
Extracellular Vesicle Analysis by Paper Spray Ionization Mass Spectrometry



Abstract
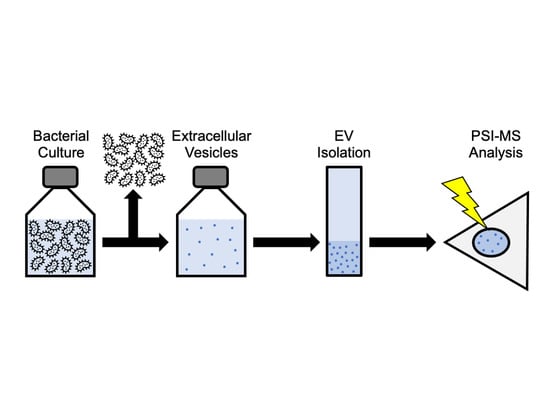
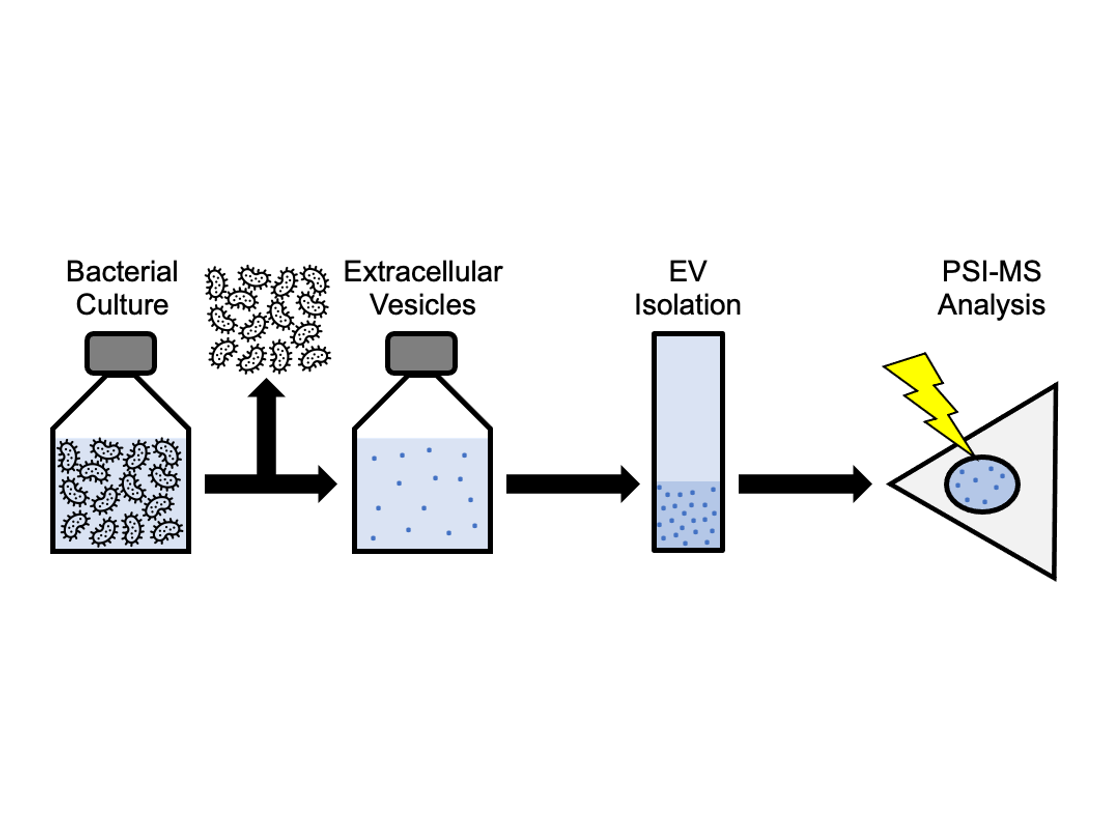
1. Introduction
1.1. Paper Spray Ionization and Bacterial Metabolomics
1.2. Bacterial Extracellular Vesicles
1.3. PSI-MS: A New Platform for Extracellular Vesicle Analysis
2. Results and Discussion
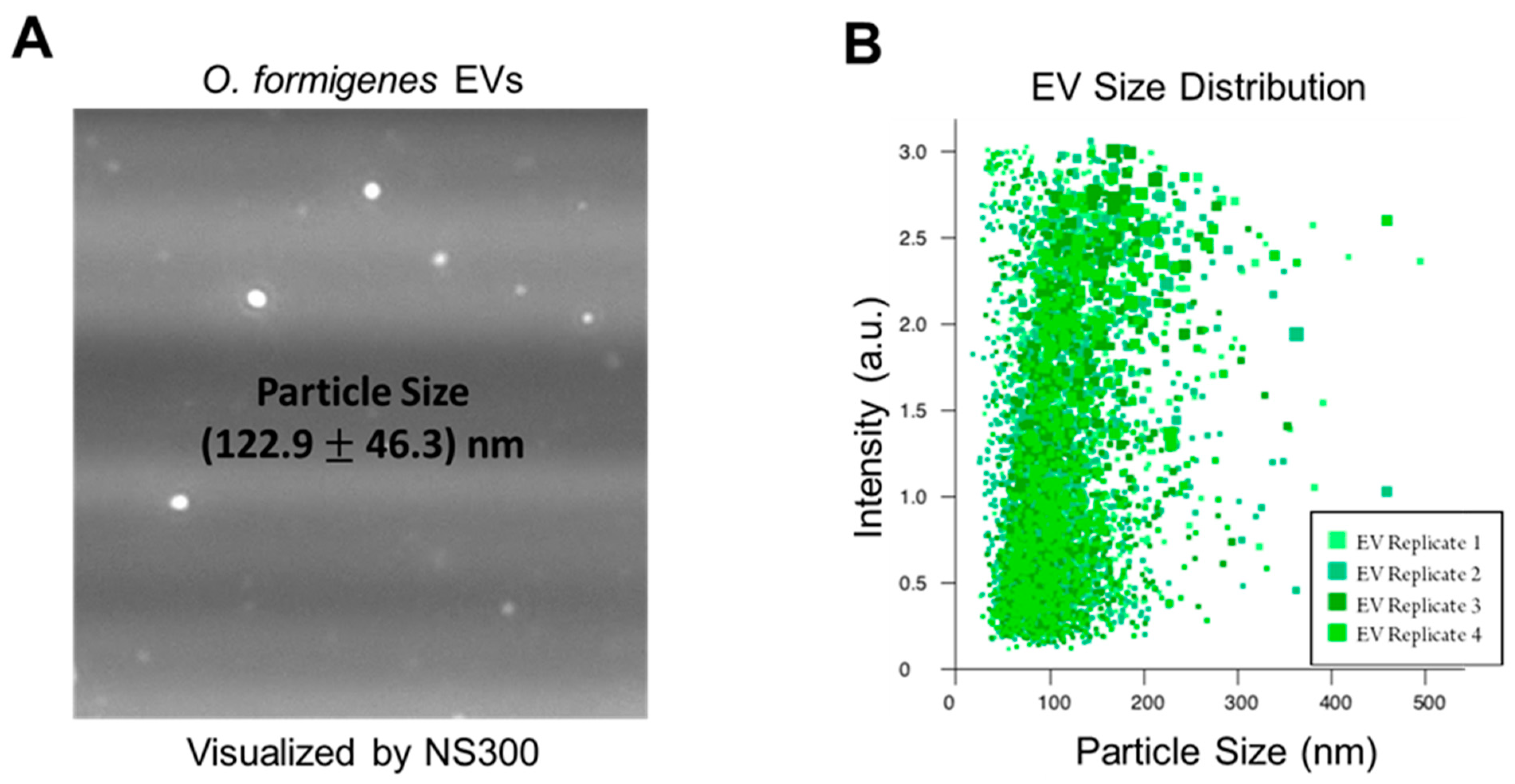
2.1. Nanoparticle Tracking Analysis Confirms O. formigenes Extracellular Vesicles
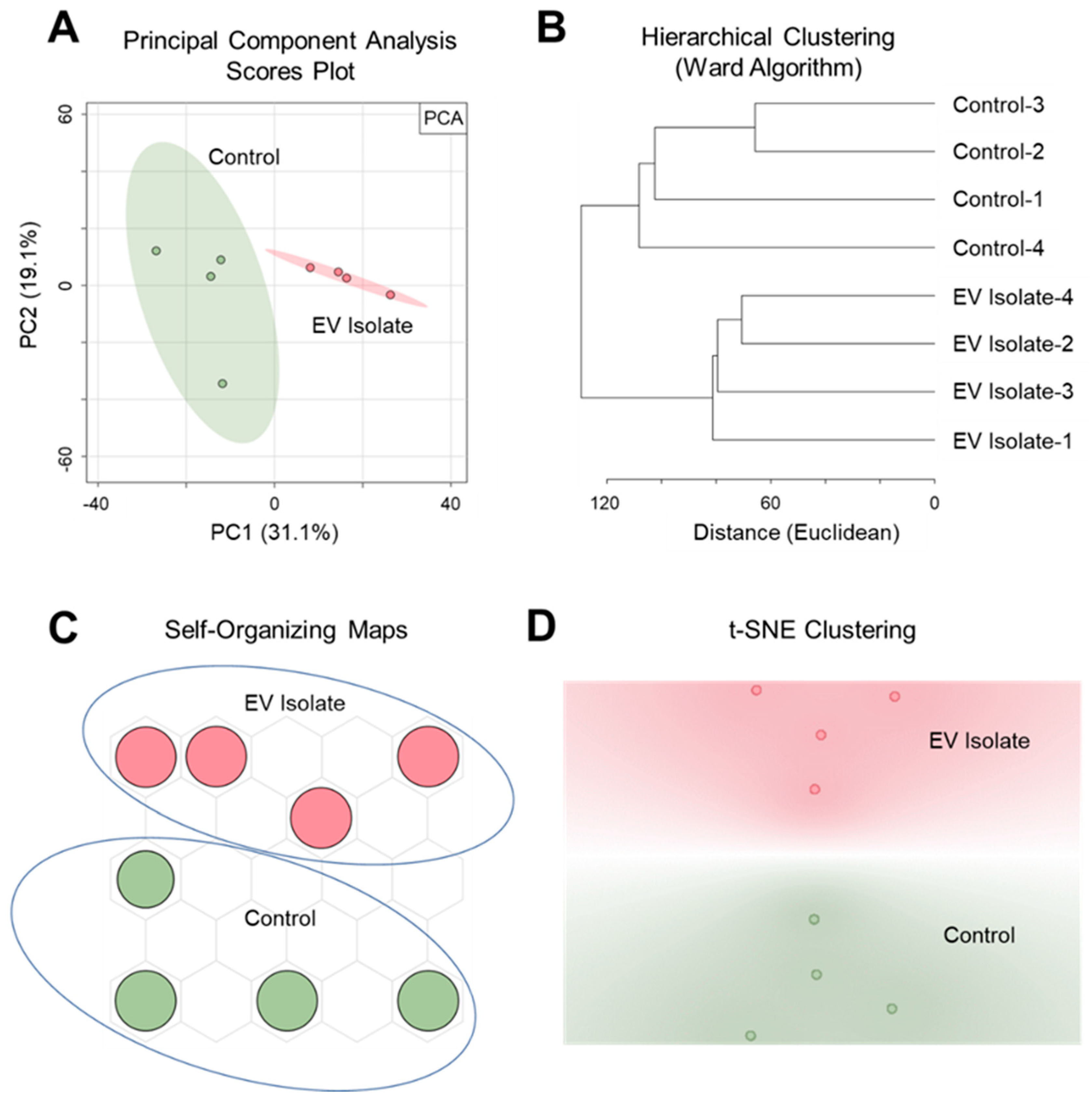
2.2. Extracellular Vesicle Metabolomics by PSI-MS
3. Materials and Methods
3.1. Isolation of O. formigenes Extracellular Vesicles from Culture Supernatant
3.2. Nanoparticle Tracking Analysis of O. formigenes Extracellular Vesicles
3.3. PSI-MS Instrumentation, Methodology, and Analysis
3.4. Data Processing, Statistics, and Feature Annotation
4. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, J.; Wang, H.; Manicke, N.E.; Lin, J.M.; Cooks, R.G.; Ouyang, Z. Development, characterization, and application of paper spray ionization. Anal. Chem. 2010, 82, 2463–2471. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, J.; Cooks, R.G.; Ouyang, Z. Paper spray for direct analysis of complex mixtures using mass spectrometry. Angew. Chem. Int. Ed. Engl. 2010, 49, 877–880. [Google Scholar] [CrossRef]
- Chiang, S.; Zhang, W.; Ouyang, Z. Paper spray ionization mass spectrometry: Recent advances and clinical applications. Expert Rev. Proteom. 2018, 15, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Dhummakupt, E.S.; Mach, P.M.; Carmany, D.; Demond, P.S.; Moran, T.S.; Connell, T.; Wylie, H.S.; Manicke, N.E.; Nilles, J.M.; Glaros, T. Direct Analysis of Aerosolized Chemical Warfare Simulants Captured on a Modified Glass-Based Substrate by “Paper-Spray” Ionization. Anal. Chem. 2017, 89, 10866–10872. [Google Scholar] [CrossRef] [PubMed]
- Wichert, W.R.; Dhummakupt, E.S.; Zhang, C.; Mach, P.M.; Bernhards, R.C.; Glaros, T.; Manicke, N.E. Detection of Protein Toxin Simulants from Contaminated Surfaces by Paper Spray Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2019, 30, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.M.; Jarmusch, A.K.; Pirro, V.; Pincus, D.H.; Clay, B.G.; Gervasi, G.; Cooks, R.G. Rapid discrimination of bacteria by paper spray mass spectrometry. Anal. Chem. 2014, 86, 7500–7507. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Yang, Q.; Zhong, X.; Yan, D.; Luo, W.; Fang, Z.; Xiao, J.; Ma, M.; Chen, B. Rapid analysis of anionic and cationic surfactants in water by paper spray mass spectrometry. Anal. Methods 2021, 13, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chang, Q.; Yin, K.; He, Q.; Deng, Y.; Chen, B.; Liu, C.; Wang, Y.; Wang, L. Rapid Analysis of Bisphenol A and Its Analogues in Food Packaging Products by Paper Spray Ionization Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 4859–4865. [Google Scholar] [CrossRef] [PubMed]
- Linhares, A.; Yonamine, M. Analysis of biofluids by paper spray-MS in forensic toxicology. Bioanalysis 2020, 12, 1087–1102. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.A.; Rubio, V.Y.; Garrett, T.J. Strain-Level Differentiation of Bacteria by Paper Spray Ionization Mass Spectrometry. Anal. Chem. 2019, 91, 4964–4968. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, C.J.; Wei, P.; Snyder, D.T.; Wang, X.; Ouyang, Z.; Pielak, R.M.; Graham Cooks, R. Rapid discrimination of bacteria using a miniature mass spectrometer. Analyst 2016, 141, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Anhalt, J.; Fenselau, C. Identification of Bacteria Using Mass-Spectrometry. Anal. Chem. 1975, 47, 219–225. [Google Scholar] [CrossRef]
- Liu, S.; Zuo, J.; Lu, Y.; Gao, L.; Zhai, Y.; Xu, W. Direct bacteria analysis using laserspray ionization miniature mass spectrometry. Anal. Bioanal. Chem. 2018, 411, 4031–4040. [Google Scholar] [CrossRef]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Bag, S.; Pulliam, C.; Snyder, D.; Pielak, R.; Cooks, R. Analysis of bacteria using zero volt paper spray. Anal. Methods 2016, 8, 1770–1773. [Google Scholar] [CrossRef]
- Hsu, C.C.; ElNaggar, M.S.; Peng, Y.; Fang, J.; Sanchez, L.M.; Mascuch, S.J.; Møller, K.A.; Alazzeh, E.K.; Pikula, J.; Quinn, R.A.; et al. Real-time metabolomics on living microorganisms using ambient electrospray ionization flow-probe. Anal. Chem. 2013, 85, 7014–7018. [Google Scholar] [CrossRef] [PubMed]
- Meetani, M.A.; Shin, Y.S.; Zhang, S.; Mayer, R.; Basile, F. Desorption electrospray ionization mass spectrometry of intact bacteria. J. Mass Spectrom. 2007, 42, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.D.; Wilkes, J.G.; Rafii, F.; Sutherland, J.B.; Persons, C.C.; Voorhees, K.J.; Lay, J.O. Rapid identification of intact whole bacteria based on spectral patterns using matrix-assisted laser desorption/ionization with time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 1996, 10, 1227–1232. [Google Scholar] [CrossRef]
- Pinu, F.R.; Villas-Boas, S.G. Extracellular Microbial Metabolomics: The State of the Art. Metabolites 2017, 7, 43. [Google Scholar] [CrossRef]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic profiling of oxalate-degrading probiotic Lactobacillus acidophilus and Lactobacillus gasseri. PLoS ONE 2019, 14, e0222393. [Google Scholar] [CrossRef]
- Behrends, V.; Williams, H.D.; Bundy, J.G. Metabolic footprinting: Extracellular metabolomic analysis. Methods Mol. Biol. 2014, 1149, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Maffei, B.; Francetic, O.; Subtil, A. Tracking Proteins Secreted by Bacteria: What’s in the Toolbox? Front. Cell. Infect. Microbiol. 2017, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Mashego, M.R.; van Gulik, W.M.; Heijnen, J.J. Metabolome dynamic responses of Saccharomyces cerevisiae to simultaneous rapid perturbations in external electron acceptor and electron donor. FEMS Yeast Res. 2007, 7, 48–66. [Google Scholar] [CrossRef] [PubMed]
- Peisl, B.Y.L.; Schymanski, E.L.; Wilmes, P. Dark matter in host-microbiome metabolomics: Tackling the unknowns—A review. Anal. Chim. Acta 2018, 1037, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 2019, 43, 273–303. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Defourny, K.A.Y.; Smid, E.J.; Abee, T. Gram-Positive Bacterial Extracellular Vesicles and Their Impact on Health and Disease. Front. Microbiol. 2018, 9, 1502. [Google Scholar] [CrossRef] [PubMed]
- Gaurivaud, P.; Ganter, S.; Villard, A.; Manso-Silvan, L.; Chevret, D.; Boulé, C.; Monnet, V.; Tardy, F. Mycoplasmas are no exception to extracellular vesicles release: Revisiting old concepts. PLoS ONE 2018, 13, e0208160. [Google Scholar] [CrossRef] [PubMed]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cruz, C.; Delgado, L.; López-Iglesias, C.; Mercade, E. Outer-inner membrane vesicles naturally secreted by gram-negative pathogenic bacteria. PLoS ONE 2015, 10, e0116896. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, J.; Park, J.; Gho, Y.S. Gram-negative and Gram-positive bacterial extracellular vesicles. Semin. Cell Dev. Biol. 2015, 40, 97–104. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A.J.; Johnson, A.P.; Vemulapalli, S.; Kuehn, M.J. Outer membrane vesicle production by Escherichia coli is independent of membrane instability. J. Bacteriol. 2006, 188, 5385–5392. [Google Scholar] [CrossRef] [PubMed]
- Dorward, D.W.; Garon, C.F.; Judd, R.C. Export and intercellular transfer of DNA via membrane blebs of Neisseria gonorrhoeae. J. Bacteriol. 1989, 171, 2499–2505. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, E.Y.; Kim, S.H.; Kim, D.K.; Park, K.S.; Kim, K.P.; Kim, Y.K.; Roh, T.Y.; Gho, Y.S. Staphylococcus aureus extracellular vesicles carry biologically active β-lactamase. Antimicrob. Agents Chemother. 2013, 57, 2589–2595. [Google Scholar] [CrossRef] [PubMed]
- Pocsfalvi, G.; Stanly, C.; Vilasi, A.; Fiume, I.; Capasso, G.; Turiák, L.; Buzas, E.I.; Vékey, K. Mass spectrometry of extracellular vesicles. Mass Spectrom. Rev. 2016, 35, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Rody, W.J.; Chamberlain, C.A.; Emory-Carter, A.K.; McHugh, K.P.; Wallet, S.M.; Spicer, V.; Krokhin, O.; Holliday, L.S. The proteome of extracellular vesicles released by clastic cells differs based on their substrate. PLoS ONE 2019, 14, e0219602. [Google Scholar] [CrossRef] [PubMed]
- Bandu, R.; Oh, J.W.; Kim, K.P. Mass spectrometry-based proteome profiling of extracellular vesicles and their roles in cancer biology. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Guo, W.B.; Zhang, W.S.; Bian, J.; Yang, J.K.; Zhou, Q.Z.; Chen, M.K.; Peng, W.; Qi, T.; Wang, C.Y.; et al. Comprehensive proteomics analysis of exosomes derived from human seminal plasma. Andrology 2017, 5, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Rody, W.J.; Krokhin, O.; Spicer, V.; Chamberlain, C.A.; Chamberlain, M.; McHugh, K.P.; Wallet, S.M.; Emory, A.K.; Crull, J.D.; Holliday, L.S. The use of cell culture platforms to identify novel markers of bone and dentin resorption. Orthod. Craniofac. Res. 2017, 20 (Suppl. 1), 89–94. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.S.; Kim, D.K.; Kim, Y.K.; Gho, Y.S. Proteomics of extracellular vesicles: Exosomes and ectosomes. Mass Spectrom. Rev. 2015, 34, 474–490. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Choi, D.S.; Kim, K.P.; Gho, Y.S. Proteomics in gram-negative bacterial outer membrane vesicles. Mass Spectrom. Rev. 2008, 27, 535–555. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, O.Y.; Gho, Y.S. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Proteom. Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Olaya-Abril, A.; Prados-Rosales, R.; McConnell, M.J.; Martín-Peña, R.; González-Reyes, J.A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Fernández, J.; Luque-García, J.L.; García-Lidón, C.; et al. Characterization of protective extracellular membrane-derived vesicles produced by Streptococcus pneumoniae. J. Proteom. 2014, 106, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Tsolakos, N.; Lie, K.; Bolstad, K.; Maslen, S.; Kristiansen, P.A.; Høiby, E.A.; Wallington, A.; Vipond, C.; Skehel, M.; Tang, C.M.; et al. Characterization of meningococcal serogroup B outer membrane vesicle vaccines from strain 44/76 after growth in different media. Vaccine 2010, 28, 3211–3218. [Google Scholar] [CrossRef] [PubMed]
- Murray, P. What Is New in Clinical Microbiology Microbial Identification by MALDI-TOF Mass Spectrometry A Paper from the 2011 William Beaumont Hospital Symposium on Molecular Pathology. J. Mol. Diagn. 2012, 14, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M. Gut microbiota and oxalate homeostasis. Ann. Transl. Med. 2017, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Whittamore, J.M.; Hatch, M. The role of intestinal oxalate transport in hyperoxaluria and the formation of kidney stones in animals and man. Urolithiasis 2017, 45, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic Alteration in the Mouse Distal Colonic Mucosa after Oral Gavage with Oxalobacter formigenes. Metabolites 2020, 10, 405. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.A. Metabolomic Characterization of the Intestinal Bacterial Oxalobiome; University of Florida: Gainesville, FL, USA, 2019. [Google Scholar]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Oxalobacter formigenes produces metabolites and lipids undetectable in oxalotrophic Bifidobacterium animalis. Metabolomics 2020, 16, 122. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.; Cornelius, J.; Allison, M.; Sidhu, H.; Peck, A.; Freel, R.W. Oxalobacter sp. reduces urinary oxalate excretion by promoting enteric oxalate secretion. Kidney Int. 2006, 69, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.; Gjymishka, A.; Salido, E.C.; Allison, M.J.; Freel, R.W. Enteric oxalate elimination is induced and oxalate is normalized in a mouse model of primary hyperoxaluria following intestinal colonization with Oxalobacter. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G461–G469. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.; Freel, R.W. A human strain of Oxalobacter (HC-1) promotes enteric oxalate secretion in the small intestine of mice and reduces urinary oxalate excretion. Urolithiasis 2013, 41, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Neven, K.Y.; Nawrot, T.S.; Bollati, V. Extracellular Vesicles: How the External and Internal Environment Can Shape Cell-To-Cell Communication. Curr. Environ. Health Rep. 2017, 4, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Metabolomic and lipidomic characterization of Oxalobacter formigenes strains HC1 and OxWR by UHPLC-HRMS. Anal. Bioanal. Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Klose, K.E. Increased chatter: Cyclic dipeptides as molecules of chemical communication in Vibrio spp. J. Bacteriol. 2006, 188, 2025–2026. [Google Scholar] [CrossRef]
- Holden, M.T.; Ram Chhabra, S.; de Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Boyer, M.; Wisniewski-Dyé, F. Cell-cell signalling in bacteria: Not simply a matter of quorum. FEMS Microbiol. Ecol. 2009, 70, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Candela, T.; Fouet, A. Poly-gamma-glutamate in bacteria. Mol. Microbiol. 2006, 60, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Aronov, P.A.; Luo, F.J.; Plummer, N.S.; Quan, Z.; Holmes, S.; Hostetter, T.H.; Meyer, T.W. Colonic contribution to uremic solutes. J. Am. Soc. Nephrol. 2011, 22, 1769–1776. [Google Scholar] [CrossRef]
- Poesen, R.; Claes, K.; Evenepoel, P.; de Loor, H.; Augustijns, P.; Kuypers, D.; Meijers, B. Microbiota-Derived Phenylacetylglutamine Associates with Overall Mortality and Cardiovascular Disease in Patients with CKD. J. Am. Soc. Nephrol. 2016, 27, 3479–3487. [Google Scholar] [CrossRef] [PubMed]
- Duran, M.; Ponezi, A.N.; Faljoni-Alario, A.; Teixeira, M.F.S.; Justo, G.Z.; Duran, N. Potential applications of violacein: A microbial pigment. Med. Chem. Res. 2012, 21, 1524–1532. [Google Scholar] [CrossRef]
- Hoshino, T. Violacein and related tryptophan metabolites produced by Chromobacterium violaceum: Biosynthetic mechanism and pathway for construction of violacein core. Appl. Microbiol. Biotechnol. 2011, 91, 1463–1475. [Google Scholar] [CrossRef]
- Dodou, H.V.; de Morais Batista, A.H.; Sales, G.W.P.; de Medeiros, S.C.; Rodrigues, M.L.; Nogueira, P.C.N.; Silveira, E.R.; Nogueira, N.A.P. Violacein antimicrobial activity on Staphylococcus epidermidis and synergistic effect on commercially available antibiotics. J. Appl. Microbiol. 2017, 123, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Andrighetti-Frohner, C.R.; Antonio, R.V.; Creczynski-Pasa, T.B.; Barardi, C.R.; Simoes, C.M. Cytotoxicity and potential antiviral evaluation of violacein produced by Chromobacterium violaceum. Memórias Inst. Oswaldo Cruz 2003, 98, 843–848. [Google Scholar] [CrossRef]
- Antonisamy, P.; Ignacimuthu, S. Immunomodulatory, analgesic and antipyretic effects of violacein isolated from Chromobacterium violaceum. Phytomedicine 2010, 17, 300–304. [Google Scholar] [CrossRef]
- Venegas, F.A.; Kollisch, G.; Mark, K.; Diederich, W.E.; Kaufmann, A.; Bauer, S.; Chavarria, M.; Araya, J.J.; Garcia-Pineres, A.J. The Bacterial Product Violacein Exerts an Immunostimulatory Effect via TLR8. Sci. Rep. 2019, 9, 13661. [Google Scholar] [CrossRef] [PubMed]
- Bachurski, D.; Schuldner, M.; Nguyen, P.H.; Malz, A.; Reiners, K.S.; Grenzi, P.C.; Babatz, F.; Schauss, A.C.; Hansen, H.P.; Hallek, M.; et al. Extracellular vesicle measurements with nanoparticle tracking analysis—An accuracy and repeatability comparison between NanoSight NS300 and ZetaView. J. Extracell Vesicles 2019, 8, 1596016. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Diedrich, J.; Chu, Y.Y.; Yates, J.R. Extracting Accurate Precursor Information for Tandem Mass Spectra by RawConverter. Anal. Chem. 2015, 87, 11361–11367. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Demsar, J.; Curk, T.; Erjavec, A.; Gorup, C.; Hocevar, T.; Milutinovic, M.; Mozina, M.; Polajnar, M.; Toplak, M.; Staric, A.; et al. Orange: Data Mining Toolbox in Python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Wehrens, R.; Hageman, J.A.; van Eeuwijk, F.; Kooke, R.; Flood, P.J.; Wijnker, E.; Keurentjes, J.J.; Lommen, A.; van Eekelen, H.D.; Hall, R.D.; et al. Improved batch correction in untargeted MS-based metabolomics. Metabolomics 2016, 12, 88. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| m/z | Annotation | Molecular Formula (M) | Ion/Adduct | Δppm | METLIN ID |
|---|---|---|---|---|---|
| 165.0153 a | Muconic Acid | C6H6O4 | [M + Na]+ | 3 | 45919 |
| 165.1016 | Kynuramine | C9H12N2O | [M + H]+ | 3 | 43923 |
| 183.0261 a | Oxoadipic Acid | C6H8O5 | [M + Na]+ | 1 | 322 |
| 185.1280 | Ala-Ile/Leu | C9H18N2O3 | [M + H – H2O]+ | 5 | 8560(6/7) |
| 196.0010 | 4-Phosphoaspartic Acid | C4H8NO7P | [M + H – H2O]+ | 0 | 360 |
| 205.1542 | 3-Hydroxy-N6,N6,N6-Trimethyl-L-Lysine | C9H20N2O3 | [M + H]+ | 2 | 6324 |
| 213.0364 a | 4-Hydroxy-4-methyl-2-Oxoadipic Acid | C7H10O6 | [M + Na]+ | 2 | 66102 |
| 213.1228 | Pro-Pro | C10H16N2O3 | [M + H]+ | 2 | 62027 |
| 218.1382 | Propionylcarnitine | C10H19NO4 | [M + H]+ | 2 | 965 |
| 227.1385 | Hydroxyprolyl-(Iso)Leucine | C11H20N2O4 | [M + H – H2O]+ | 4 | 8577(3/4) |
| 229.1180 | Prolylhydroxyproline | C10H16N2O4 | [M + H]+ | 1 | 58518 |
| 241.1177 | Gamma-Glutamyl-Pipecolic Acid | C11H18N2O5 | [M + H – H2O]+ | 4 | 93275 |
| 246.0731 a | Acetyltyrosine | C11H13NO4 | [M + Na]+ | 2 | 5827 |
| 251.0522 | Homocystine | C8H16N2O4S2 | [M + H – H2O]+ | 0 | 4189 |
| 254.1378 | 3-Indolecarboxylic Acid | C13H19NO4 | [M + H]+ | 3 | 6660 |
| 262.0851 | Ser-Ala-Cys | C9H17N3O5S | [M + H – H2O]+ | 4 | 15654 |
| 265.1168 | Phenylacetylglutamine | C13H16N2O4 | [M + H]+ | 5 | 58397 |
| 295.2238 a | Hydroxypalmitic Acid | C16H32O3 | [M + Na]+ | 1 | 35428 |
| 297.0483 | 5′-Phosphoribosyl-N-Formylglycinamide | C8H15N2O9P | [M + H – H2O]+ | 2 | 3443 |
| 311.1456 a | Arg-Asn | C10H20N6O4 | [M + Na]+ | 5 | 23959 |
| 317.1929 | Ala-Arg-Ala | C12H24N6O4 | [M + H]+ | 0 | 21376 |
| 326.0909 | Violacein | C20H13N3O3 | [M + H – H2O]+ | 5 | C21136 b |
| 337.1605 | Ala-Gln-His | C14H22N6O5 | [M + H – H2O]+ | 5 | 16023 |
| 345.1875 | Ser-Arg-Thr | C13H26N6O6 | [M + H – H2O]+ | 3 | 16028 |
| 359.1690 | Asp-Arg-Ser | C13H24N6O7 | [M + H – H2O]+ | 3 | 17672 |
| 361.1965 | Arg-Trp | C17H24N6O3 | [M + H]+ | 4 | 23686 |
| 367.1084 | Met-Cys-Asn | C12H22N4O5S2 | [M + H]+ | 5 | 15764 |
| 385.3061 | N-Palmitoyl Glutamine | C21H40N2O4 | [M + H]+ | 0 | 75509 |
| 407.2034 | Ser-Arg-Tyr | C18H28N6O6 | [M + H – H2O]+ | 2 | 15751 |
| 415.2289 | Gly-Lys-Asn-Pro | C17H30N6O6 | [M + H]+ | 2 | 146911 |
| 421.2315 | His-His-Lys | C18H28N8O4 | [M + H]+ | 2 | 18791 |
| 431.2394 | Phe-His-Lys | C21H30N6O4 | [M + H]+ | 1 | 18657 |
| 441.1496 | Cys-Met-Ser-Thr | C15H28N4O7S2 | [M + H]+ | 5 | 115796 |
| 445.1208 | Cys-Cys-Gly-Tyr | C17H24N4O6S2 | [M + H]+ | 0 | 111999 |
| 473.3075 | Ile/Leu-Lys-Asn-Val | C21H40N6O6 | [M + H]+ | 1 | 162916 |
| 475.2862 | Ala-Glu-Lys-Lys | C20H38N6O7 | [M + H]+ | 2 | 104848 |
| 479.1988 | Ala-Asp-His-His | C19H26N8O7 | [M + H]+ | 1 | 104406 |
| 501.1806 | Polyglutamic Acid | C20H30N4O12 | [M + H – H2O]+ | 5 | 58212 |
| 657.3238 | Gln-Arg-Trp-Trp | C33H42N10O6 | [M + H – H2O]+ | 3 | 213457 |
| 663.4264 | Phosphatidylglycerol (28:2) | C34H63O10P | [M + H]+ | 4 | 79745 |
| 670.5166 | Phosphatidylethanolamine (38:1) | C38H74NO7P | [M + H – H2O]+ | 1 | 60361 |
| 674.5555 | GlcCer(d18:0/14:0) | C38H75NO8 | [M + H]+ | 1 | 53987 |
| 734.5109 | Phosphatidylethanolamine (37:5) | C42H74NO8P | [M + H – H2O]+ | 2 | 60354 |
| 761.5136 | Phosphatidic Acid (41:7) | C44H73O8P | [M + H]+ | 2 | 81674 |
| 765.5086 | Phosphatidylglycerol (37:5) | C43H75O10P | [M + H – H2O]+ | 2 | 79015 |
| 769.5023 | Phosphatidylglycerol (36:5) | C42H73O10P | [M + H]+ | 1 | 61870 |
| 835.5720 | Phosphatidylinositol (35:0) | C44H85O13P | [M + H – H2O]+ | 2 | 80078 |
| 862.6525 | Phosphatidylserine (41:0) | C47H92NO10P | [M + H]+ | 0 | 78139 |
| 958.3124 | Pentaglutamyl Folic Acid | C39H47N11O18 | [M + H]+ | 5 | 58426 |
| 960.3109 | Tetradecanoyl-CoA | C35H62N7O17P3S | [M + H – H2O]+ | 0 | 3707 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chamberlain, C.A.; Hatch, M.; Garrett, T.J. Extracellular Vesicle Analysis by Paper Spray Ionization Mass Spectrometry. Metabolites 2021, 11, 308. https://doi.org/10.3390/metabo11050308
Chamberlain CA, Hatch M, Garrett TJ. Extracellular Vesicle Analysis by Paper Spray Ionization Mass Spectrometry. Metabolites. 2021; 11(5):308. https://doi.org/10.3390/metabo11050308
Chicago/Turabian StyleChamberlain, Casey A., Marguerite Hatch, and Timothy J. Garrett. 2021. "Extracellular Vesicle Analysis by Paper Spray Ionization Mass Spectrometry" Metabolites 11, no. 5: 308. https://doi.org/10.3390/metabo11050308
APA StyleChamberlain, C. A., Hatch, M., & Garrett, T. J. (2021). Extracellular Vesicle Analysis by Paper Spray Ionization Mass Spectrometry. Metabolites, 11(5), 308. https://doi.org/10.3390/metabo11050308