
Metabolomic Markers of Storage Temperature and Time in Pasteurized Milk



Abstract
:1. Introduction
2. Results and Discussion
2.1. Multivariate Analysis
2.2. Supervised Untargeted Analysis
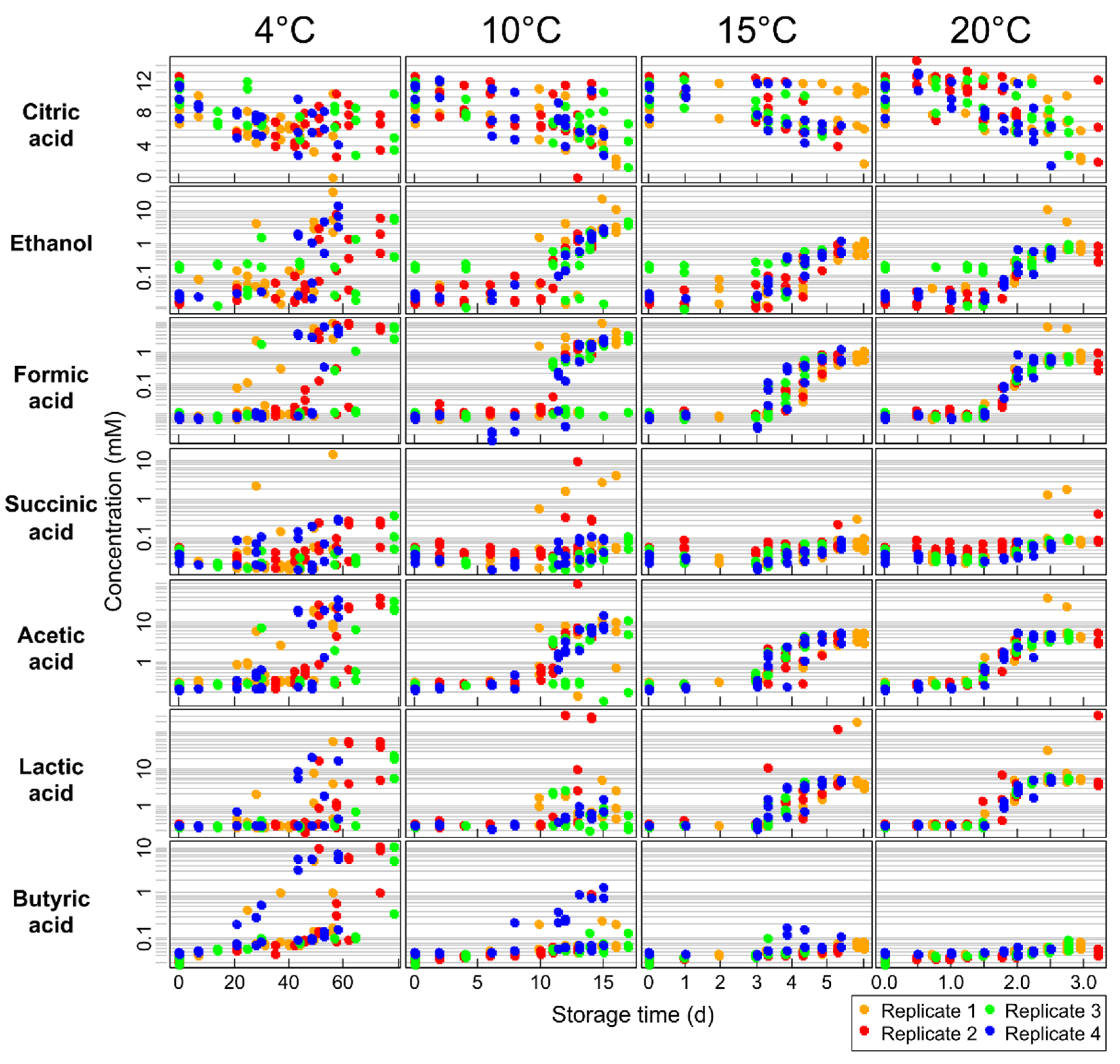
2.3. Targeted Analysis
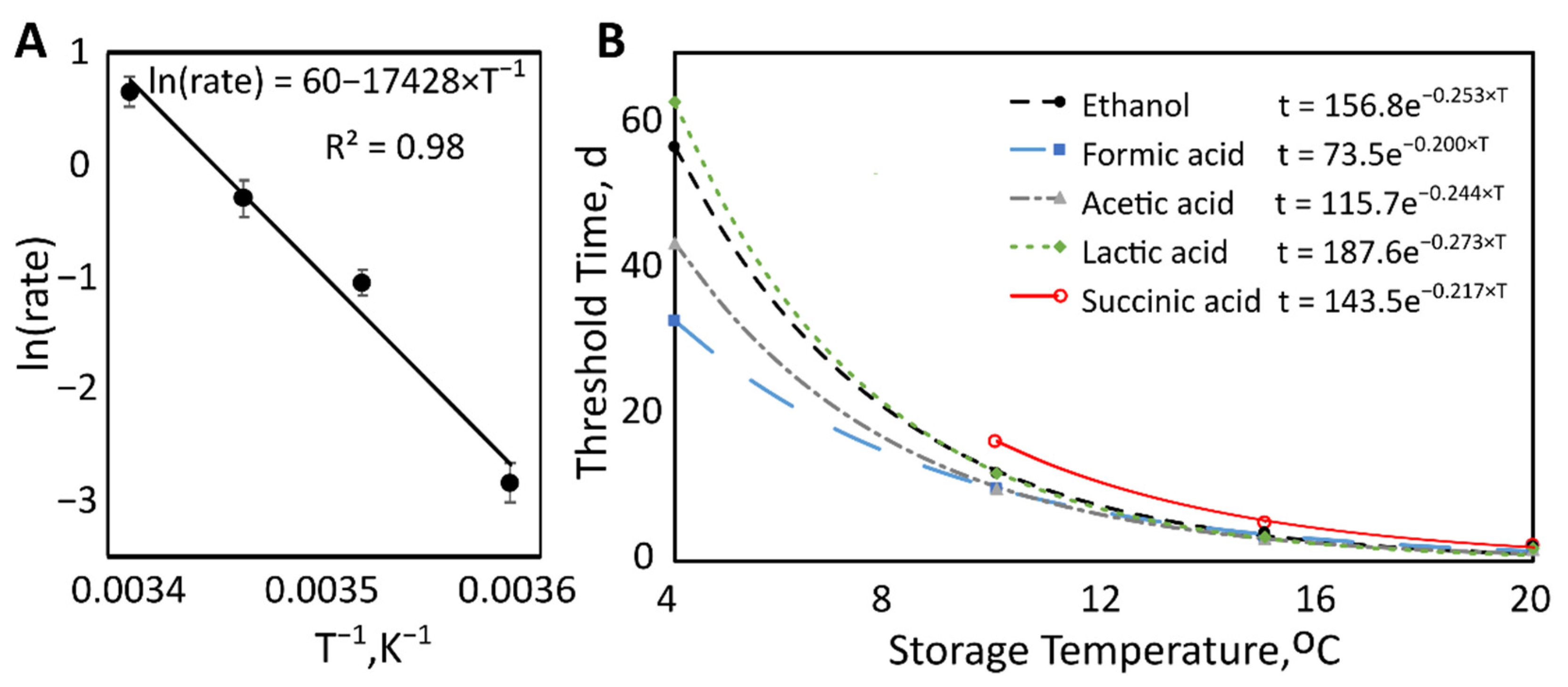
2.4. Effect of Storage Temperature
2.5. Effect of Milk Batch
2.6. Citric Acid
2.7. Products of Lactose Fermentation
2.7.1. Ethanol
2.7.2. Acetic Acid
2.7.3. Formic Acid
2.7.4. Lactic Acid
2.8. Succinic Acid
2.9. Free Butyric Acid
3. Materials and Methods
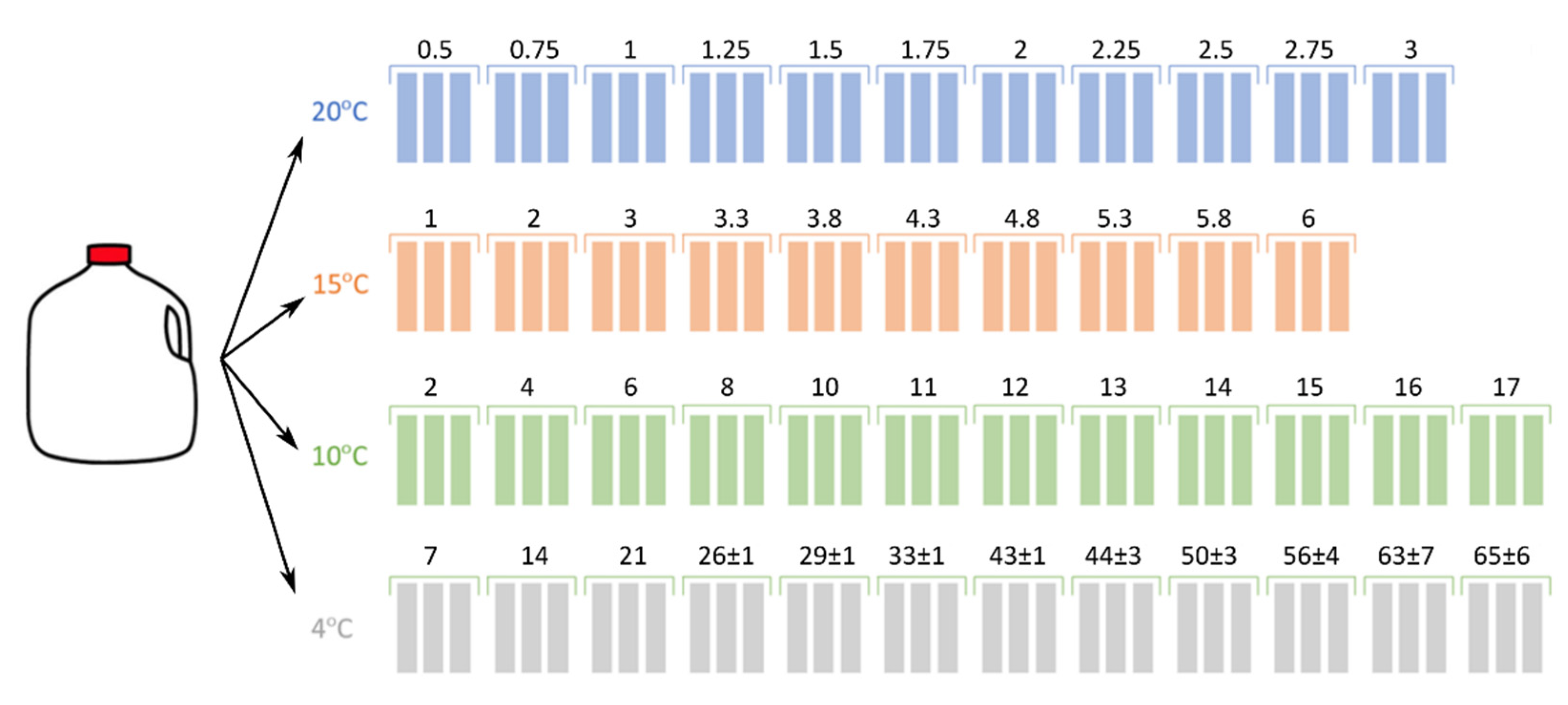
3.1. Sample Collection and Preparation
3.2. Nuclear Magnetic Resonance Spectroscopy
3.3. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food Outlook—Biannual Report on Global Food Markets; FAO: Rome, Italy, 2019. [Google Scholar]
- OECD/FAO. OECD-FAO Agricultural Outlook 2019–2028; OECD Publishing: Paris, France, 2019. [Google Scholar]
- FAO. Milk Production. Gateway to Dairy Production and Products. 2020. Available online: http://www.fao.org/dairy-production-products/production/en (accessed on 1 February 2020).
- USDA. U.S. Milk Production and Related Data (Quarterly and Annual); USDA: Washington, DC, USA, 2019.
- USDA. Estimated Fluid Milk Products Sales Report; USDA: Washington, DC, USA, 2019.
- Buzby, J.C.; Wells, H.F.; Hyman, J. The estimated amount, value, and calories of postharvest food losses at the retail and consumer levels in the United States. USDA Econ. Inf. Bull. 2014, 121. [Google Scholar] [CrossRef] [Green Version]
- Newsome, R.; Balestrini, C.G.; Baum, M.D.; Corby, J.; Fisher, W.; Goodburn, K.; Labuza, T.P.; Prince, G.; Thesmar, H.S.; Yiannas, F. Applications and perceptions of date labeling of food. Compr. Rev. Food Sci. Food Saf. 2014, 13, 745–769. [Google Scholar] [CrossRef] [Green Version]
- Roe, B.E.; Phinney, D.M.; Simons, C.T.; Badiger, A.S.; Bender, K.E.; Heldman, D.R. Discard intentions are lower for milk presented in containers without date labels. Food Qual. Prefer. 2018, 66, 13–18. [Google Scholar] [CrossRef]
- Ranieri, M.; Boor, K. Short communication: Bacterial ecology of high-temperature, short-time pasteurized milk processed in the United States. J. Dairy Sci. 2009, 92, 4833–4840. [Google Scholar] [CrossRef]
- Vallejo-Cordoba, B.; Nakai, S. Keeping-Quality Assessment of Pasteurized Milk by Multivariate Analysis of Dynamic Headspace Gas Chromatographic Data. 2. Flavor Classification by Linear Discriminant Analysis. J. Agric. Food Chem. 1994, 42, 994–999. [Google Scholar] [CrossRef]
- De Jonghe, V.; Coorevits, A.; De Block, J.; Van Coillie, E.; Grijspeerdt, K.; Herman, L.; De Vos, P.; Heyndrickx, M. Toxinogenic and spoilage potential of aerobic spore-formers isolated from raw milk. Int. J. Food Microbiol. 2010, 136, 318–325. [Google Scholar] [CrossRef]
- Lu, M.; Wang, N.S. Spoilage of milk and dairy products. Microbiol. Qual. Food 2017, 151–178. [Google Scholar] [CrossRef]
- Muir, D.D. The shelf-life of dairy products: 1. Factors influencing raw milk and fresh products. Int. J. Dairy Technol. 1996, 49, 24–32. [Google Scholar] [CrossRef]
- Chrisope, G.L.; Marshall, R.T. Combined Action of lipase and microbial phospholipase C on a model fat globule emulsion and raw milk. J. Dairy Sci. 1976, 59, 2024–2030. [Google Scholar] [CrossRef]
- Deeth, H.C.; Fitz-Gerald, C.H. Lipolytic Enzymes and Hydrolytic Rancidity in Milk and Milk Products. Dev. Dairy Chem. 1983, 2, 195–239. [Google Scholar] [CrossRef]
- Champagne, C.P.; Laing, R.R.; Roy, D.; Mafu, A.A.; Griffiths, M.W.; White, C. Psychrotrophs in dairy products: Their effects and their control. Crit. Rev. Food Sci. Nutr. 1994, 34, 1–30. [Google Scholar] [CrossRef]
- Baker, S. The keeping quality of refrigerated pasteurized milk. Aust. J. Dairy Technol. 1983, 38, 124–127. [Google Scholar]
- Maxcy, R.B.; Wallen, S.E. Heterogeneity of samples as a problem in shelf-life prediction. J. Food Prot. 1983, 46, 542–544. [Google Scholar] [CrossRef]
- Alothman, M.; Lusk, K.A.; Silcock, P.; Bremer, P.J. Comparing PTR-MS profile of milk inoculated with pure or mixed cultures of spoilage bacteria. Food Microbiol. 2017, 64, 155–163. [Google Scholar] [CrossRef]
- Ghaani, M.; Cozzolino, C.A.; Castelli, G.; Farris, S. An overview of the intelligent packaging technologies in the food sector. Trends Food Sci. Technol. 2016, 51, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Taoukis, P.S.; Labuza, T.P. Time-temperature indicators (TTIs). In Novel Food Packaging Techniques; Woodhead Publishing Limited: Cambridge, UK, 2003; pp. 103–126. [Google Scholar] [CrossRef]
- De Abreu, D.A.P.; Cruz, J.M.; Losada, P.P. Active and intelligent packaging for the food industry. Food Rev. Int. 2012, 28, 146–187. [Google Scholar] [CrossRef]
- Wells, J.H.; Singh, R.P. Application of time-temperature indicators in monitoring changes in quality attributes of perishable and semiperishable foods. J. Food Sci. 1988, 53, 148–152. [Google Scholar] [CrossRef]
- Grisius, R.; Wells, J.H.; Barrett, E.L.; Singh, R.P. Correlation of time-temperature indicator response with microbial growth in pasteurized milk. J. Food Process. Preserv. 1987, 11, 309–324. [Google Scholar] [CrossRef]
- Ziyaina, M.; Rasco, B.; Coffey, T.; Ünlü, G.; Sablani, S.S. Colorimetric detection of volatile organic compounds for shelf-life monitoring of milk. Food Control 2019, 100, 220–226. [Google Scholar] [CrossRef]
- Alothman, M.; Lusk, K.A.; Silcock, P.J.; Bremer, P.J. Relationship between total microbial numbers, volatile organic compound composition, and the sensory characteristics of whole fresh chilled pasteurized milk. Food Packag. Shelf Life 2018, 15, 69–75. [Google Scholar] [CrossRef]
- Silcock, P.; Alothman, M.; Zardin, E.; Heenan, S.P.; Siefarth, C.; Bremer, P.J.; Beauchamp, J. Microbially induced changes in the volatile constituents of fresh chilled pasteurised milk during storage. Food Packag. Shelf Life 2014, 2, 81–90. [Google Scholar] [CrossRef]
- Ziyaina, M.; Rasco, B.; Coffey, T.; Mattinson, D.S.; Sablani, S. Correlation of volatile compound concentrations with bacterial counts in whole pasteurised milk under various storage conditions. Int. J. Dairy Technol. 2019, 72, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Karatapanis, A.E.; Badeka, A.V.; Riganakos, K.A.; Savvaidis, I.N.; Kontominas, M.G. Changes in flavour volatiles of whole pasteurized milk as affected by packaging material and storage time. Int. Dairy J. 2006, 16, 750–761. [Google Scholar] [CrossRef]
- Sundekilde, U.K.; Larsen, L.B.; Bertram, H.C. NMR-Based Milk Metabolomics. Metabolites 2013, 3, 204–222. [Google Scholar] [CrossRef]
- Hu, F.; Furihata, K.; Ito-Ishida, M.; Kaminogawa, S.; Tanokura, M. Nondestructive observation of bovine milk by NMR spectroscopy: Analysis of existing states of compounds and detection of new compounds. J. Agric. Food Chem. 2004, 52, 4969–4974. [Google Scholar] [CrossRef] [PubMed]
- Sundekilde, U.K.; Frederiksen, P.D.; Clausen, M.R.; Larsen, L.B.; Bertram, H.C. Relationship between the metabolite profile and technological properties of bovine milk from two dairy breeds elucidated by NMR-based metabolomics. J. Agric. Food Chem. 2011, 59, 7360–7367. [Google Scholar] [CrossRef] [PubMed]
- Sacco, D.; Brescia, M.; Sgaramella, A.; Casiello, G.; Buccolieri, A.; Ogrinc, N.; Sacco, A. Discrimination between Southern Italy and foreign milk samples using spectroscopic and analytical data. Food Chem. 2009, 114, 1559–1563. [Google Scholar] [CrossRef]
- Lamanna, R.; Braca, A.; Di Paolo, E.; Imparato, G. Identification of milk mixtures by 1 H NMR profiling. Magn. Reson. Chem. 2011, 49, S22–S26. [Google Scholar] [CrossRef] [PubMed]
- Belloque, J.; Carrascosa, A.V.; López-Fandiño, R. Changes in Phosphoglyceride Composition during Storage of Ultrahigh-Temperature Milk, as Assessed by 31P-Nuclear Magnetic Resonance: Possible Involvement of Thermoresistant Microbial Enzymes. J. Food Prot. 2001, 64, 850–855. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.S.; Almstetter, M.F.; Schlamberger, G.; Nürnberger, N.; Dettmer, K.; Oefner, P.J.; Meyer, H.H.D.; Wiedemann, S.; Gronwald, W. Nuclear magnetic resonance and mass spectrometry-based milk metabolomics in dairy cows during early and late lactation. J. Dairy Sci. 2010, 93, 1539–1550. [Google Scholar] [CrossRef]
- Tsioulpas, A.; Lewis, M.J.; Grandison, A.S. Effect of minerals on casein micelle stability of cows’ milk. J. Dairy Res. 2007, 74, 167–173. [Google Scholar] [CrossRef]
- Foroutan, A.; Guo, A.C.; Vazquez-Fresno, R.; Lipfert, M.; Zhang, L.; Zheng, J.; Badran, H.; Budinski, Z.; Mandal, R.; Ametaj, B.N.; et al. Chemical Composition of Commercial Cow’s Milk. J. Agric. Food Chem. 2019, 67, 4897–4914. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, A.; Brechany, E.Y.; Mabon, R.M.; Pollock, H.T. Seasonal changes in the fat composition and concentration of citrate and related metabolites in cows’ milk. J. Dairy Res. 1986, 53, 223–227. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Masson, L.L.; Lock, A.L.; Mottram, T.T. Variation of milk citrate with stage of lactation and de novo fatty acid synthesis in dairy cows. J. Dairy Sci. 2006, 89, 1604–1612. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.S.; Buttchereit, N.; Miemczyk, S.P.; Immervoll, A.-K.; Louis, C.; Wiedemann, S.; Junge, W.; Thaller, G.; Oefner, P.J.; Gronwald, W. NMR Metabolomic Analysis of Dairy Cows Reveals Milk Glycerophosphocholine to Phosphocholine Ratio as Prognostic Biomarker for Risk of Ketosis. J. Proteome Res. 2011, 11, 1373–1381. [Google Scholar] [CrossRef]
- Bosworth, A.W.; Prucha, M. The fermentation of citric acid in milk. J. Biol. Chem. 1910, 8, 479–482. [Google Scholar] [CrossRef]
- Ajmal, M.; Nadeem, M.; Imran, M.; Junaid, M. Lipid compositional changes and oxidation status of ultra-high temperature treated Milk. Lipids Health Dis. 2018, 17, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocaign-Bousquet, M.; Garrigues, C.; Loubiere, P.; Lindley, N.D. Physiology of pyruvate metabolism in Lactococcus lactis. Antonie Leeuwenhoek 1996, 70, 253–267. [Google Scholar] [CrossRef]
- Kaneuchi, C.; Seki, M.; Komagata, K. Production of succinic acid from citric acid and related acids by lactobacillus strains. Appl. Environ. Microbiol. 1988, 54, 3053–3056. [Google Scholar] [CrossRef] [Green Version]
- Starrenburg, M.J.C.; Hugenholtz, J. Citrate Fermentation by Lactococcus and Leuconostoc spp. Appl. Environ. Microbiol. 1991, 57, 3535–3540. [Google Scholar] [CrossRef] [Green Version]
- Villota, R.; Hawkes, J.G. Reaction Kinetics in Food Systems. In Handbook of Food Engineering; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Toso, B.; Procida, G.; Stefanon, B. Determination of volatile compounds in cows’ milk using headspace GC-MS. J. Dairy Res. 2002, 69, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Urbach, G.; Milne, T. The concentration of volatiles in pasteurized milk as a function of storage time and storage tem-perature—A possible indicator of keeping quality. Aust. J. Dairy Technol. 1987, 42, 53–58. [Google Scholar]
- Morales, P.; Feliu, I.; Fernández-García, E.; Nuñez, M. Volatile Compounds Produced in Cheese by Enterobacteriaceae Strains of Dairy Origin. J. Food Prot. 2004, 67, 567–573. [Google Scholar] [CrossRef]
- Lees, G.J.; Jago, G.R. Acetaldehyde: An intermediate in the formation of ethanol from glucose by lactic acid bacteria. J. Dairy Res. 1976, 43, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Pierami, R.; Stevenson, K. Detection of Metabolites Produced by Psychrotrophic Bacteria Growing in Milk. J. Dairy Sci. 1976, 59, 1010–1015. [Google Scholar] [CrossRef]
- Haugen, J.E.; Rudi, K.; Langsrud, S.; Bredholt, S. Application of gas-sensor array technology for detection and monitoring of growth of spoilage bacteria in milk: A model study. Anal. Chim. Acta 2006, 565, 10–16. [Google Scholar] [CrossRef]
- Valero, E.; Villamiel, M.; Miralles, B.; Sanz, J.; Martınez-Castro, I. Changes in flavour and volatile components during storage of whole and skimmed UHT milk. Food Chem. 2001, 72, 51–58. [Google Scholar] [CrossRef]
- Curioni, P.M.G.; Bosset, J.O. Key odorants in various cheese types as determined by gas chromatography-olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Harper, W.; Gould, I.; Hankinson, C.L. Observations on the Free Volatile Acids in Milk. J. Dairy Sci. 1961, 44, 1764–1765. [Google Scholar] [CrossRef]
- Ozçelik, S.; Kuley, E.; Özogul, F. Formation of lactic, acetic, succinic, propionic, formic and butyric acid by lactic acid bacteria. Food Sci. Technol. 2016, 73, 536–542. [Google Scholar] [CrossRef]
- Soukoulis, C.; Aprea, E.; Biasioli, F.; Cappellin, L.; Schuhfried, E.; Märk, T.D.; Gasperi, F. Proton transfer reaction time-of-flight mass spectrometry monitoring of the evolution of volatile compounds during lactic acid fermentation of milk. Rapid Commun. Mass Spectrom. 2010, 24, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Hickey, M.W.; Hillier, A.; Jago, G. Metabolism of pyruvate and citrate in lactobacilli. Aust. J. Biol. Sci. 1983, 36, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindmark-Månsson, H.; Fondén, R.; Pettersson, H.-E. Composition of Swedish dairy milk. Int. Dairy J. 2003, 13, 409–425. [Google Scholar] [CrossRef]
- Pasvolsky, R.; Zakin, V.; Ostrova, I.; Shemesh, M. Butyric acid released during milk lipolysis triggers biofilm formation of Bacillus species. Int. J. Food Microbiol. 2014, 181, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Machado, S.G.; Baglinière, F.; Marchand, S.; Van Coillie, E.; Vanetti, M.C.D.; De Block, J.; Heyndrickx, M. The biodiversity of the microbiota producing heat-resistant enzymes responsible for spoilage in processed bovine milk and dairy products. Front. Microbiol. 2017, 8, 302. [Google Scholar] [CrossRef] [Green Version]
- Belenguer, A.; Duncan, S.H.; Calder, A.G.; Holtrop, G.; Louis, P.; Lobley, G.E.; Flint, H.J. Two Routes of Metabolic Cross-Feeding between Bifidobacterium adolescentis and Butyrate-Producing Anaerobes from the Human Gut. Appl. Environ. Microbiol. 2006, 72, 3593–3599. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wang, Y.; You, C.; Ren, J.; Chen, W.; Zheng, H.; Liu, Z. Variation in Raw Milk Microbiota Throughout 12 Months and the Impact of Weather Conditions. Sci. Rep. 2018, 8, 2371. [Google Scholar] [CrossRef] [Green Version]
- Duranti, S.; Milani, C.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Sánchez, B.; Margolles, A.; et al. Evaluation of genetic diversity among strains of the human gut commensal Bifidobacterium adolescentis. Sci. Rep. 2016, 6, 23971. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Klein, M.S.; Oefner, P.J.; Gronwald, W. MetaboQuant: A tool combining individual peak calibration and outlier detection for accurate metabolite quantification in 1D1H and1H-13C HSQC NMR spectra. Biotechniques 2013, 54, 251–256. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Citric Acid | Ethanol | Formic Acid | Acetic Acid | Lactic Acid | Succinic Acid | Butyric Acid |
|---|---|---|---|---|---|---|---|
| Storage Time | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Temperature | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Replicate | 0.0031 | <0.0001 | 0.0144 | 0.0002 | <0.0001 | <0.0001 | <0.0001 |
| Time × Time | 0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0793 |
| Temp × Temp | 0.0514 | 0.0007 | <0.0001 | <0.0001 | 0.0001 | 0.0007 | 0.0008 |
| Time × Temp | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0007 | <0.0001 | <0.0001 |
| Time × Replicate | 0.5097 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0626 |
| Temp × Replicate | 0.2335 | 0.4694 | 0.0269 | 0.0674 | 0.5476 | 0.7023 | 0.1943 |
| Metabolite | Temperature (°C) | Threshold Time (d) | p-Value |
|---|---|---|---|
| Ethanol | 4 | 57 ± 7 | 0.035 |
| 10 | 12.01 ± 0.01 | 0.043 | |
| 15 | 3.82 ± 0.01 | 0.037 | |
| 20 | 1.98 ± 0.02 | 0.006 | |
| Formic acid | 4 | 33 ± 2 | 0.045 |
| 10 | 9.97 ± 0.01 | 0.021 | |
| 15 | 3.31 ± 0.01 | 0.007 | |
| 20 | 1.78 ± 0.01 | 0.008 | |
| Acetic acid | 4 | 44 ± 5 | 0.027 |
| 10 | 9.97 ± 0.01 | 0.008 | |
| 15 | 3.00 ± 0.03 | 0.023 | |
| 20 | 1.49 ± 0.02 | 0.002 | |
| Lactic acid | 4 | 63 ± 12 | 0.035 |
| 10 | 12.01 ± 0.01 | 0.043 | |
| 15 | 3.31 ± 0.01 | 0.037 | |
| 20 | 1.78 ± 0.01 | 0.006 | |
| Succinic acid | 4 | - | - |
| 10 | 16.5 ± 0.5 | 0.021 | |
| 15 | 5.33 ± 0.04 | 0.006 | |
| 20 | 2.23 ± 0.01 | 0.009 |
| Metabolite | Decay Rate (°C−1) | 95% Confidence Interval |
|---|---|---|
| Ethanol | −0.254 | −0.273 to −0.235 |
| Formic Acid | −0.200 | −0.211 to −0.190 |
| Acetic Acid | −0.244 | −0.256 to −0.231 |
| Lactic Acid | −0.273 | −0.290 to −0.255 |
| Succinic Acid | −0.217 | −0.243 to −0.191 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edwards, K.M.; Badiger, A.; Heldman, D.R.; Klein, M.S. Metabolomic Markers of Storage Temperature and Time in Pasteurized Milk. Metabolites 2021, 11, 419. https://doi.org/10.3390/metabo11070419
Edwards KM, Badiger A, Heldman DR, Klein MS. Metabolomic Markers of Storage Temperature and Time in Pasteurized Milk. Metabolites. 2021; 11(7):419. https://doi.org/10.3390/metabo11070419
Chicago/Turabian StyleEdwards, Kara M., Aishwarya Badiger, Dennis R. Heldman, and Matthias S. Klein. 2021. "Metabolomic Markers of Storage Temperature and Time in Pasteurized Milk" Metabolites 11, no. 7: 419. https://doi.org/10.3390/metabo11070419
APA StyleEdwards, K. M., Badiger, A., Heldman, D. R., & Klein, M. S. (2021). Metabolomic Markers of Storage Temperature and Time in Pasteurized Milk. Metabolites, 11(7), 419. https://doi.org/10.3390/metabo11070419