
Metabolite Profile of Treatment-Naive Metabolic Syndrome Subjects in Relation to Cardiovascular Disease Risk

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Baseline Characteristics
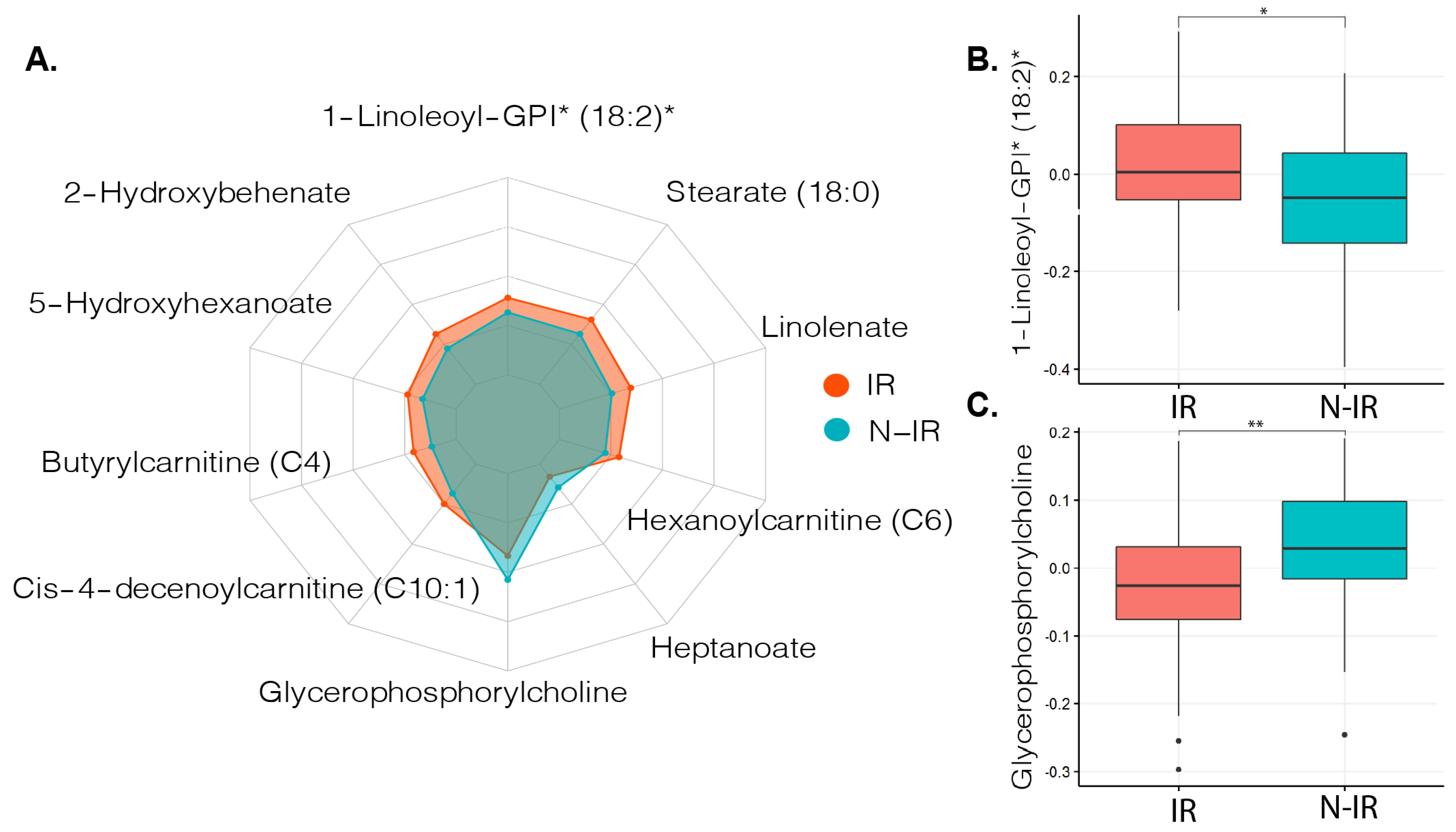
2.2. Metabolites in Relation to Peripheral Insulin Resistance
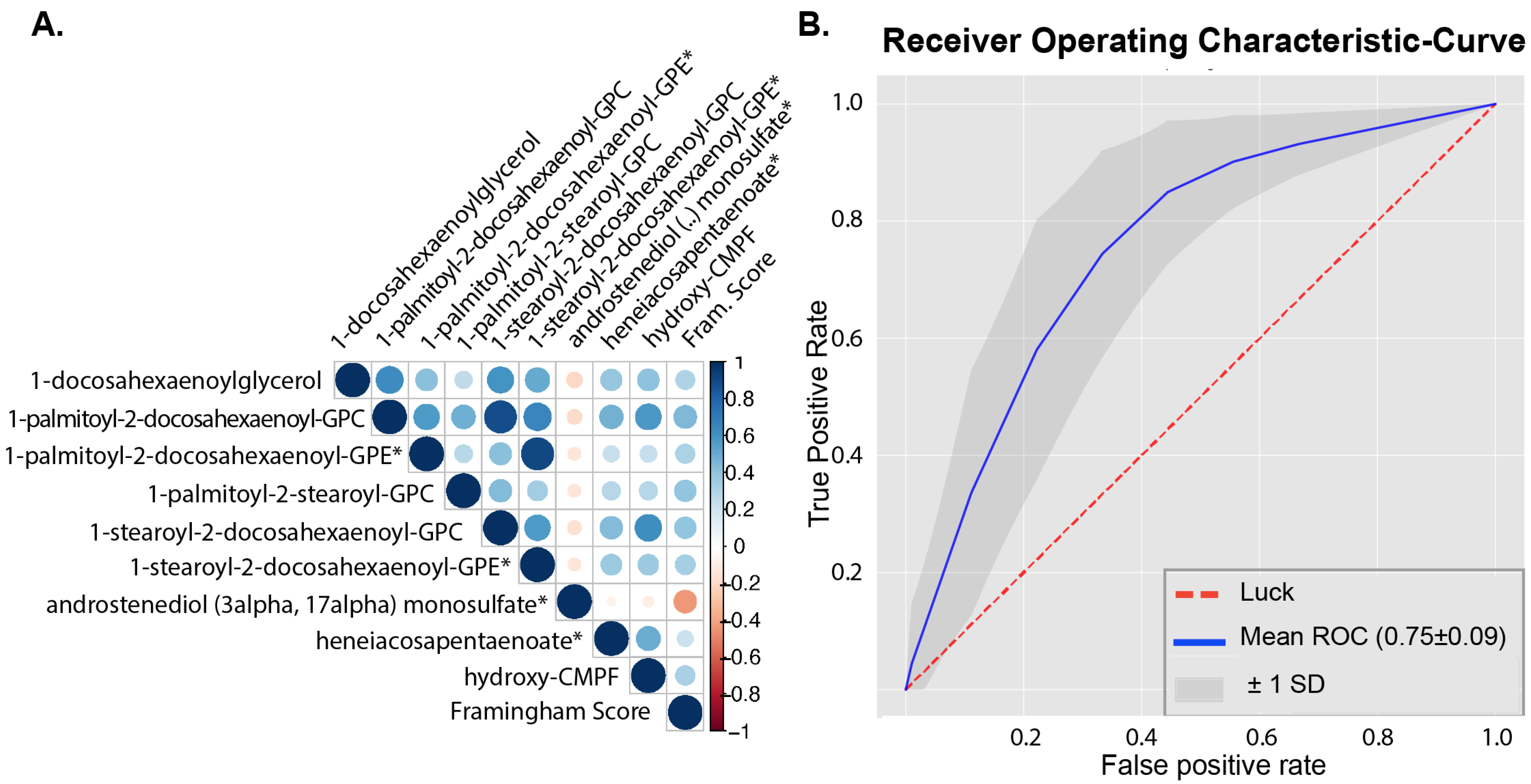
2.3. Metabolites in Relation to Cardiovascular Disease Risk
2.4. Metabolites in Relation to Lipolysis
3. Discussion
4. Materials and Methods
4.1. Study Design and Population
4.2. Clinical Parameters and Framingham Score
4.3. Metabolite Analysis
4.4. Two-Step Hyperinsulinemic Euglycemic Clamp and Lipolysis
4.5. Statistical Analysis and Machine Learning Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Apovian, C.M. Obesity: Definition, comorbidities, causes, and burden. Am. J. Manag. Care 2016, 22, s176–s185. [Google Scholar] [PubMed]
- Prospective Studies Collaboration. Body-mass index and cause-specific mortality in 900 000 adults: Collaborative analyses of 57 prospective studies. Lancet 2009, 373, 1083–1096. [Google Scholar] [CrossRef]
- Despres, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Ford, E.S.; Li, C.; Sattar, N. Metabolic syndrome and incident diabetes: Current state of the evidence. Diabetes Care 2008, 31, 1898–1904. [Google Scholar] [CrossRef]
- Gami, A.S.; Witt, B.J.; Howard, D.E.; Erwin, P.J.; Gami, L.A.; Somers, V.K.; Montori, V.M. Metabolic syndrome and risk of incident cardiovascular events and death: A systematic review and meta-analysis of longitudinal studies. J. Am. Coll. Cardiol. 2007, 49, 403–414. [Google Scholar] [CrossRef]
- Hamaguchi, M.; Kojima, T.; Takeda, N.; Nakagawa, T.; Taniguchi, H.; Fujii, K.; Omatsu, T.; Nakajima, T.; Sarui, H.; Shimazaki, M.; et al. The metabolic syndrome as a predictor of nonalcoholic fatty liver disease. Ann. Intern. Med. 2005, 143, 722–728. [Google Scholar] [CrossRef]
- Kurella, M.; Lo, J.C.; Chertow, G.M. Metabolic syndrome and the risk for chronic kidney disease among nondiabetic adults. J. Am. Soc. Nephrol. 2005, 16, 2134–2140. [Google Scholar] [CrossRef]
- Koopen, A.M.; de Clercq, N.C.; Warmbrunn, M.V.; Herrema, H.; Davids, M.; de Groot, P.F.; Kootte, R.S.; Bouter, K.E.C.; Nieuwdorp, M.; Groen, A.K.; et al. Plasma Metabolites Related to Peripheral and Hepatic Insulin Sensitivity Are Not Directly Linked to Gut Microbiota Composition. Nutrients 2020, 12, 2308. [Google Scholar] [CrossRef]
- Hartstra, A.V.; de Groot, P.F.; Mendes Bastos, D.; Levin, E.; Serlie, M.J.; Soeters, M.R.; Pekmez, C.T.; Dragsted, L.O.; Ackermans, M.T.; Groen, A.K.; et al. Correlation of plasma metabolites with glucose and lipid fluxes in human insulin resistance. Obes. Sci. Pract. 2020, 6, 340–349. [Google Scholar] [CrossRef]
- Guasch-Ferre, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Roberts, C.K.; Hevener, A.L.; Barnard, R.J. Metabolic syndrome and insulin resistance: Underlying causes and modification by exercise training. Compr. Physiol. 2013, 3, 1–58. [Google Scholar] [CrossRef]
- Pyle, L.; Bergman, B.C.; Nadeau, K.J.; Cree-Green, M. Modeling changes in glucose and glycerol rates of appearance when true basal rates of appearance cannot be readily determined. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E323–E331. [Google Scholar] [CrossRef] [PubMed]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuniga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Tobin, J.D.; Andres, R. Glucose clamp technique: A method for quantifying insulin secretion and resistance. Am. J. Physiol. 1979, 237, E214–E223. [Google Scholar] [CrossRef]
- Tam, C.S.; Xie, W.; Johnson, W.D.; Cefalu, W.T.; Redman, L.M.; Ravussin, E. Defining insulin resistance from hyperinsulinemic-euglycemic clamps. Diabetes Care 2012, 35, 1605–1610. [Google Scholar] [CrossRef]
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 38, 2459–2472. [Google Scholar] [CrossRef]
- Wurtz, P.; Havulinna, A.S.; Soininen, P.; Tynkkynen, T.; Prieto-Merino, D.; Tillin, T.; Ghorbani, A.; Artati, A.; Wang, Q.; Tiainen, M.; et al. Metabolite profiling and cardiovascular event risk: A prospective study of 3 population-based cohorts. Circulation 2015, 131, 774–785. [Google Scholar] [CrossRef]
- Nemet, I.; Saha, P.P.; Gupta, N.; Zhu, W.; Romano, K.A.; Skye, S.M.; Cajka, T.; Mohan, M.L.; Li, L.; Wu, Y.; et al. A Cardiovascular Disease-Linked Gut Microbial Metabolite Acts via Adrenergic Receptors. Cell 2020, 180, 862–877.e22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, K.W.; Gilijamse, P.W.; Koopman, K.E.; de Weijer, B.A.; Brands, M.; Kootte, R.S.; Romijn, J.A.; Ackermans, M.T.; Nieuwdorp, M.; Soeters, M.R.; et al. Insulin resistance in obesity can be reliably identified from fasting plasma insulin. Int. J. Obes. 2015, 39, 1703–1709. [Google Scholar] [CrossRef]
- Ke, C.; Zhu, X.; Zhang, Y.; Shen, Y. Metabolomic characterization of hypertension and dyslipidemia. Metabolomics 2018, 14, 117. [Google Scholar] [CrossRef]
- Meikle, P.J.; Summers, S.A. Sphingolipids and phospholipids in insulin resistance and related metabolic disorders. Nat. Rev. Endocrinol. 2017, 13, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Weir, J.M.; Wong, G.; Barlow, C.K.; Greeve, M.A.; Kowalczyk, A.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Jowett, J.B.; Shaw, J.; et al. Plasma lipid profiling in a large population-based cohort. J. Lipid Res. 2013, 54, 2898–2908. [Google Scholar] [CrossRef]
- Arendt, B.M.; Ma, D.W.; Simons, B.; Noureldin, S.A.; Therapondos, G.; Guindi, M.; Sherman, M.; Allard, J.P. Nonalcoholic fatty liver disease is associated with lower hepatic and erythrocyte ratios of phosphatidylcholine to phosphatidylethanolamine. Appl. Physiol. Nutr. Metab. 2013, 38, 334–340. [Google Scholar] [CrossRef]
- Meikle, P.J.; Wong, G.; Barlow, C.K.; Weir, J.M.; Greeve, M.A.; MacIntosh, G.L.; Almasy, L.; Comuzzie, A.G.; Mahaney, M.C.; Kowalczyk, A.; et al. Plasma lipid profiling shows similar associations with prediabetes and type 2 diabetes. PLoS ONE 2013, 8, e74341. [Google Scholar] [CrossRef] [PubMed]
- Warmbrunn, M.V.; Herrema, H.; Aron-Wisnewsky, J.; Soeters, M.R.; Van Raalte, D.H.; Nieuwdorp, M. Gut microbiota: A promising target against cardiometabolic diseases. Expert Rev. Endocrinol. Metab. 2020, 15, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Surowiec, I.; Noordam, R.; Bennett, K.; Beekman, M.; Slagboom, P.E.; Lundstedt, T.; van Heemst, D. Metabolomic and lipidomic assessment of the metabolic syndrome in Dutch middle-aged individuals reveals novel biological signatures separating health and disease. Metabolomics 2019, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Boden, G. Obesity, insulin resistance and free fatty acids. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 139–143. [Google Scholar] [CrossRef]
- Gao, C.L.; Zhu, C.; Zhao, Y.P.; Chen, X.H.; Ji, C.B.; Zhang, C.M.; Zhu, J.G.; Xia, Z.K.; Tong, M.L.; Guo, X.R. Mitochondrial dysfunction is induced by high levels of glucose and free fatty acids in 3T3-L1 adipocytes. Mol. Cell Endocrinol. 2010, 320, 25–33. [Google Scholar] [CrossRef]
- Tuboly, E.; Gaspar, R.; Ibor, M.O.; Gomori, K.; Kiss, B.; Strifler, G.; Hartmann, P.; Ferdinandy, P.; Bartekova, M.; Boros, M.; et al. L-Alpha-glycerylphosphorylcholine can be cytoprotective or cytotoxic in neonatal rat cardiac myocytes: A double-edged sword phenomenon. Mol. Cell Biochem. 2019, 460, 195–203. [Google Scholar] [CrossRef]
- Strifler, G.; Tuboly, E.; Gorbe, A.; Boros, M.; Pecz, D.; Hartmann, P. Targeting Mitochondrial Dysfunction with L-Alpha Glycerylphosphorylcholine. PLoS ONE 2016, 11, e0166682. [Google Scholar] [CrossRef]
- Hurrle, S.; Hsu, W.H. The etiology of oxidative stress in insulin resistance. Biomed. J. 2017, 40, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.N.; Hovik, K.; Bremer, J.; Holm, K.H.; Myhren, F.; Borretzen, B. Heneicosapentaenoate (21:5n-3): Its incorporation into lipids and its effects on arachidonic acid and eicosanoid synthesis. Lipids 1997, 32, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Mihalik, S.J.; Goodpaster, B.H.; Kelley, D.E.; Chace, D.H.; Vockley, J.; Toledo, F.G.; DeLany, J.P. Increased levels of plasma acylcarnitines in obesity and type 2 diabetes and identification of a marker of glucolipotoxicity. Obesity 2010, 18, 1695–1700. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Neufer, P.D. Lipid-induced mitochondrial stress and insulin action in muscle. Cell Metab. 2012, 15, 595–605. [Google Scholar] [CrossRef]
- Aguer, C.; McCoin, C.S.; Knotts, T.A.; Thrush, A.B.; Ono-Moore, K.; McPherson, R.; Dent, R.; Hwang, D.H.; Adams, S.H.; Harper, M.E. Acylcarnitines: Potential implications for skeletal muscle insulin resistance. FASEB J. 2015, 29, 336–345. [Google Scholar] [CrossRef]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [PubMed]
- National Cholesterol Education Program (US). Expert Panel on Detection and Treatment of High Blood Cholesterol in Adults. Executive Summary of the Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). JAMA 2002, 285, 2486–2497. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojarvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, R.B., Sr.; Vasan, R.S.; Pencina, M.J.; Wolf, P.A.; Cobain, M.; Massaro, J.M.; Kannel, W.B. General cardiovascular risk profile for use in primary care: The Framingham Heart Study. Circulation 2008, 117, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; Molinaro, A.; Stahlman, M.; Khan, M.T.; Schmidt, C.; Manneras-Holm, L.; Wu, H.; Carreras, A.; Jeong, H.; Olofsson, L.E.; et al. Microbially Produced Imidazole Propionate Impairs Insulin Signaling through mTORC1. Cell 2018, 175, 947–961.e917. [Google Scholar] [CrossRef] [PubMed]
- Finegood, D.T.; Bergman, R.N.; Vranic, M. Estimation of endogenous glucose production during hyperinsulinemic-euglycemic glucose clamps. Comparison of unlabeled and labeled exogenous glucose infusates. Diabetes 1987, 36, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Steele, R. Influences of glucose loading and of injected insulin on hepatic glucose output. Ann. N. Y. Acad. Sci. 1959, 82, 420–430. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Overall | IR (Rd < 37.3) | N-IR (Rd ≥ 37.3) | p-Value | |
|---|---|---|---|---|
| n | 132 | 92 | 40 | |
| Age (years) | 53.83 (9.38) | 53.61 (10.31) | 54.35 (6.89) | 0.678 |
| BMI (kg/m2) | 33.91 [31.45, 37.05] | 34.50 [31.58, 38.69] | 33.40 [30.82, 35.00] | 0.024 |
| Weight (kg) | 115.40 [102.10, 124.62] | 117.70 [105.45, 130.32] | 108.45 [99.95, 118.17] | 0.003 |
| Syst (mmHg) | 143.61 (18.20) | 144.59 (18.63) | 140.72 (16.85) | 0.326 |
| Diast (mmHg) | 89.45 (11.11) | 90.58 (10.93) | 86.14 (11.15) | 0.063 |
| Gluc (mmol/L) | 5.72 (0.66) | 5.74 (0.70) | 5.68 (0.55) | 0.599 |
| Insulin (pmol/L) | 109.00 [70.75, 141.75] | 123.00 [93.00, 158.50] | 69.00 [54.75, 87.00] | <0.001 |
| Rd (μmol kg−1 min−1) | 31.37 [22.65, 40.02] | 27.00 [19.86, 33.01] | 48.10 [41.08, 55.45] | <0.001 |
| HbA1c (mmol/mol) | 39.00 [36.00, 41.00] | 39.00 [36.00, 42.00] | 38.50 [37.00, 40.75] | 0.58 |
| HOMA-IR | 3.70 [2.50, 5.16] | 4.29 [3.19, 5.46] | 2.55 [1.90, 3.05] | <0.001 |
| Total chol (mmol/L) | 5.00 [4.59, 5.89] | 5.09 [4.56, 5.85] | 4.90 [4.63, 6.02] | 0.831 |
| LDL (mmol/L) | 3.30 [2.70, 4.10] | 3.26 [2.70, 4.00] | 3.38 [2.66, 4.15] | 0.974 |
| HDL (mmol/L) | 1.08 [0.93, 1.23] | 1.04 [0.93, 1.21] | 1.10 [0.96, 1.33] | 0.268 |
| Trig (mmol/L) | 1.40 [1.12, 1.79] | 1.42 [1.16, 1.80] | 1.23 [1.10, 1.66] | 0.097 |
| ALAT (U/L) | 33.00 [26.00, 41.00] | 34.00 [27.00, 43.00] | 31.00 [22.50, 36.25] | 0.022 |
| CRP (mg/L) | 2.00 [1.30, 4.35] | 2.20 [1.37, 4.70] | 2.00 [1.05, 3.80] | 0.544 |
| Leuko (10E9/L) | 6.01 (1.41) | 6.02 (1.37) | 5.95 (1.57) | 0.845 |
| REE (kcal/day) | 1939.00 [1804.00, 2190.50] | 1952.00 [1806.00, 2246.70] | 1924.00 [1760.00, 2083.25] | 0.172 |
| Biochemical Annotation | Subpathway Annotation | Super-Pathway Annotation | Low Framingham (n = 42, Median [IQR])) | High Framingham (n = 69, median [IQR]) |
|---|---|---|---|---|
| Eicosapentaenoate (EPA) | Long-Chain Polyunsaturated Fatty Acid (n3 and n6) | Lipid | 0.85 [0.69, 1.20] | 1.19 [0.91, 1.65] |
| N-acetyltyrosine | Tyrosine Metabolism | Amino Acid | 0.93 [0.82, 1.17] | 1.22 [0.88, 1.57] |
| 1-stearoyl-2-docosahexaenoyl-GPE (18:0/22:6) * | Phosphatidylethanolamine (PE) | Lipid | 0.79 [0.60, 1.05] | 1.21 [0.77, 1.50] |
| Docosahexaenoylcholine | Fatty Acid Metabolism (Acyl Choline) | Lipid | 0.81 [0.62, 1.28] | 1.14 [0.89, 1.58] |
| Heneicosapentaenoate (21:5n3) | Long-Chain Polyunsaturated Fatty Acid (n3 and n6) | Lipid | 0.26 [0.26, 0.60] | 0.71 [0.26, 1.70] |
| Androstenediol (3alpha, 17alpha) monosulfate (2) | Androgenic Steroids | Lipid | 1.16 [0.91, 1.69] | 0.81 [0.65, 1.02] |
| 3-carboxy-4-methyl-5-propyl-2-furanpropanoate (CMPF) | Fatty Acid, Dicarboxylate | Lipid | 0.68 [0.26, 1.63] | 1.61 [0.62, 2.62] |
| 1-palmitoyl-2-docosahexaenoyl-GPE (16:0/22:6) * | Phosphatidylethanolamine (PE) | Lipid | 0.74 [0.58, 1.19] | 1.23 [0.78, 1.45] |
| 1-palmitoyl-2-stearoyl-GPC (16:0/18:0) | Phosphatidylcholine (PC) | Lipid | 0.96 [0.84, 1.02] | 1.04 [0.93, 1.12] |
| lanthionine | Methionine, Cysteine, SAM, and Taurine Metabolism | Amino Acid | 0.23 [0.23, 0.38] | 0.69 [0.23, 1.22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warmbrunn, M.V.; Koopen, A.M.; de Clercq, N.C.; de Groot, P.F.; Kootte, R.S.; Bouter, K.E.C.; ter Horst, K.W.; Hartstra, A.V.; Serlie, M.J.; Ackermans, M.T.; et al. Metabolite Profile of Treatment-Naive Metabolic Syndrome Subjects in Relation to Cardiovascular Disease Risk. Metabolites 2021, 11, 236. https://doi.org/10.3390/metabo11040236
Warmbrunn MV, Koopen AM, de Clercq NC, de Groot PF, Kootte RS, Bouter KEC, ter Horst KW, Hartstra AV, Serlie MJ, Ackermans MT, et al. Metabolite Profile of Treatment-Naive Metabolic Syndrome Subjects in Relation to Cardiovascular Disease Risk. Metabolites. 2021; 11(4):236. https://doi.org/10.3390/metabo11040236
Chicago/Turabian StyleWarmbrunn, Moritz V., Annefleur M. Koopen, Nicolien C. de Clercq, Pieter F. de Groot, Ruud S. Kootte, Kristien E. C. Bouter, Kasper W. ter Horst, Annick V. Hartstra, Mireille J. Serlie, Mariette T. Ackermans, and et al. 2021. "Metabolite Profile of Treatment-Naive Metabolic Syndrome Subjects in Relation to Cardiovascular Disease Risk" Metabolites 11, no. 4: 236. https://doi.org/10.3390/metabo11040236
APA StyleWarmbrunn, M. V., Koopen, A. M., de Clercq, N. C., de Groot, P. F., Kootte, R. S., Bouter, K. E. C., ter Horst, K. W., Hartstra, A. V., Serlie, M. J., Ackermans, M. T., Soeters, M. R., van Raalte, D. H., Davids, M., Nieuwdorp, M., & Groen, A. K. (2021). Metabolite Profile of Treatment-Naive Metabolic Syndrome Subjects in Relation to Cardiovascular Disease Risk. Metabolites, 11(4), 236. https://doi.org/10.3390/metabo11040236