
Developing a Yeast Platform Strain for an Enhanced Taxadiene Biosynthesis by CRISPR/Cas9

 , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
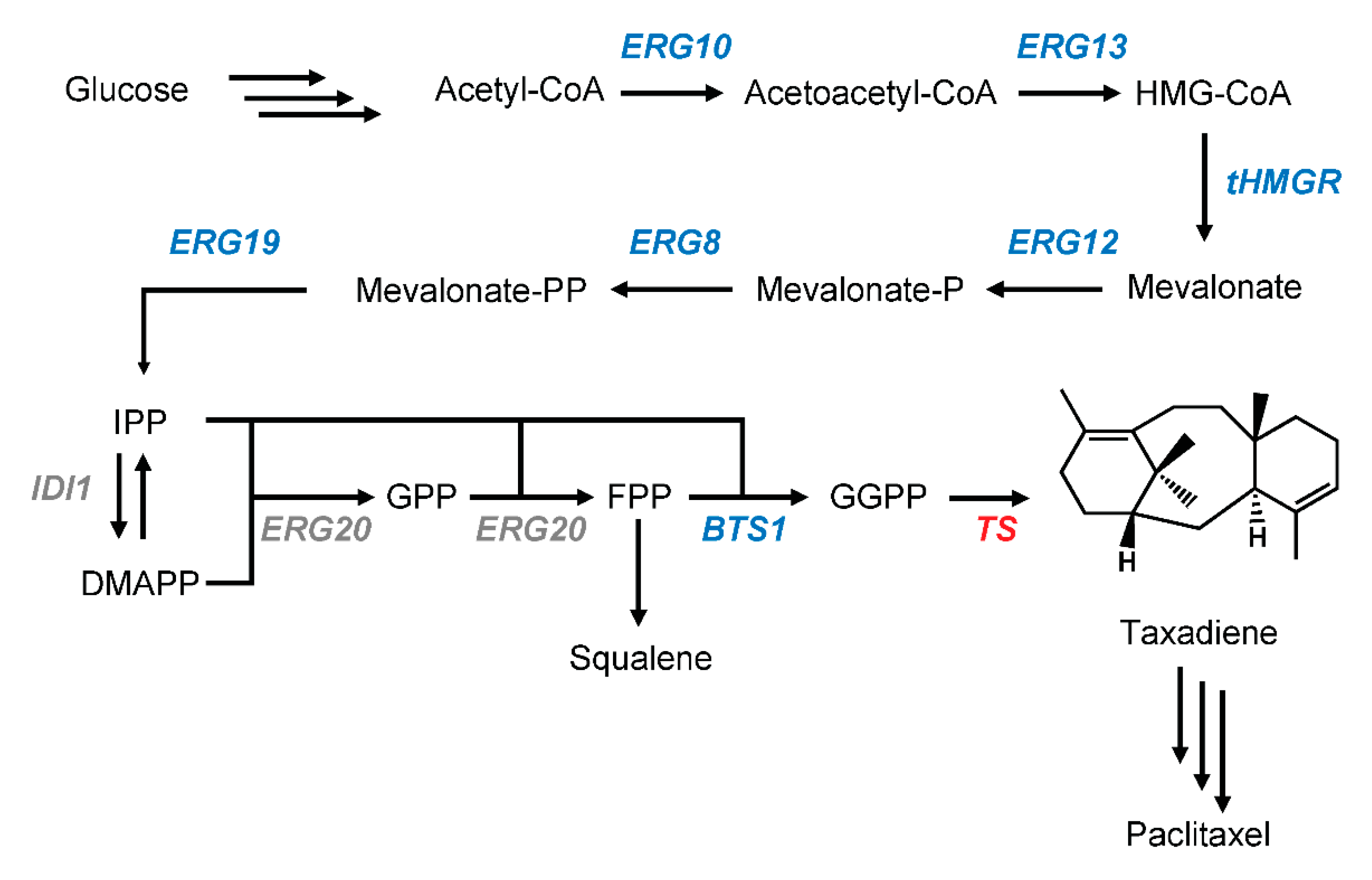
2.1. pIPP Construction to Increase the Flux towards Terpenoid Pathway
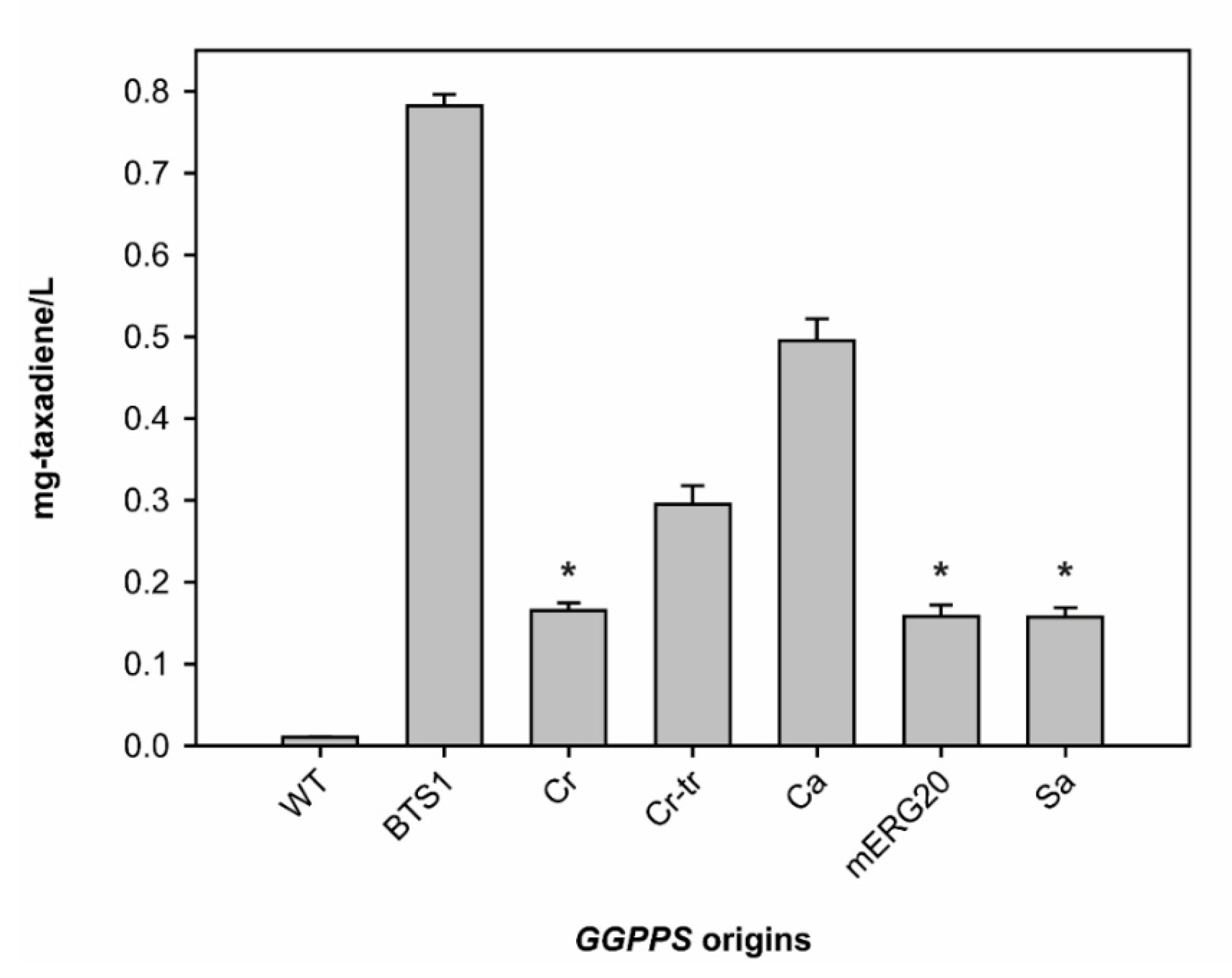
2.2. Various GGPPS Overexpression
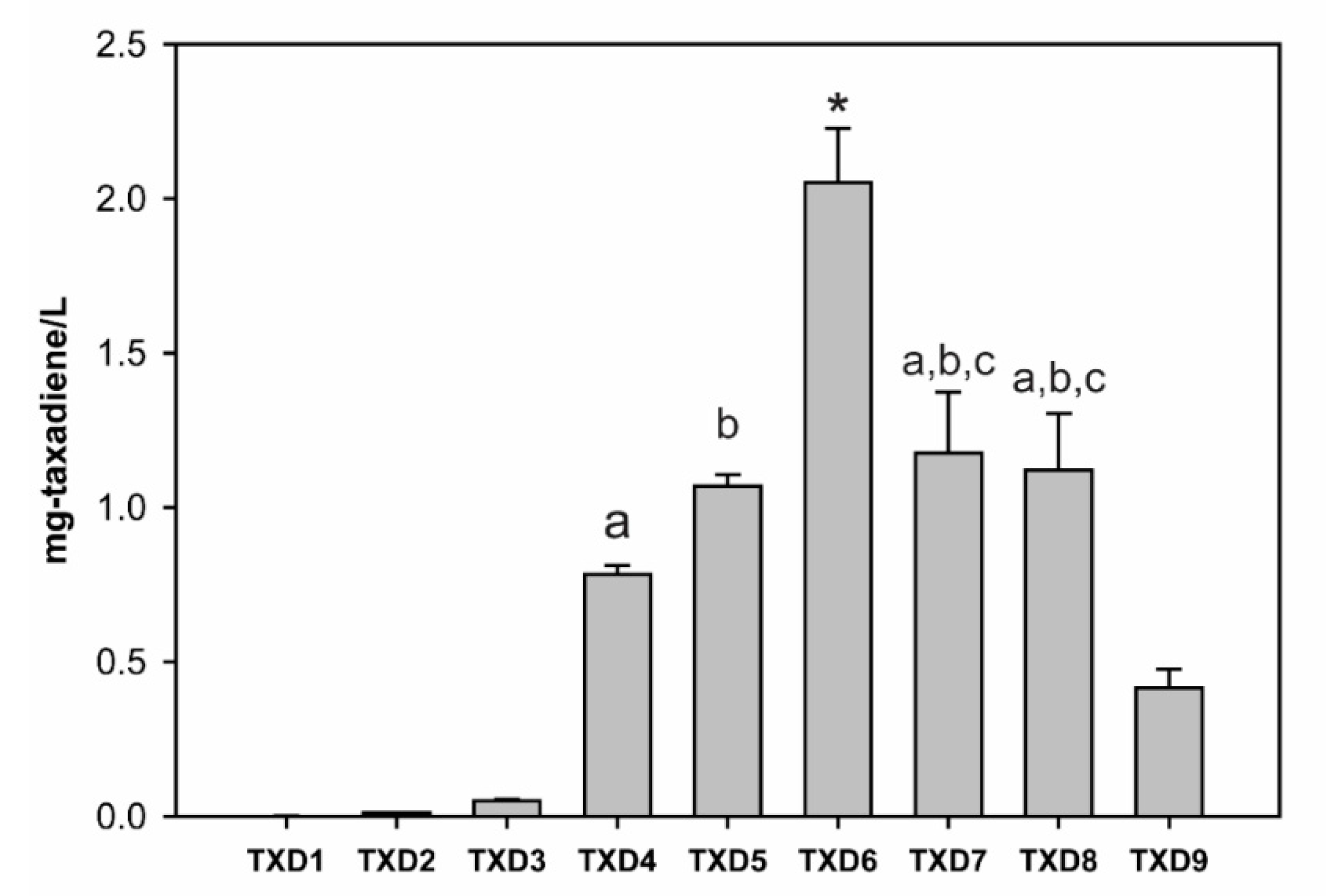
2.3. Optimization of Taxadiene Production
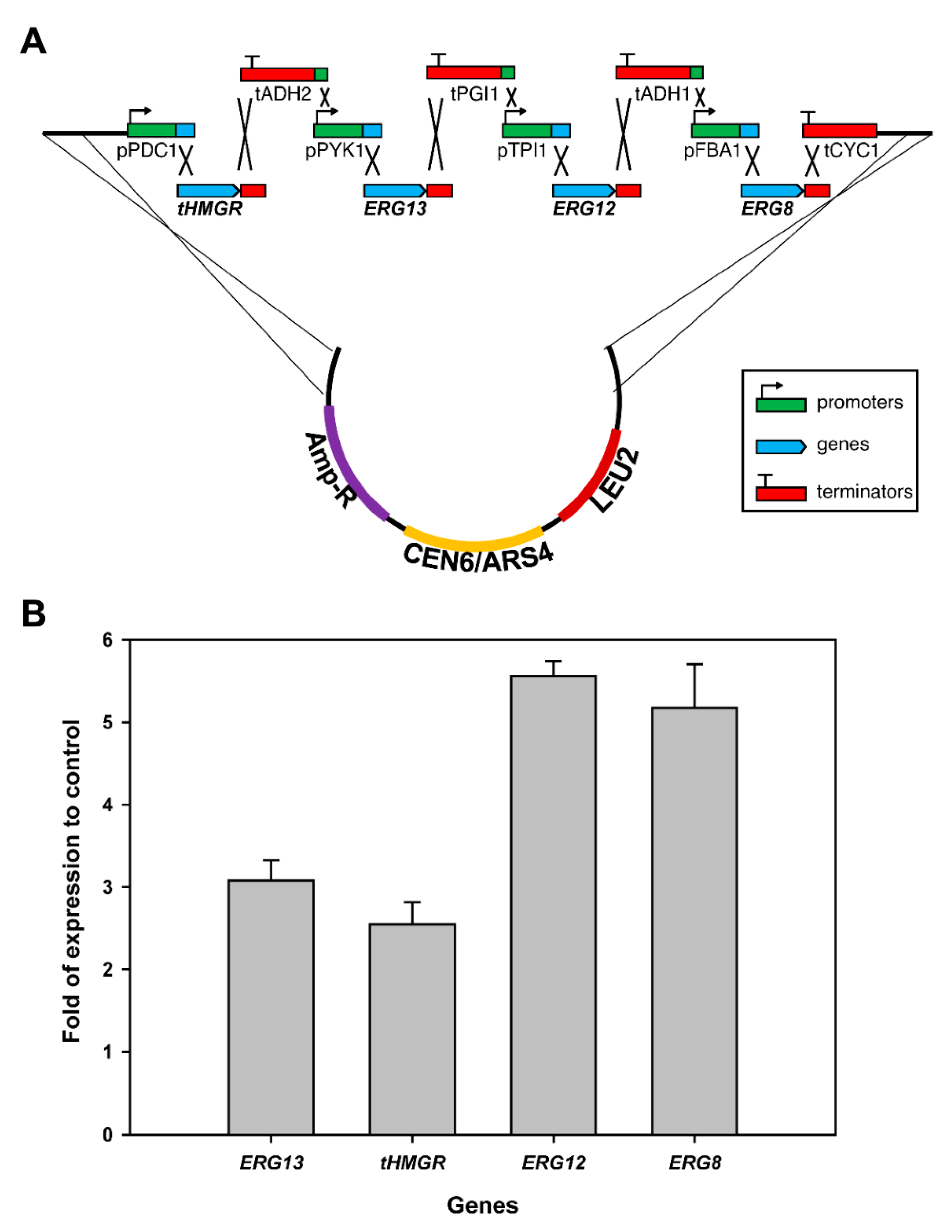
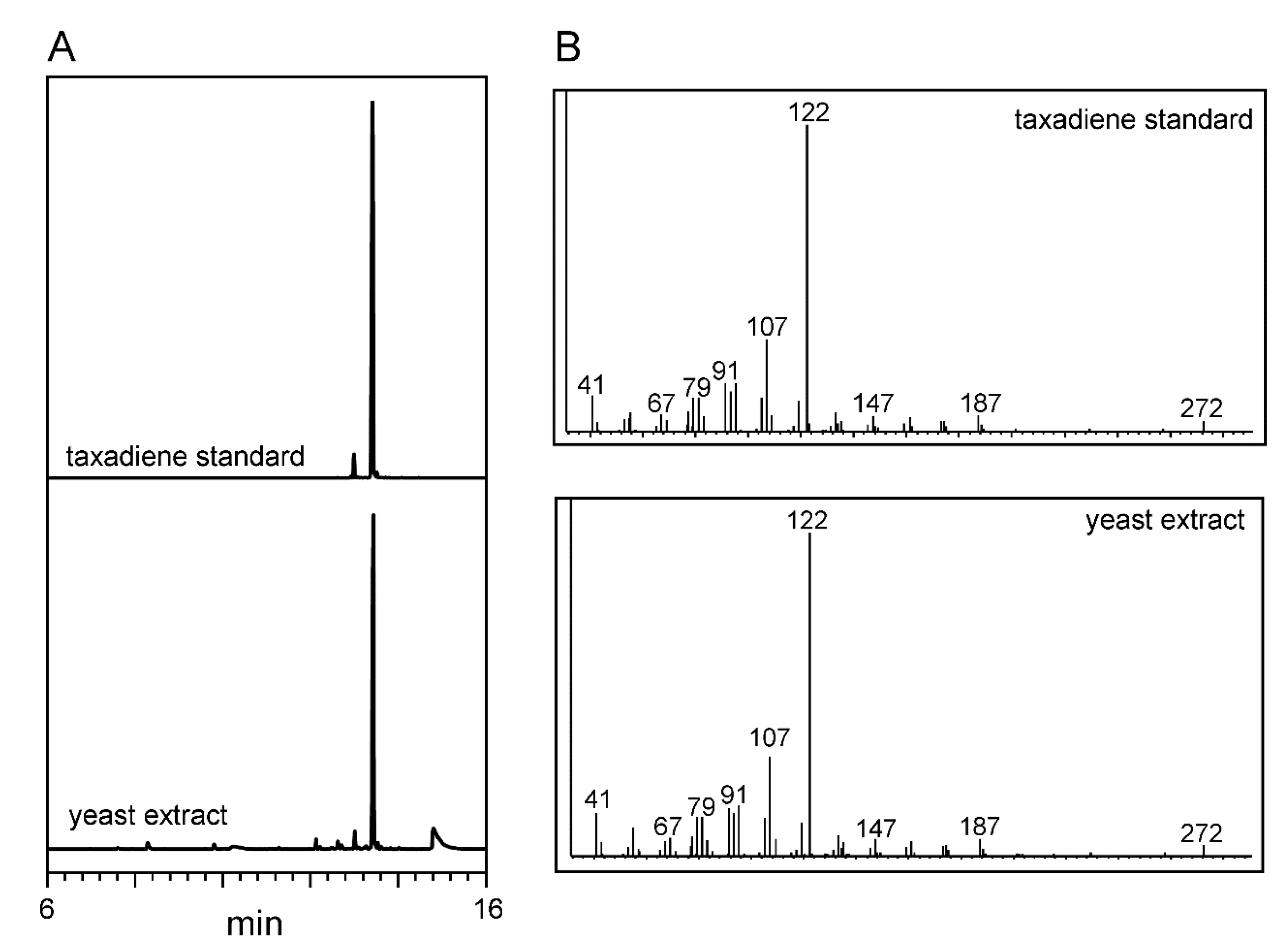
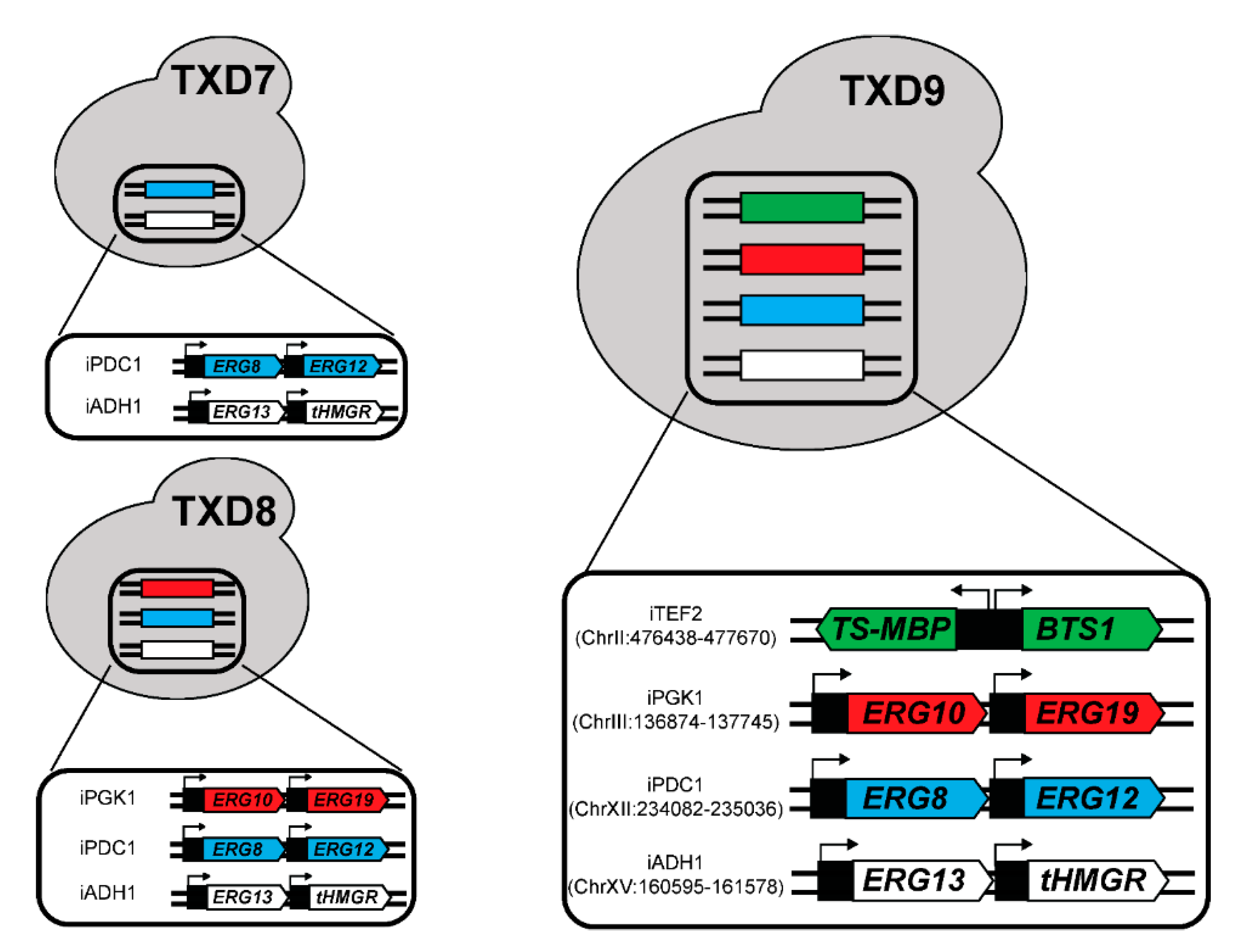
2.4. CRISPR-Mediated Gene Integration
3. Materials and Methods
3.1. Yeast and Bacterial Strains
3.2. Plasmid and Synthetic DNA Construction
3.3. RNA Isolation, cDNA Synthesis, and qPCR Analysis
3.4. Yeast Expression and Cultivation
3.5. Immunoblot
3.6. Yeast Metabolites Extraction and Analysis
3.7. Genes Integration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Khanna, C.; Rosenberg, M.; Vail, D.M. A Review of Paclitaxel and Novel Formulations Including Those Suitable for Use in Dogs. J. Vet. Intern. Med. 2015, 29, 1006–1012. [Google Scholar] [CrossRef]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Bernabeu, E.; Cagel, M.; Lagomarsino, E.; Moretton, M.; Chiappetta, D.A. Paclitaxel: What has been done and the challenges remain ahead. Int. J. Pharm. 2017, 526, 474–495. [Google Scholar] [CrossRef] [PubMed]
- Kampan, N.C.; Madondo, M.T.; McNally, O.M.; Quinn, M.; Plebanski, M. Paclitaxel and Its Evolving Role in the Management of Ovarian Cancer. Biomed. Res. Int. 2015, 2015, 413076. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, G.; Pfeifer, B.A. Current and Emerging Options for Taxol Production. In Biotechnology of Isoprenoids; Schrader, J., Bohlmann, J., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 405–425. [Google Scholar]
- Yuzuru, K.; Hugh, N.; Shigenobu, U.; Ravi Kumar, P.; Venkata Ramana Murthy, A.; Gopi Krishna, G.; Bheema Rao, P.; Phil, B. Two-Phase Synthesis of Taxol. J. Am. Chem. Soc. 2020, 142, 10526–10533. [Google Scholar]
- Wilson, S.A.; Roberts, S.C. Recent advances towards development and commercialization of plant cell culture processes for synthesis of biomolecules. Plant. Biotechnol. J. 2012, 10, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Cusido, R.M.; Onrubia, M.; Sabater-Jara, A.B.; Moyano, E.; Bonfill, M.; Goossens, A.; Angeles Pedreno, M.; Palazon, J. A rational approach to improving the biotechnological production of taxanes in plant cell cultures of Taxus spp. Biotechnol. Adv. 2014, 32, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.L.; English, D.P.; Roque, D.M.; Santin, A.D. Taxanes: Their Impact on Gynecologic Malignancy. Anticancer Drugs 2014, 25, 522–535. [Google Scholar] [CrossRef]
- Jennewein, S.; Wildung, M.R.; Chau, M.; Walker, K.; Croteau, R. Random sequencing of an induced Taxus cell cDNA library for identification of clones involved in Taxol biosynthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 9149–9154. [Google Scholar] [CrossRef]
- Wildung, M.R.; Croteau, R. A cDNA clone for taxadiene synthase, the diterpene cyclase that catalyzes the committed step of taxol biosynthesis. J. Biol. Chem. 1996, 271, 9201–9204. [Google Scholar] [CrossRef] [PubMed]
- Jennewein, S.; Croteau, R. Taxol: Biosynthesis, molecular genetics, and biotechnological applications. Appl. Microbiol. Biotechnol. 2001, 57, 13–19. [Google Scholar] [PubMed]
- Chau, M.; Croteau, R. Molecular cloning and characterization of a cytochrome P450 taxoid 2alpha-hydroxylase involved in Taxol biosynthesis. Arch. BioChem. Biophys. 2004, 427, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Chau, M.; Jennewein, S.; Walker, K.; Croteau, R. Taxol biosynthesis: Molecular cloning and characterization of a cytochrome P450 taxoid 7 beta-hydroxylase. Chem. Biol. 2004, 11, 663–672. [Google Scholar] [PubMed]
- Jennewein, S.; Rithner, C.D.; Williams, R.M.; Croteau, R.B. Taxol biosynthesis: Taxane 13 alpha-hydroxylase is a cytochrome P450-dependent monooxygenase. Proc. Natl. Acad. Sci. USA 2001, 98, 13595–13600. [Google Scholar] [CrossRef]
- Hefner, J.; Rubenstein, S.M.; Ketchum, R.E.; Gibson, D.M.; Williams, R.M.; Croteau, R. Cytochrome P450-catalyzed hydroxylation of taxa-4(5),11(12)-diene to taxa-4(20),11(12)-dien-5alpha-ol: The first oxygenation step in taxol biosynthesis. Chem. Biol. 1996, 3, 479–489. [Google Scholar] [CrossRef]
- Rontein, D.; Onillon, S.; Herbette, G.; Lesot, A.; Werck-Reichhart, D.; Sallaud, C.; Tissier, A. CYP725A4 from yew catalyzes complex structural rearrangement of taxa-4(5),11(12)-diene into the cyclic ether 5(12)-oxa-3(11)-cyclotaxane. J. Biol. Chem. 2008, 283, 6067–6075. [Google Scholar] [CrossRef]
- Edgar, S.; Zhou, K.; Qiao, K.; King, J.R.; Simpson, J.H.; Stephanopoulos, G. Mechanistic Insights into Taxadiene Epoxidation by Taxadiene-5alpha-Hydroxylase. ACS Chem. Biol. 2016, 11, 460–469. [Google Scholar] [CrossRef]
- Huang, Q.; Roessner, C.A.; Croteau, R.; Scott, A.I. Engineering Escherichia coli for the synthesis of taxadiene, a key intermediate in the biosynthesis of taxol. Bioorg. Med. Chem. 2001, 9, 2237–2242. [Google Scholar] [CrossRef]
- Ajikumar, P.K.; Xiao, W.H.; Tyo, K.E.; Wang, Y.; Simeon, F.; Leonard, E.; Mucha, O.; Phon, T.H.; Pfeifer, B.; Stephanopoulos, G. Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 2010, 330, 70–74. [Google Scholar] [CrossRef]
- Biggs, B.W.; Lim, C.G.; Sagliani, K.; Shankar, S.; Stephanopoulos, G.; De Mey, M.; Ajikumar, P.K. Overcoming heterologous protein interdependency to optimize P450-mediated Taxol precursor synthesis in Escherichia coli. Proc. Natl. Acad. Sci. USA 2016, 113, 3209–3214. [Google Scholar] [CrossRef]
- Dejong, J.M.; Liu, Y.; Bollon, A.P.; Long, R.M.; Jennewein, S.; Williams, D.; Croteau, R.B. Genetic engineering of taxol biosynthetic genes in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2006, 93, 212–224. [Google Scholar] [CrossRef]
- Engels, B.; Dahm, P.; Jennewein, S. Metabolic engineering of taxadiene biosynthesis in yeast as a first step towards Taxol (Paclitaxel) production. Metab. Eng. 2008, 10, 201–206. [Google Scholar] [CrossRef]
- Apel, A.R.; d’Espaux, L.; Wehrs, M.; Sachs, D.; Li, R.A.; Tong, G.J.; Garber, M.; Nnadi, O.; Zhuang, W.; Hillson, N.J.; et al. A Cas9-based toolkit to program gene expression in Saccharomyces cerevisiae. Nucleic Acids Res. 2017, 45, 496–508. [Google Scholar]
- Nowrouzi, B.; Li, R.A.; Walls, L.E.; d’Espaux, L.; Malci, K.; Liang, L.; Jonguitud-Borrego, N.; Lerma-Escalera, A.I.; Morones-Ramirez, J.R.; Keasling, J.D.; et al. Enhanced production of taxadiene in Saccharomyces cerevisiae. Microb. Cell Fact. 2020, 19, 200. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Proteau, P.; Poulter, D.; Ferro-Novick, S. BTS1 Encodes a Geranylgeranyl Diphosphate Synthase in Saccharomyces cerevisiae. J. Biol. Chem. 1995, 270, 21793–21799. [Google Scholar] [CrossRef]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Donald, K.A.; Hampton, R.Y.; Fritz, I.B. Effects of overproduction of the catalytic domain of 3-hydroxy-3-methylglutaryl coenzyme A reductase on squalene synthesis in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1997, 63, 3341–3344. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Sánchez, R.; Saavedra, E.; Rodríguez-Enríquez, S.; Olín-Sandoval, V. Metabolic Control Analysis: A Tool for Designing Strategies to Manipulate Metabolic Pathways. J. Biomed. Biotechnol. 2008, 2008, 597913. [Google Scholar] [CrossRef] [PubMed]
- Tanner, L.B.; Goglia, A.G.; Wei, M.H.; Sehgal, T.; Parsons, L.R.; Park, J.O.; White, E.; Toettcher, J.E.; Rabinowitz, J.D. Four Key Steps Control Glycolytic Flux in Mammalian Cells. Cell Syst. 2018, 7, 49–62.e48. [Google Scholar] [CrossRef]
- Shao, Z.; Zhao, H. DNA assembler, an in vivo genetic method for rapid construction of biochemical pathways. Nucleic Acids Res. 2009, 37, e16. [Google Scholar] [CrossRef] [PubMed]
- Hefner, J.; Ketchum, R.E.; Croteau, R. Cloning and functional expression of a cDNA encoding geranylgeranyl diphosphate synthase from Taxus canadensis and assessment of the role of this prenyltransferase in cells induced for taxol production. Arch. BioChem. Biophys. 1998, 360, 62–74. [Google Scholar] [CrossRef]
- Ignea, C.; Trikka, F.A.; Nikolaidis, A.K.; Georgantea, P.; Ioannou, E.; Loupassaki, S.; Kefalas, P.; Kanellis, A.K.; Roussis, V.; Makris, A.M.; et al. Efficient diterpene production in yeast by engineering Erg20p into a geranylgeranyl diphosphate synthase. Metab. Eng. 2015, 27, 65–75. [Google Scholar] [CrossRef]
- Pelot, K.A.; Mitchell, R.; Kwon, M.; Hagelthorn, D.M.; Wardman, J.F.; Chiang, A.; Bohlmann, J.; Ro, D.K.; Zerbe, P. Biosynthesis of the psychotropic plant diterpene salvinorin A: Discovery and characterization of the Salvia divinorum clerodienyl diphosphate synthase. Plant. J. 2017, 89, 885–897. [Google Scholar] [CrossRef]
- Ohnuma, S.; Suzuki, M.; Nishino, T. Archaebacterial ether-linked lipid biosynthetic gene. Expression cloning, sequencing, and characterization of geranylgeranyl-diphosphate synthase. J. Biol. Chem. 1994, 269, 14792–14797. [Google Scholar] [CrossRef]
- Dogbo, O.; Camara, B. Purification of isopentenyl pyrophosphate isomerase and geranylgeranyl pyrophosphate synthase from Capsicum chromoplasts by affinity chromatography. Biochim. Biophys. Acta Lipids Lipid Metab. 1987, 920, 140–148. [Google Scholar] [CrossRef]
- Lohr, M.; Im, C.S.; Grossman, A.R. Genome-based examination of chlorophyll and carotenoid biosynthesis in Chlamydomonas reinhardtii. Plant. Physiol. 2005, 138, 490–515. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, M.; Romer, S.; Suire, C.; Hugueney, P.; Weil, J.H.; Schantz, R.; Camara, B. Identification of a cDNA for the plastid-located geranylgeranyl pyrophosphate synthase from Capsicum annuum: Correlative increase in enzyme activity and transcript level during fruit ripening. Plant. J. 1992, 2, 25–34. [Google Scholar]
- Grabińska, K.; Palamarczyk, G. Dolichol biosynthesis in the yeast Saccharomyces cerevisiae: An insight into the regulatory role of farnesyl diphosphate synthase. FEMS Yeast Res. 2002, 2, 259–265. [Google Scholar]
- Yuan, J.; Ching, C.B. Combinatorial engineering of mevalonate pathway for improved amorpha-4,11-diene production in budding yeast. Biotechnol. Bioeng. 2014, 111, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.S.; Thodey, K.; Trenchard, I.; Smolke, C.D. Advancing secondary metabolite biosynthesis in yeast with synthetic biology tools. FEMS Yeast Res. 2012, 12, 144–170. [Google Scholar] [CrossRef]
- Baek, S.; Utomo, J.; Lee, J.Y.; Dalal, K.; Yoon, Y.J.; Ro, D.K. The yeast platform engineered for synthetic gRNA-landing pads enables multiple gene integrations by a single gRNA/Cas9 system. Metab. Eng. 2021, 64, 111–121. [Google Scholar] [CrossRef]
- Karim, A.S.; Curran, K.A.; Alper, H.S. Characterization of plasmid burden and copy number in Saccharomyces cerevisiae for optimization of metabolic engineering applications. FEMS Yeast Res. 2013, 13, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Berges, T.; Guyonnet, D.; Karst, F. The Saccharomyces cerevisiae mevalonate diphosphate decarboxylase is essential for viability, and a single Leu-to-Pro mutation in a conserved sequence leads to thermosensitivity. J. Bacteriol. 1997, 179, 4664–4670. [Google Scholar] [CrossRef]
- Dimster-Denk, D.; Rine, J. Transcriptional regulation of a sterol-biosynthetic enzyme by sterol levels in Saccharomyces cerevisiae. Mol. Cell Biol. 1996, 16, 3981–3989. [Google Scholar] [CrossRef]
- Paulo, J.A.; O’Connell, J.D.; Gaun, A.; Gygi, S.P. Proteome-wide quantitative multiplexed profiling of protein expression: Carbon-source dependency in Saccharomyces cerevisiae. Mol. Biol. Cell 2015, 26, 4063–4074. [Google Scholar] [CrossRef] [PubMed]
- Jansen, G.; Wu, C.; Schade, B.; Thomas, D.Y.; Whiteway, M. Drag & Drop cloning in yeast. Gene 2005, 344, 43–51. [Google Scholar] [PubMed]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Chen, S.; Zhu, J.; Gao, K.; Zha, W.; Lin, P.; Zi, J. Enhance production of diterpenoids in yeast by overexpression of the fused enzyme of ERG20 and its mutant mERG20. J. Biotechnol. 2020, 307, 29–34. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Strain | Description 1 |
|---|---|
| TXD1 | pESC-URA: Tm-TS |
| TXD2 | pESC-URA: Tm-TS; pIPP |
| TXD3 | pESC-URA: Tm-TS//Sc-BTS1 |
| TXD4 | pESC-URA: Tm-TS//Sc-BTS1; pIPP |
| TXD5 | pESC-URA: Opt Tm-TS//Sc-BTS1; pIPP |
| TXD6 | pESC-URA: Opt Tm-TS~MBP//Sc-BTS1; pIPP |
| TXD7 | pESC-URA: Opt Tm-TS~MBP//Sc-BTS1 Integrated: iADH1: pPYK1-ERG13; pPDC1-tHMGR iPDC1: pTPI1-ERG8; pFBA1-ERG12 |
| TXD8 | pESC-URA: Opt Tm-TS~MBP//Sc-BTS1 Integrated: iADH1: pPYK1-ERG13; pPDC1-tHMGR iPDC1: pTPI1-ERG8; pFBA1-ERG12 iPGK1: pTDH3-ERG10; pTEF1-ERG19 |
| TXD9 | All integrated: iADH1: pPYK1-ERG13; pPDC1-tHMGR iPDC1: pTPI1-ERG8; pFBA1-ERG12 iPGK1: pTDH3-ERG10; pTEF1-ERG19 iTEF2: pGAL1,10-Opt Tm-TS~MBP-ScBTS1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utomo, J.C.; Chaves, F.C.; Bauchart, P.; Martin, V.J.J.; Ro, D.-K. Developing a Yeast Platform Strain for an Enhanced Taxadiene Biosynthesis by CRISPR/Cas9. Metabolites 2021, 11, 147. https://doi.org/10.3390/metabo11030147
Utomo JC, Chaves FC, Bauchart P, Martin VJJ, Ro D-K. Developing a Yeast Platform Strain for an Enhanced Taxadiene Biosynthesis by CRISPR/Cas9. Metabolites. 2021; 11(3):147. https://doi.org/10.3390/metabo11030147
Chicago/Turabian StyleUtomo, Joseph C., Fabio C. Chaves, Philippe Bauchart, Vincent J. J. Martin, and Dae-Kyun Ro. 2021. "Developing a Yeast Platform Strain for an Enhanced Taxadiene Biosynthesis by CRISPR/Cas9" Metabolites 11, no. 3: 147. https://doi.org/10.3390/metabo11030147
APA StyleUtomo, J. C., Chaves, F. C., Bauchart, P., Martin, V. J. J., & Ro, D.-K. (2021). Developing a Yeast Platform Strain for an Enhanced Taxadiene Biosynthesis by CRISPR/Cas9. Metabolites, 11(3), 147. https://doi.org/10.3390/metabo11030147