with recurrent glioblastoma: A case report. J. Int. Med. Res. 2020, 48. [Google Scholar] [CrossRef]
- Wang, L.; Yekula, A.; Muralidharan, K.; Small, J.L.; Rosh, Z.S.; Kang, K.M.; Carter, B.S.; Balaj, L. Novel Gene Fusions in Glioblastoma Tumor Tissue and Matched Patient Plasma. Cancers 2020, 12, 1219. [Google Scholar] [CrossRef]
- Bi, J.; Chowdhry, S.; Wu, S.; Zhang, W.; Masui, K.; Mischel, P.S. Altered cellular metabolism in gliomas—an emerging landscape of actionable co-dependency targets. Nat. Rev. Cancer 2020, 20, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Lyssiotis, C.A.; Nagrath, D. Metabolic reprogramming and vulnerabilities in cancer. Cancer 2020, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Ward, P.S.; Thompson, C.B. Metabolic reprogramming: A cancer hallmark even warburg did not anticipate. Cancer Cell 2012, 21, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.Q.; Russo, J. Dysregulation of glucose transport, glycolysis, TCA cycle and glutaminolysis by oncogenes and tumor suppressors in cancer cells. Biochim. Biophys. Acta 2012, 1826, 370–384. [Google Scholar] [CrossRef] [PubMed]
- McCann, C.; Kerr, E.M. Metabolic reprogramming: A friend or foe to cancer therapy? Cancers (Basel) 2021, 13, 3351. [Google Scholar] [CrossRef] [PubMed]
- Venneti, S.; Thompson, C.B. Metabolic Reprogramming in Brain Tumors. Annu. Rev. Pathol. 2017, 12, 515–545. [Google Scholar] [CrossRef] [PubMed]
- Guda, M.R.; Labak, C.M.; Omar, S.I.; Asuthkar, S.; Airala, S.; Tuszynski, J.; Tsung, A.J.; Velpula, K.K. GLUT1 and TUBB4 in glioblastoma could be efficacious targets. Cancers 2019, 11, 1308. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Ma, M.; Becerra, L.; Ptak, T.; Tracey, I.; Lackner, A.; Gonzalez, R. Quantitative neuropathology by high resolution magic angle spinning proton magnetic resonance spectroscopy. Proc. Natl. Acad. Sci. USA 1997, 94, 6408–6413. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.L.; Chang, I.-W.; Louis, D.N.; Gonzalez, R.G. Correlation of high-resolution magic angle spinning proton magnetic resonance spectroscopy with histopathology of intact human brain tumor specimens. Cancer Res. 1998, 58, 1825–1832. [Google Scholar]
- Dietz, C.; Ehret, F.; Palmas, F.; Vandergrift, L.A.; Jiang, Y.; Schmitt, V.; Dufner, V.; Habbel, P.; Nowak, J.; Cheng, L.L. Applications of high-resolution magic angle spinning MRS in biomedical studies II-Human diseases. NMR Biomed. 2017, 30. [Google Scholar] [CrossRef]
- Cheng, L.L.; Anthony, D.C.; Comite, A.R.; Black, P.M.; Tzika, A.A.; Gonzalez, R.G. Quantification of microheterogeneity in glioblastoma multiforme with ex vivo high-resolution magic-angle spinning (HRMAS) proton magnetic resonance spectroscopy. Neuro-oncology 2000, 2, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Purohit, V.; Simeone, D.M.; Lyssiotis, C.A. Metabolic regulation of redox balance in cancer. Cancers 2019, 11, 955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, S.K.; Venneti, S. Glutamine metabolism in brain tumors. Cancers 2019, 11, 1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Heimberger, A.B.; Lu, Z.; Wu, X.; Hodges, T.R.; Song, R.; Shen, J. Metabolomics profiling in plasma samples from glioma patients correlates with tumor phenotypes. Oncotarget 2016, 7, 20486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, K.A.; Chinnaiyan, P. Applying metabolomics to understand the aggressive phenotype and identify novel therapeutic targets in glioblastoma. Metabolites 2014, 4, 740–750. [Google Scholar] [CrossRef] [Green Version]
- Jothi, J.; Vanisree, A.J.; Krishnaswamy, R. Metabolic variations among low grade and high grade gliomas–profiling by 1h nmr spectroscopy. J. Proteome Res. 2020. [Google Scholar] [CrossRef]
- Shen, J.; Song, R.; Hodges, T.R.; Heimberger, A.B.; Zhao, H. Identification of metabolites in plasma for predicting survival in glioblastoma. Mol. Carcinog. 2018, 57, 1078–1084. [Google Scholar] [CrossRef]
- Feng, S.; Liu, Y. Metabolomics of glioma. Adv. Exp. Med. Bio. 2021, 1280, 261–276. [Google Scholar] [CrossRef]
- Heinemann, J. Microbial Metabolomics; Springer: Berlin/Heidelberg, Germany, 2019; pp. 287–299. [Google Scholar] [CrossRef]
- Alakwaa, F.M.; Chaudhary, K.; Garmire, L.X. Deep learning accurately predicts estrogen receptor status in breast cancer metabolomics data. J. Proteome Res. 2018, 17, 337–347. [Google Scholar] [CrossRef]
- Ijare, O.; Baskin, D.; Pichumani, K. Cbmt-01. Alanine Fuels Energy Metabolism of Glioblastoma Cells. Neuro Oncol. 2019, 21 (Suppl. 6), vi32. [Google Scholar] [CrossRef]
- Waitkus, M.S.; Diplas, B.H.; Yan, H. Biological role and therapeutic potential of IDH mutations in cancer. Cancer Cell 2018, 34, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Srivastava, S.; Zhang, J. Starve cancer cells of glutamine: Break the spell or make a hungry monster? Cancers 2019, 11, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Márquez, J.; Alonso, F.J.; Matés, J.M.; Segura, J.A.; Martín-Rufián, M.; Campos-Sandoval, J.A. Glutamine addiction in gliomas. Neurochem. Res. 2017, 42, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Mancuso, A.; Daikhin, E.; Nissim, I.; Yudkoff, M.; Wehrli, S.; Thompson, C.B. Beyond aerobic glycolysis: Transformed cells can engage in glutamine metabolism that exceeds the requirement for protein and nucleotide synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 19345–19350. [Google Scholar] [CrossRef] [Green Version]
- Wise, D.R.; Thompson, C.B. Glutamine addiction: A new therapeutic target in cancer. Trends Biochem. Sci. 2010, 35, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, S.M.; Sontheimer, H. Glutamate transporters in the biology of malignant gliomas. Cell. Mol. Life. Sci. 2014, 71, 1839–1854. [Google Scholar] [CrossRef] [Green Version]
- Daye, D.; Wellen, K.E. Metabolic Reprogramming in Cancer: Unraveling the Role of Glutamine in Tumorigenesis; Elsevier: Amsterdam, The Netherlands, 2012; pp. 362–369. [Google Scholar] [CrossRef]
- Kallenberg, K.; Bock, H.C.; Helms, G.; Jung, K.; Wrede, A.; Buhk, J.-H.; Giese, A.; Frahm, J.; Strik, H.; Dechent, P. Untreated glioblastoma multiforme: Increased myo-inositol and glutamine levels in the contralateral cerebral hemisphere at proton MR spectroscopy. Radiology 2009, 253, 805–812. [Google Scholar] [CrossRef]
- Zhang, B.; Chen, Y.; Shi, X.; Zhou, M.; Bao, L.; Hatanpaa, K.J.; Patel, T.; DeBerardinis, R.J.; Wang, Y.; Luo, W. Regulation of branched-chain amino acid metabolism by hypoxia-inducible factor in glioblastoma. Cell. Mol. Life. Sci. 2020, 1–12. [Google Scholar] [CrossRef]
- Ananieva, E.A.; Wilkinson, A.C. Branched-chain amino acid metabolism in cancer. Curr. Opin. Clin. Nutr. Metab. Care. 2018, 21, 64. [Google Scholar] [CrossRef] [Green Version]
- Palanichamy, K.; Thirumoorthy, K.; Kanji, S.; Gordon, N.; Singh, R.; Jacob, J.R.; Sebastian, N.; Litzenberg, K.T.; Patel, D.; Bassett, E. Methionine and kynurenine activate oncogenic kinases in glioblastoma, and methionine deprivation compromises proliferation. Clin. Cancer Res. 2016, 22, 3513–3523. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.M.; Sheaff, M.T.; Szlosarek, P.W. Targeting arginine-dependent cancers with arginine-degrading enzymes: Opportunities and challenges. Cancer Res. Treat. 2013, 45, 251. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.J.; Fellows, G.A.; Griffiths, J.R.; Wilson, M.; Bell, B.A.; Howe, F.A. Ex-vivo HRMAS of adult brain tumours: Metabolite quantification and assignment of tumour biomarkers. Mol. Cancer 2010, 9, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, R.M.; Stern, P.H.; Coalson, D.W.; Wallace, C.D.; Erbe, R.W. Methionine Dependence of Cancer and Aging; Springer: Berlin/Heidelberg, Germany, 2019; pp. 13–26. [Google Scholar] [CrossRef]
- Tilgner, M.; Vater, T.S.; Habbel, P.; Cheng, L.L. High-Resolution Magic Angle Spinning (HRMAS) NMR Methods in Metabolomics. Methods Mol. Biol. 2019, 2037, 49–67. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | LGG (grade I–II) | HGG (grade III–IV) | Healthy Control | Total |
|---|---|---|---|---|
| No. of Subjects (n) | 4 + 5 = 9 | 1 + 16 = 17 | 16 | 42 |
| Mean Age (Years) | 33 ± 17 | 43 ± 16 | 34 ± 13 | 38 ± 16 |
| Gender | ||||
| Male | 07 | 11 | 07 | 25 |
| Female | 02 | 06 | 09 | 17 |
| Headache | ||||
| Yes | 05 | 13 | 0 | 18 |
| No | 04 | 04 | 16 | 24 |
| Epileptic Seizures | ||||
| Yes | 03 | 07 | 0 | 10 |
| No | 06 | 10 | 16 | 32 |
| Gait/Balance Changes | ||||
| Yes | 05 | 13 | 0 | 18 |
| No | 04 | 04 | 16 | 24 |
| Neurologic Deficits | ||||
| Yes | 07 | 15 | 0 | 22 |
| No | 02 | 02 | 16 | 20 |
| Reduced Vision | ||||
| Yes | 03 | 08 | 0 | 11 |
| No | 06 | 09 | 16 | 31 |
| Cancer History | ||||
| Yes | 01 | 03 | 0 | 04 |
| No | 08 | 14 | 16 | 38 |
| Sample Type | Algorithm Applied | Confusion Matrices of ML Algorithms 1 | No. of Features Identified 2 | Group | Precision | Recall | F1-Measure |
|---|---|---|---|---|---|---|---|
| Glioma vs. Control (n = 42) | Extra Tree Classifier | [16 0] | 104 | Control | 1.00 | 1.00 | 1.00 |
| [0 26] | Tumor | 1.00 | 1.00 | 1.00 | |||
| Logistic Regression | [16 0] | 01 | Control | 0.94 | 1.00 | 0.97 | |
| [1 25] | Tumor | 1.00 | 0.96 | 0.98 | |||
| Random Forest | [16 0] | 158 | Control | 1.00 | 1.00 | 1.00 | |
| [0 26] | Tumor | 1.00 | 1.00 | 1.00 | |||
| LGG vs. HGG (n = 25) | Extra Tree Classifier | [4 5] | 107 | LGG | 0.80 | 0.44 | 0.57 |
| [1 15] | HGG | 0.75 | 0.94 | 0.83 | |||
| Logistic Regression | [4 5] | 92 | LGG | 1.00 | 0.44 | 0.62 | |
| [0 16] | HGG | 0.76 | 1.00 | 0.86 | |||
| Random Forest | [2 7] | 88 | LGG | 0.67 | 0.22 | 0.33 | |
| [1 15] | HGG | 0.68 | 0.94 | 0.79 |
| Group 1: Glioma vs Control | ||||
|---|---|---|---|---|
| ML Analysis | Statistical Validation | |||
| Feature | Importance | Corresponding metabolite | p-value | log2(FC) |
| Model: Logistics Regression | ||||
| 1.47 | 1.0 | Alanine | <0.0001 | –3.7744 |
| Model: Extra Tree Classifier | ||||
| 2.55 | 0.0642 | β-Alanine | <0.0001 | −2.5717 |
| 2.12 | 0.0524 | Methionine | <0.0001 | −3.3458 |
| 3.1 | 0.0491 | Phenylalanine | <0.0001 | −7.3967 |
| 3.07 | 0.0415 | X | <0.0001 | −2.5445 |
| 2.69 | 0.0405 | NAA | <0.0001 | −4.7668 |
| 1.47 | 0.0383 | Alanine | <0.0001 | −3.7744 |
| 0.98 | 0.0299 | Valine | <0.0001 | −2.7959 |
| 1.71 | 0.028 | Leucine | <0.0001 | −3.5926 |
| 3.13 | 0.025 | Glutathione | 0.0013 | −5.7592 |
| 2.45 | 0.0222 | Glutamine | <0.0001 | −3.0169 |
| 1.88 | 0.022 | GABA | 0.0028 | −1.1886 |
| 3.95 | 0.0204 | Serine | 0.0003 | −2.4757 |
| 3.16 | 0.0182 | Alanine | <0.0001 | −5.5148 |
| 3.69 | 0.0179 | α-glucose | <0.0001 | −1.3382 |
| 1.44 | 0.0175 | Deoxycholic acid | <0.0001 | 10.114 |
| 3.18 | 0.0168 | Taurocholic acid | <0.0001 | −4.2704 |
| 4.11 | 0.0161 | Lactate | <0.0001 | 2.7555 |
| 1.72 | 0.0157 | Arginine | 0.0003 | −2.6918 |
| 3.58 | 0.0149 | Threonine | <0.0001 | −6.6301 |
| 2.35 | 0.0148 | Glutamate | 0.0022 | −1.4198 |
| 3.94 | 0.0143 | Serine | <0.0001 | −2.3249 |
| 3.14 | 0.014 | Spermidine | 0.0119 | −4.8047 |
| 3.88 | 0.014 | Aspartic acid | <0.0001 | 1.5565 |
| 2.09 | 0.0133 | Glutamate | <0.0001 | −1.859 |
| 3.21 | 0.0126 | N-Acetylcholine | 0.0004 | −4.3221 |
| 1.04 | 0.0126 | Valine | <0.0001 | −1.8092 |
| 1.01 | 0.0124 | Isoleucine | 0.0002 | 3.8182 |
| 1.89 | 0.0121 | GABA | 0.001 | −2.2659 |
| 3.68 | 0.0118 | α-glucose | <0.0001 | 4.8319 |
| 1.99 | 0.0117 | Isoleucine | 0.0006 | −1.2174 |
| Model: Random Forest | ||||
| 2.12 | 0.0491 | Methionine | <0.0001 | −3.3458 |
| 0.98 | 0.0488 | Valine | <0.0001 | −2.7959 |
| 3.16 | 0.0438 | Alanine | <0.0001 | −5.5148 |
| 4.11 | 0.0344 | Lactate | <0.0001 | 2.7555 |
| 1.47 | 0.0316 | Alanine | <0.0001 | −3.7744 |
| 3.53 | 0.03 | Myoinositol | 0.0003 | 1.0134 |
| 1.45 | 0.025 | Isoleucine | 0.0002 | 2.9475 |
| 1.44 | 0.0209 | Deoxycholic acid | <0.0001 | 10.114 |
| 1.99 | 0.0195 | Isoleucine | 0.0006 | −1.2174 |
| 2.69 | 0.019 | NAA | <0.0001 | −4.7668 |
| 2.32 | 0.0172 | Glutamate | 0.0003 | 2.1726 |
| 2.13 | 0.0172 | Glutamine | <0.0001 | −3.4879 |
| 2.45 | 0.0165 | Glutamine | <0.0001 | −3.0169 |
| 3.66 | 0.0157 | Isoleucine | 0.0050 | 4.9598 |
| 3.72 | 0.015 | β-glucose | 0.3716 | NA |
| 1.3 | 0.0146 | Fatty acids | <0.0001 | −1.3971 |
| 3.94 | 0.0144 | Serine | <0.0001 | −2.3249 |
| 3.69 | 0.0142 | α-glucose | <0.0001 | −1.3382 |
| 1.01 | 0.0142 | Isoleucine | 0.0002 | 3.8182 |
| 0.92 | 0.0139 | Isoleucine | 0.0002 | 2.3071 |
| 2.09 | 0.0133 | Glutamate | <0.0001 | −1.859 |
| 3.88 | 0.0117 | Aspartate | <0.0001 | 1.5565 |
| 3.59 | 0.0114 | L-Valine | 0.001‘ | −6.9614 |
| Age | 0.0107 | NA | NA | NA |
| 1.17 | 0.0105 | X | 0.006 | −2.41 |
| 1.88 | 0.0105 | GABA | 0.0028 | −1.1886 |
| 3.46 | 0.0097 | β-glucose | 0.0178 | NA |
| 3.22 | 0.0093 | Arginine | 0.0191 | NA |
| 3.58 | 0.0091 | Threonine | <0.0001 | −6.6301 |
| 3.47 | 0.0084 | β-glucose | 0.7007 | NA |
| Group 2: LGG vs HGG | ||||
| ML Analysis | Statistical Validation | |||
| Feature | Importance | Corresponding metabolite | p-value | log2(FC) |
| Model: Logistics Regression | ||||
| 3.51 | 0.2059 | Choline | 0.030 | NA |
| 2.01 | 0.1116 | NAA | 0.041 | 1.239 |
| 3.2 | 0.0719 | P-Choline | 0.127 | NA |
| 1.02 | 0.0531 | Valine | 0.040 | NA |
| 3.48 | 0.0523 | β-Glucose | 0.040 | NA |
| 2.39 | 0.0442 | Succinate/Malate | 0.040 | −1.1293 |
| 1.68 | 0.0441 | L-Arginine | 0.227 | 1.678 |
| 1.82 | 0.0435 | X | 0.015 | 1.4804 |
| 2.4 | 0.0416 | Succinate | 0.092 | NA |
| 2.31 | 0.0386 | X | 0.207 | −1.647 |
| 3.5 | 0.0324 | NAA | 0.133 | −2.3824 |
| 2.3 | 0.0311 | GABA | 0.133 | −3.0367 |
| 1.84 | 0.026 | X | 0.054 | 1.6947 |
| 3.53 | 0.026 | Myoinositol | 0.871 | NA |
| 1.26 | 0.0236 | Isoleucine | 0.064 | −2.4499 |
| 1.99 | 0.0171 | Isoleucine | 0.039 | NA |
| 2.45 | 0.0156 | L-Glutamine | 0.195 | NA |
| 0.88 | 0.015 | Fatty acid | 0.239 | −2.0748 |
| 3.91 | 0.0149 | Creatine | 0.054 | 1.2722 |
| 1.21 | 0.0125 | X | 0.054 | 3.6568 |
| 2.25 | 0.0122 | Fatty acid | 0.206 | 2.6199 |
| 0.9 | 0.012 | Fatty acid | 0.041 | 1.6497 |
| 3.26 | 0.0028 | Taurine | 0.009 | 1.1339 |
| 1.83 | 0 | X | 0.388 | −1.791 |
| 1.63 | 0 | X | 0.195 | NA |
| 1.86 | 0 | GABA | 0.206 | 1.9942 |
| 1.97 | 0 | Isoleucine | 0.182 | 1.679 |
| 1.47 | 0 | Alanine | 0.640 | 3.2255 |
| 1.87 | 0 | GABA | 0.195 | NA |
| 1.88 | 0 | GABA | 0.195 | NA |
| Model: Extra Tree Classifier | ||||
| 3.6 | 0.0637 | Valine | 0.015 | 1.5337 |
| 1.82 | 0.0352 | X | 0.015 | 1.4804 |
| 2.5 | 0.0265 | NAA | 0.015 | 1.3842 |
| 1.4 | 0.0254 | X | 0.071 | 1.5535 |
| 2.4 | 0.0254 | Succinate | 0.092 | 1.1227 |
| 1.97 | 0.0247 | Isoleucine | 0.182 | 1.679 |
| 2.04 | 0.0244 | Glutamate | 0.030 | 1.4984 |
| 0.99 | 0.0237 | Isoleucine | 0.104 | 1.2242 |
| 2.53 | 0.0232 | X | 0.015 | 2.0478 |
| 3.26 | 0.0232 | Taurine | 0.009 | 1.1339 |
| 2.08 | 0.0211 | Glutamate | 0.036 | NA |
| 3.59 | 0.0207 | Threonine | 0.053 | NA |
| 0.91 | 0.0186 | Fatty acids | 0.011 | 1.6453 |
| 0.98 | 0.017 | Valine | 0.249 | 1.7767 |
| 1.99 | 0.0167 | Isoleucine | 0.039 | 1.429 |
| 0.9 | 0.0167 | Fatty acids | 0.041 | 1.6497 |
| 3.33 | 0.0164 | Scyllo inositol | 0.015 | 1.5207 |
| 2.43 | 0.0162 | Glutamine | 0.222 | NA |
| 1.22 | 0.016 | X | 0.103 | NA |
| 3.91 | 0.0159 | Creatine | 0.054 | 1.2722 |
| 2.72 | 0.0154 | NAA | 0.136 | 2.2715 |
| 2.84 | 0.0149 | X | 0.054 | 1.2673 |
| 3.51 | 0.0144 | Choline | 0.030 | |
| 0.85 | 0.0143 | Tauro-cholicacid | 0.726 | −1.0208 |
| 1.68 | 0.0137 | Leucine | 0.227 | 1.678 |
| 3.54 | 0.0131 | Glycine | 0.050 | 1.1865 |
| 1.36 | 0.0127 | Fatty acids | 0.519 | NA |
| 3.42 | 0.0124 | Taurine/Proline | 0.031 | NA |
| 1.05 | 0.0123 | Valine | 0.097 | 1.5722 |
| 2.32 | 0.0119 | Glutamate | 0.026 | NA |
| Model: Random Forest | ||||
| 3.6 | 0.0637 | Valine | 0.015 | 1.5337 |
| 1.82 | 0.0352 | X | 0.015 | 1.4804 |
| 2.5 | 0.0265 | NAA | 0.015 | 1.3842 |
| 1.4 | 0.0254 | X | 0.071 | 1.5535 |
| 2.4 | 0.0254 | Succinate | 0.092 | 1.1227 |
| 1.97 | 0.0247 | Isoleucine | 0.182 | 1.679 |
| 2.04 | 0.0244 | Glutamate | 0.030 | 1.4984 |
| 0.99 | 0.0237 | Isoleucine | 0.104 | 1.2242 |
| 2.53 | 0.0232 | X | 0.015 | 2.0478 |
| 3.26 | 0.0232 | Taurine | 0.009 | 1.1339 |
| 2.08 | 0.0211 | Glutamate | 0.036 | NA |
| 3.59 | 0.0207 | Threonine | 0.053 | NA |
| 0.91 | 0.0186 | Fatty acids | 0.011 | 1.6453 |
| 0.98 | 0.017 | Valine | 0.249 | 1.7767 |
| 1.99 | 0.0167 | Isoleucine | 0.039 | 1.429 |
| 0.9 | 0.0167 | Fatty acids | 0.041 | NA |
| 3.33 | 0.0164 | scyllo inositol | 0.015 | 1.5207 |
| 2.43 | 0.0162 | Glutamine | 0.222 | NA |
| 1.22 | 0.016 | X | 0.103 | NA |
| 3.91 | 0.0159 | Creatine | 0.054 | 1.2722 |
| 2.72 | 0.0154 | NAAG | 0.136 | 2.2715 |
| 2.84 | 0.0149 | X | 0.054 | 1.2673 |
| 3.51 | 0.0144 | Choline | 0.030 | NA |
| 0.85 | 0.0143 | Tauro-cholicacid | 0.726 | −1.0208 |
| 1.68 | 0.0137 | Leucine | 0.227 | 1.678 |
| 3.54 | 0.0131 | Glycine | 0.050 | 1.1865 |
| 1.36 | 0.0127 | Fatty acids | 0.519 | NA |
| 3.42 | 0.0124 | Taurine/Proline | 0.031 | NA |
| 1.05 | 0.0123 | Valine | 0.097 | 1.5722 |
| 2.32 | 0.0119 | Glutamate | 0.026 | NA |
| Sr.# | Pathway | Glioma vs. Control | LGG vs. HGG | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Hits | RawP | FDR | Impact | Hits | RawP | FDR | Impact | ||
| 1 | Alanine, aspartate, and glutamate metabolism | 06//28 | 2.13 × 10−6 | 8.34 × 10−5 | 0.70754 | 06//28 | 4.57 × 10−7 | 1.92 × 10−5 | 0.48398 |
| 2 | D-Glutamine and D-glutamate metabolism | 2//6 | 0.00332 | 0.03984 | 0.5 | 2//6 | 0.00207 | 0.01937 | 0.5 |
| 3 | Arginine biosynthesis | 4//14 | 4.00 × 10−5 | 0.00084 | 0.19289 | 3//14 | 0.00052 | 0.00739 | 0.19289 |
| 4 | Glutathione metabolism | 3//28 | 0.00829 | 0.08708 | 0.28281 | 2//28 | 0.04453 | 1 | 0.12939 |
| 5 | Aminoacyl-tRNA biosynthesis | 13//48 | 8.36 × 10−15 | 7.02 × 10−13 | 0.16667 | 10//48 | 2.30 × 10−11 | 1.93 × 10−9 | 0 |
| 6 | Glycine, serine, and threonine metabolism | 2//33 | 0.09068 | 0.40088 | 0.21707 | 4//33 | 0.00053 | 0.00739 | 0.24577 |
| 7 | Arginine and proline metabolism | 4//38 | 0.0023 | 0.0386 | 0.20172 | 5//38 | 6.13 × 10−5 | 0.00129 | 0.25763 |
| 8 | Phenylalanine, tyrosine, and tryptophan biosynthesis | 1//4 | 0.06057 | 0.31497 | 0.5 | NA | |||
| 9 | Tryptophan metabolism | 1//41 | 0.47705 | 1 | 0.14305 | NA | |||
| 10 | Phenylalanine metabolism | 1//10 | 0.14488 | 0.55316 | 0.35714 | NA | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Firdous, S.; Abid, R.; Nawaz, Z.; Bukhari, F.; Anwer, A.; Cheng, L.L.; Sadaf, S. Dysregulated Alanine as a Potential Predictive Marker of Glioma—An Insight from Untargeted HRMAS-NMR and Machine Learning Data. Metabolites 2021, 11, 507. https://doi.org/10.3390/metabo11080507
Firdous S, Abid R, Nawaz Z, Bukhari F, Anwer A, Cheng LL, Sadaf S. Dysregulated Alanine as a Potential Predictive Marker of Glioma—An Insight from Untargeted HRMAS-NMR and Machine Learning Data. Metabolites. 2021; 11(8):507. https://doi.org/10.3390/metabo11080507
Chicago/Turabian StyleFirdous, Safia, Rizwan Abid, Zubair Nawaz, Faisal Bukhari, Ammar Anwer, Leo L. Cheng, and Saima Sadaf. 2021. "Dysregulated Alanine as a Potential Predictive Marker of Glioma—An Insight from Untargeted HRMAS-NMR and Machine Learning Data" Metabolites 11, no. 8: 507. https://doi.org/10.3390/metabo11080507
APA StyleFirdous, S., Abid, R., Nawaz, Z., Bukhari, F., Anwer, A., Cheng, L. L., & Sadaf, S. (2021). Dysregulated Alanine as a Potential Predictive Marker of Glioma—An Insight from Untargeted HRMAS-NMR and Machine Learning Data. Metabolites, 11(8), 507. https://doi.org/10.3390/metabo11080507