
A Medicago truncatula Metabolite Atlas Enables the Visualization of Differential Accumulation of Metabolites in Root Tissues



Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
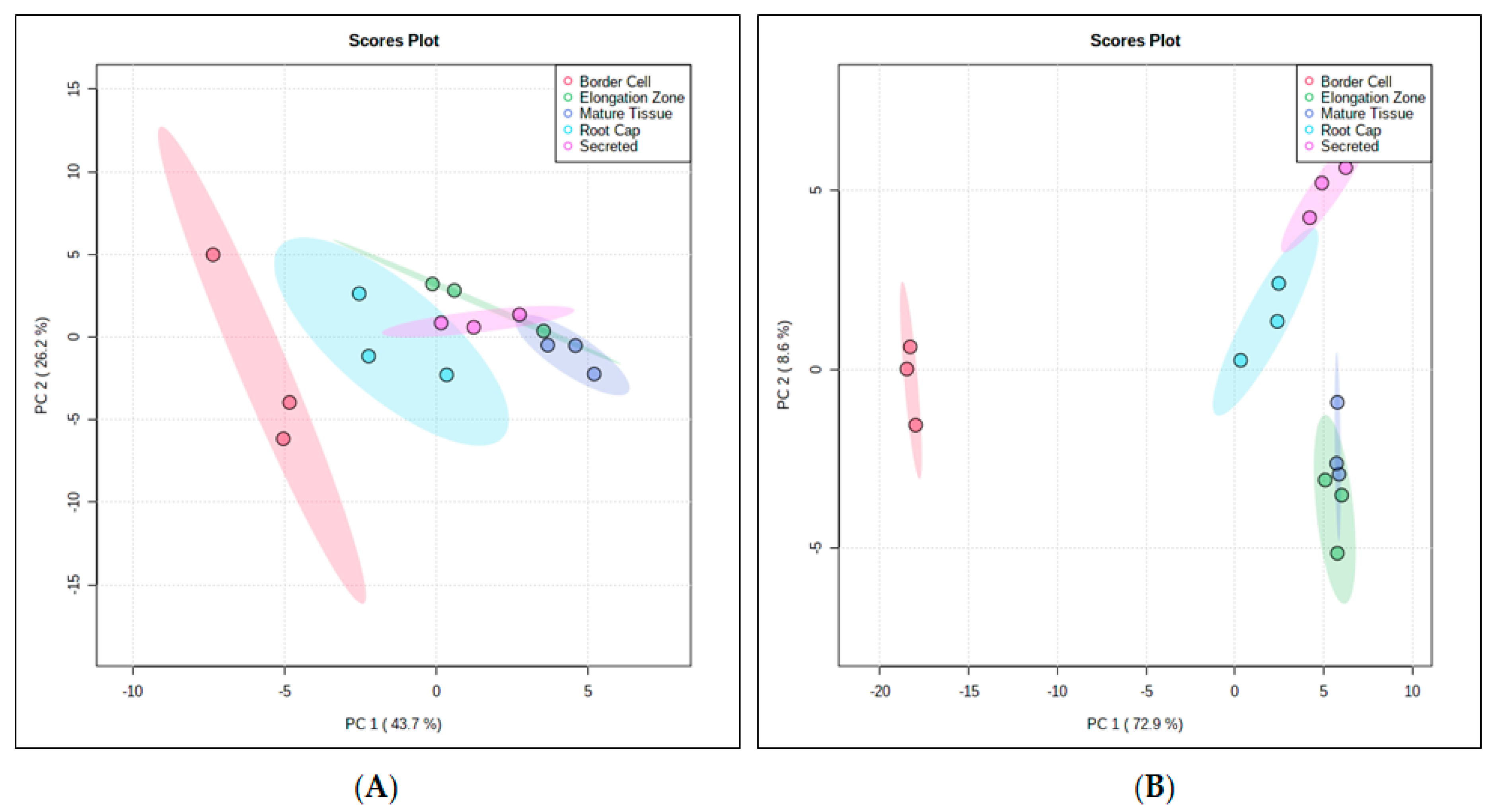{kind=link}
1. Introduction
2. Results
2.1. Data Generation and Compilation in the M. truncatula Metabolite Atlas
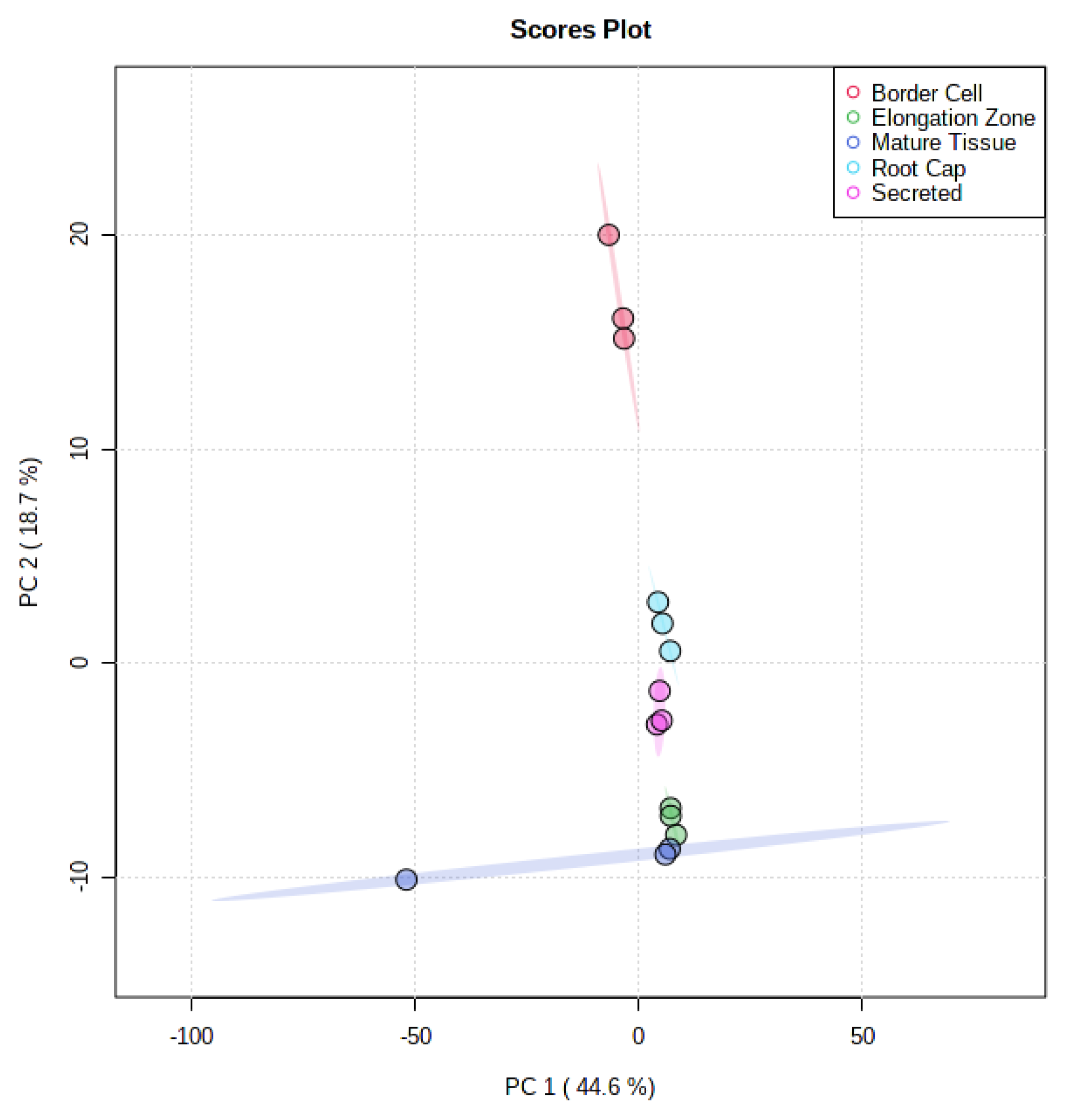
2.2. UHPLC-MS Analyses of Root Sections Show Differential Metabolite Accumulation
2.3. GC-MS Analyses of Root Sections Show Differential Accumulation of Metabolites
3. Discussion
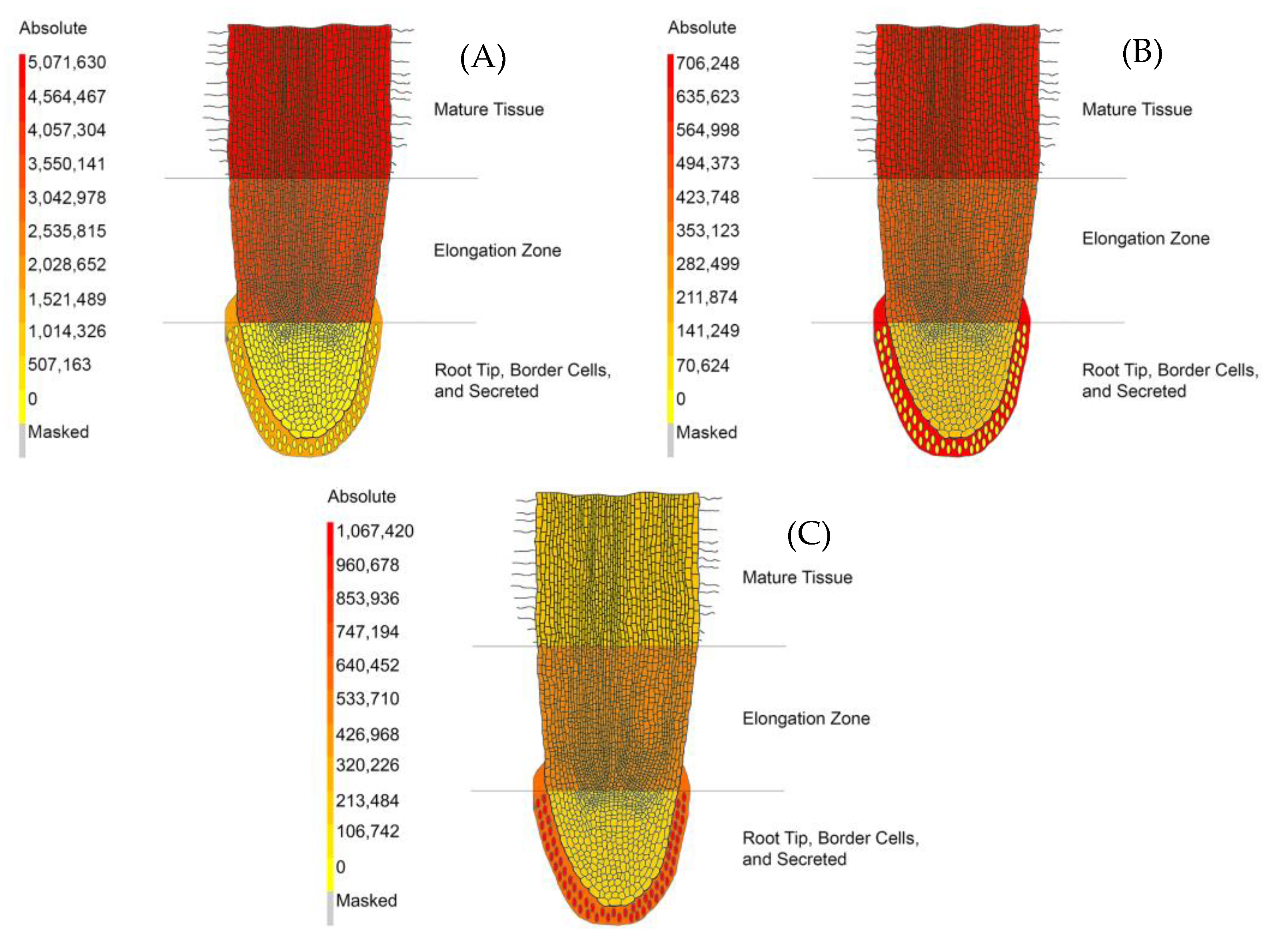
3.1. Visualization of Metabolite Spatial Localization Is Improved Using the Medicago truncatula Metabolite Atlas
3.2. UHPLC-MS and GC-MS Analyses Reveal Spatial Localization of Potentially Significant Metabolites
4. Materials and Methods
4.1. Tissue Collection
4.2. UHPLC-ESI-QTOF-MS
4.3. GC-Q-MS
4.4. Metabolite Atlas Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gunawardena, U.; Hawes, M.C. Tissue Specific Localization of Root Infection by Fungal Pathogens: Role of Root Border Cells. Mol. Plant Microbe Interact. 2002, 15, 1128–1136. [Google Scholar] [CrossRef]
- Gunawardena, U.; Rodriguez, M.; Straney, D.; Romeo, J.T.; VanEtten, H.D.; Hawes, M.C. Tissue-Specific Localization of Pea Root Infection by Nectria Haematococca. Mechanisms and Consequences. Plant Physiol. 2005, 137, 1363–1374. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.; Allen, C.; Turgeon, B.G.; Curlango-Rivera, G.; Tran, T.M.; Huskey, D.A.; Xiong, Z. Root Border Cells and Their Role in Plant Defense. Annu. Rev. Phytopathol. 2016, 54, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.S.; Bedair, M.F.; Urbanczyk-Wochniak, E.; Huhman, D.V.; Yang, D.S.; Allen, S.N.; Li, W.; Tang, Y.; Sumner, L.W. Integrated Metabolomics and Transcriptomics Reveal Enhanced Specialized Metabolism in Medicago Truncatula Root Border Cells. Plant Physiol. 2015, 167, 1699–1716. [Google Scholar] [CrossRef] [PubMed]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root Exudates Regulate Soil Fungal Community Composition and Diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef]
- Driouich, A.; Durand, C.; Vicré-Gibouin, M. Formation and Separation of Root Border Cells. Trends Plant Sci. 2007, 12, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; Vanetten, H.D.; Tsaprailis, G.; Hawes, M.C. Extracellular Proteins in Pea Root Tip and Border Cell Exudates. Plant Physiol. 2007, 143, 773–783. [Google Scholar] [CrossRef]
- Weiller, F.; Moore, J.P.; Young, P.; Driouich, A.; Vivier, M.A. The Brassicaceae Species Heliophila Coronopifolia Produces Root Border-like Cells That Protect the Root Tip and Secrete Defensin Peptides. Ann. Bot. 2017, 119, 803–813. [Google Scholar]
- Hawes, M.C.; Lin, H.J. Correlation of Pectolytic Enzyme Activity with the Programmed Release of Cells from Root Caps of Pea (Pisum Sativum). Plant Physiol. 1990, 94, 1855–1859. [Google Scholar] [CrossRef]
- Stephenson, M.B.; Hawes, M.C. Correlation of Pectin Methylesterase Activity in Root Caps of Pea with Root Border Cell Separation. Plant Physiol. 1994, 106, 739–745. [Google Scholar] [CrossRef]
- Wen, F.; Zhu, Y.; Hawes, M.C. Effect of Pectin Methylesterase Gene Expression on Pea Root Development. Plant Cell 1999, 11, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.C.; Pueppke, S.G. Sloughed Peripheral Root Cap Cells: Yield from Different Species and Callus Formation from Single Cells. Am. J. Bot. 1986, 73, 1466. [Google Scholar] [CrossRef]
- Huskey, D.A.; Curlango-Rivera, G.; Root, R.A.; Wen, F.; Amistadi, M.K.; Chorover, J.; Hawes, M.C. Trapping of Lead (Pb) by Corn and Pea Root Border Cells. Plant Soil 2018, 430, 205–217. [Google Scholar] [CrossRef]
- Huskey, D.A.; Curlango-Rivera, G.; Hawes, M.C. Use of Rhodizonic Acid for Rapid Detection of Root Border Cell Trapping of Lead and Reversal of Trapping with DNase. Appl. Plant Sci. 2019, 7, e01240. [Google Scholar] [CrossRef]
- Wang, P.; Chen, X.; Goldbeck, C.; Chung, E.; Kang, B.H. A Distinct Class of Vesicles Derived from the Trans-Golgi Mediates Secretion of Xylogalacturonan in the Root Border Cell. Plant J. 2017, 92, 596–610. [Google Scholar] [CrossRef]
- Knee, E.M.; Gong, F.C.; Gao, M.; Teplitski, M.; Jones, A.R.; Foxworthy, A.; Mort, A.J.; Bauer, W.D. Root Mucilage from Pea and Its Utilization by Rhizosphere Bacteria as a Sole Carbon Source. Mol. Plant Microbe Interact. 2001, 14, 775–784. [Google Scholar] [CrossRef]
- Wen, F.; White, G.J.; Vanetten, H.D.; Xiong, Z.; Hawes, M.C. Extracellular DNA is Required for Root Tip Resistance to Fungal Infection. Plant Physiol. 2009, 151, 820–829. [Google Scholar] [CrossRef]
- Park, H.J.; Wang, W.; Curlango-Rivera, G.; Xiong, Z.; Lin, Z.; Huskey, D.A.; Hawes, M.C.; Vanetten, H.D.; Turgeon, B.G. A Dnase from a Fungal Phytopathogen is a Virulence Factor Likely Deployed as Counter Defense against Host-Secreted Extracellular Dna. MBio 2019, 10. [Google Scholar] [CrossRef]
- Wen, F.; Curlango-Rivera, G.; Huskey, D.A.; Xiong, Z.; Hawes, M.C. Visualization of Extracellular DNA Released during Border Cell Separation from the Root Cap. Am. J. Bot. 2017, 104, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.C.; Curlango-Rivera, G.; Wen, F.; White, G.J.; VanEtten, H.D.; Xiong, Z. Extracellular DNA: The Tip of Root Defenses? Plant Sci. 2011, 180, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; MacIntyre, A.; Hawes, M.; Allen, C. Escaping Underground Nets: Extracellular DNases Degrade Plant Extracellular Traps and Contribute to Virulence of the Plant Pathogenic Bacterium Ralstonia Solanacearum. PLoS Pathog. 2016, 12, e1005686. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Brigham, L.A.; Woo, H.H.; Nicoll, S.M.; Hawes, M.C. Differential Expression of Proteins and MRNAs from Border Cells and Root Tips of Pea. Plant Physiol. 1995, 109, 457–463. [Google Scholar] [CrossRef]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” Browser for Exploring and Analyzing Large-Scale Biological Data Sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.C.; Bengough, G.; Cassab, G.; Ponce, G. Root Caps and Rhizosphere. J. Plant Growth Regul. 2002, 21, 352–367. [Google Scholar] [CrossRef]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H.; et al. Potential of Metabolomics as a Functional Genomics Tool. Trends Plant Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed Minimum Reporting Standards for Chemical Analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef]
- Yoo, H.; Widhalm, J.R.; Qian, Y.; Maeda, H.; Cooper, B.R.; Jannasch, A.S.; Gonda, I.; Lewinsohn, E.; Rhodes, D.; Dudareva, N. An Alternative Pathway Contributes to Phenylalanine Biosynthesis in Plants via a Cytosolic Tyrosine:Phenylpyruvate Aminotransferase. Nat. Commun. 2013, 4, 2833. [Google Scholar] [CrossRef]
- Qian, Y.; Lynch, J.H.; Guo, L.; Rhodes, D.; Morgan, J.A.; Dudareva, N. Completion of the Cytosolic Post-Chorismate Phenylalanine Biosynthetic Pathway in Plants. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef]
- Jassbi, A.R.; Zamanizadehnajari, S.; Baldwin, I.T. Phytotoxic Volatiles in the Roots and Shoots of Artemisia Tridentata as Detected by Headspace Solid-Phase Microextraction and Gas Chromatographic-Mass Spectrometry Analysis. J. Chem. Ecol. 2010, 36, 1398–1407. [Google Scholar] [CrossRef]
- Ninkovic, V.; Rensing, M.; Dahlin, I.; Markovic, D. Who is My Neighbor? Volatile Cues in Plant Interactions. Plant Signal. Behav. 2019, 14, 1634993. [Google Scholar] [CrossRef]
- Webb, B.A.; Compton, K.K.; del Campo, J.S.M.; Taylor, D.; Sobrado, P.; Scharf, B.E. Sinorhizobium Meliloti Chemotaxis to Multiple Amino Acids is Mediated by the Chemoreceptor Mcp U. Mol. Plant Microbe Interact. 2017, 30, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.N.V.; Signer, E.R. Metabolism of Tryptophan and Tryptophan Analogs by Rhizobium Meliloti. Plant Physiol. 1990, 92, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Gage, D.J. Infection and Invasion of Roots by Symbiotic, Nitrogen-Fixing Rhizobia during Nodulation of Temperate Legumes. Microbiol. Mol. Biol. Rev. 2004, 68, 280–300. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, C.A.; Hartwig, U.A.; Joseph, C.M.; Phillips, D.A. A Chalcone and Two Related Flavonoids Released from Alfalfa Roots Induce Nod Genes of Rhizobium Meliloti. Plant Physiol. 1989, 91, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Redmond, J.W.; Batley, M.; Djordjevic, M.A.; Innes, R.W.; Kuempel, P.L.; Rolfe, B.G. Flavones Induce Expression of Nodulation Genes in Rhizobium. Nature 1986, 323, 632–635. [Google Scholar] [CrossRef]
- Rudrappa, T.; Czymmek, K.J.; Paré, P.W.; Bais, H.P. Root-Secreted Malic Acid Recruits Beneficial Soil Bacteria. Plant Physiol. 2008, 148, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F. Role of Organic Acids in Detoxification of Aluminum in Higher Plants. Plant Cell Physiol. 2000, 41, 383–390. [Google Scholar] [CrossRef]
- Jones, D.L. Organic Acids in the Rhizosphere—A Critical Review. Plant Soil 1998, 205, 25–44. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef]
- Gherardi, M.J.; Rengel, Z. The Effect of Manganese Supply on Exudation of Carboxylates by Roots of Lucerne (Medicago Sativa). Plant Soil 2004, 260, 271–282. [Google Scholar] [CrossRef]
- Marquis, D.A. Seed Storage and Germination Under Northern Hardwood Forests. Can. J. For. Res. 1975, 5, 478–484. [Google Scholar] [CrossRef]
- Lei, Z.; Jing, L.; Qiu, F.; Zhang, H.; Huhman, D.; Zhou, Z.; Sumner, L.W. Construction of an Ultrahigh Pressure Liquid Chromatography-Tandem Mass Spectral Library of Plant Natural Products and Comparative Spectral Analyses. Anal. Chem. 2015, 87, 7373–7381. [Google Scholar] [CrossRef] [PubMed]
- Broeckling, C.D.; Huhman, D.V.; Farag, M.A.; Smith, J.T.; May, G.D.; Mendes, P.; Dixon, R.A.; Sumner, L.W. Metabolic Profiling of Medicago Truncatula Cell Cultures Reveals the Effects of Biotic and Abiotic Elicitors on Metabolism. J. Exp. Bot. 2005, 56, 323–336. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kranawetter, C.; Zeng, S.; Joshi, T.; Sumner, L.W. A Medicago truncatula Metabolite Atlas Enables the Visualization of Differential Accumulation of Metabolites in Root Tissues. Metabolites 2021, 11, 238. https://doi.org/10.3390/metabo11040238
Kranawetter C, Zeng S, Joshi T, Sumner LW. A Medicago truncatula Metabolite Atlas Enables the Visualization of Differential Accumulation of Metabolites in Root Tissues. Metabolites. 2021; 11(4):238. https://doi.org/10.3390/metabo11040238
Chicago/Turabian StyleKranawetter, Clayton, Shuai Zeng, Trupti Joshi, and Lloyd W. Sumner. 2021. "A Medicago truncatula Metabolite Atlas Enables the Visualization of Differential Accumulation of Metabolites in Root Tissues" Metabolites 11, no. 4: 238. https://doi.org/10.3390/metabo11040238
APA StyleKranawetter, C., Zeng, S., Joshi, T., & Sumner, L. W. (2021). A Medicago truncatula Metabolite Atlas Enables the Visualization of Differential Accumulation of Metabolites in Root Tissues. Metabolites, 11(4), 238. https://doi.org/10.3390/metabo11040238