
Untargeted and Targeted Metabolomic Profiling of Preterm Newborns with EarlyOnset Sepsis: A Case-Control Study

 , ,
, ,
Abstract
1. Introduction
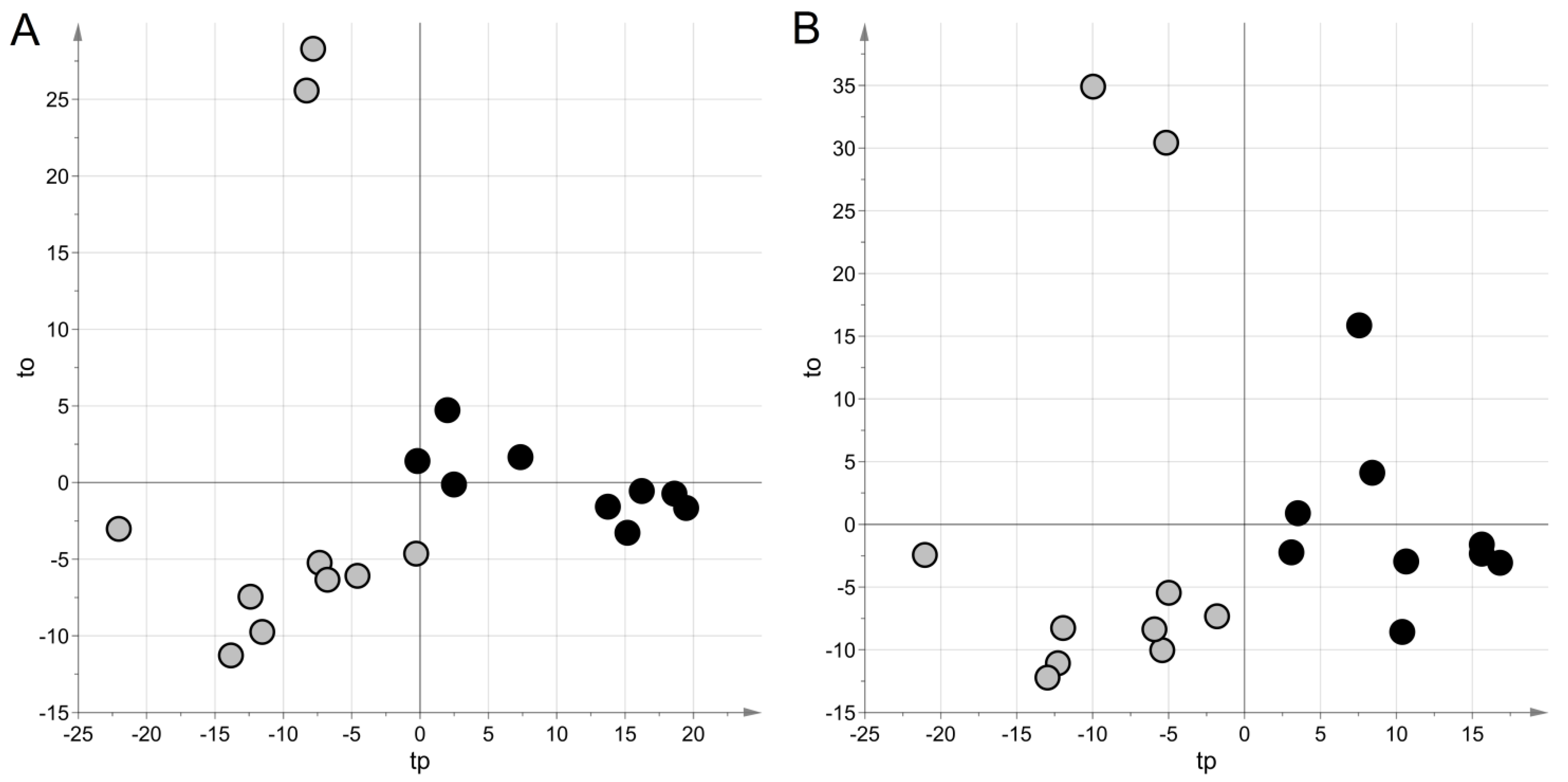
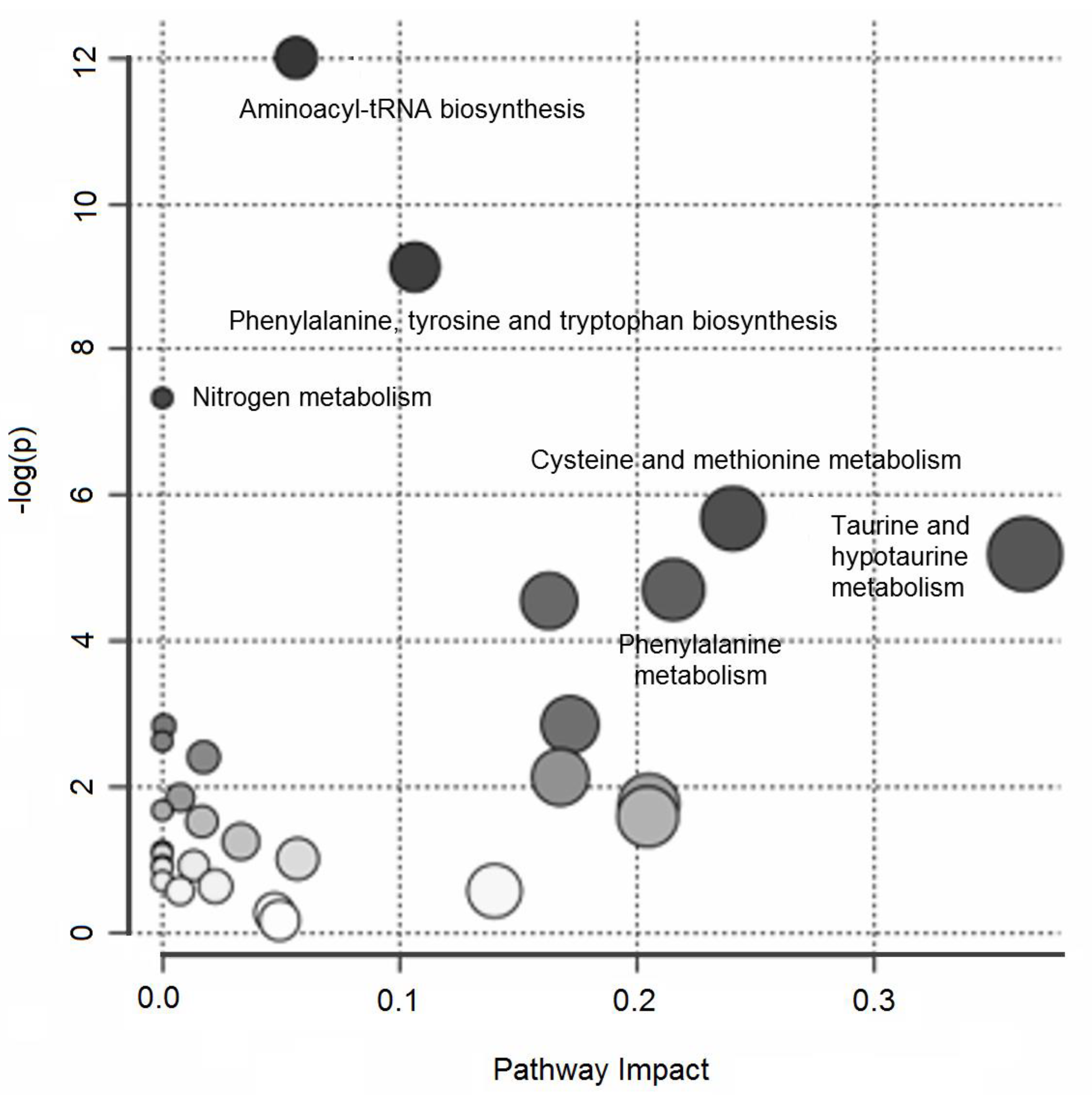
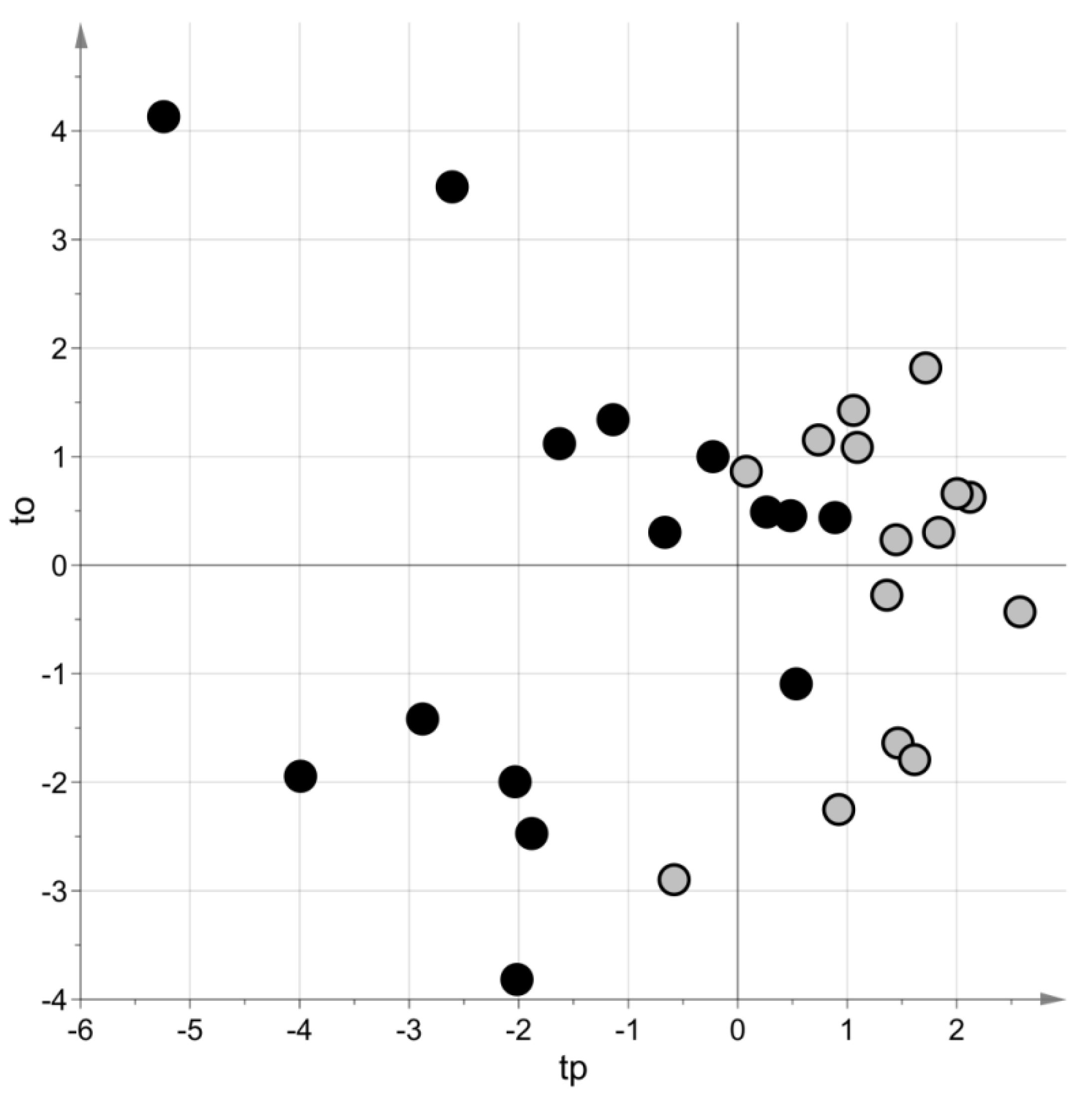
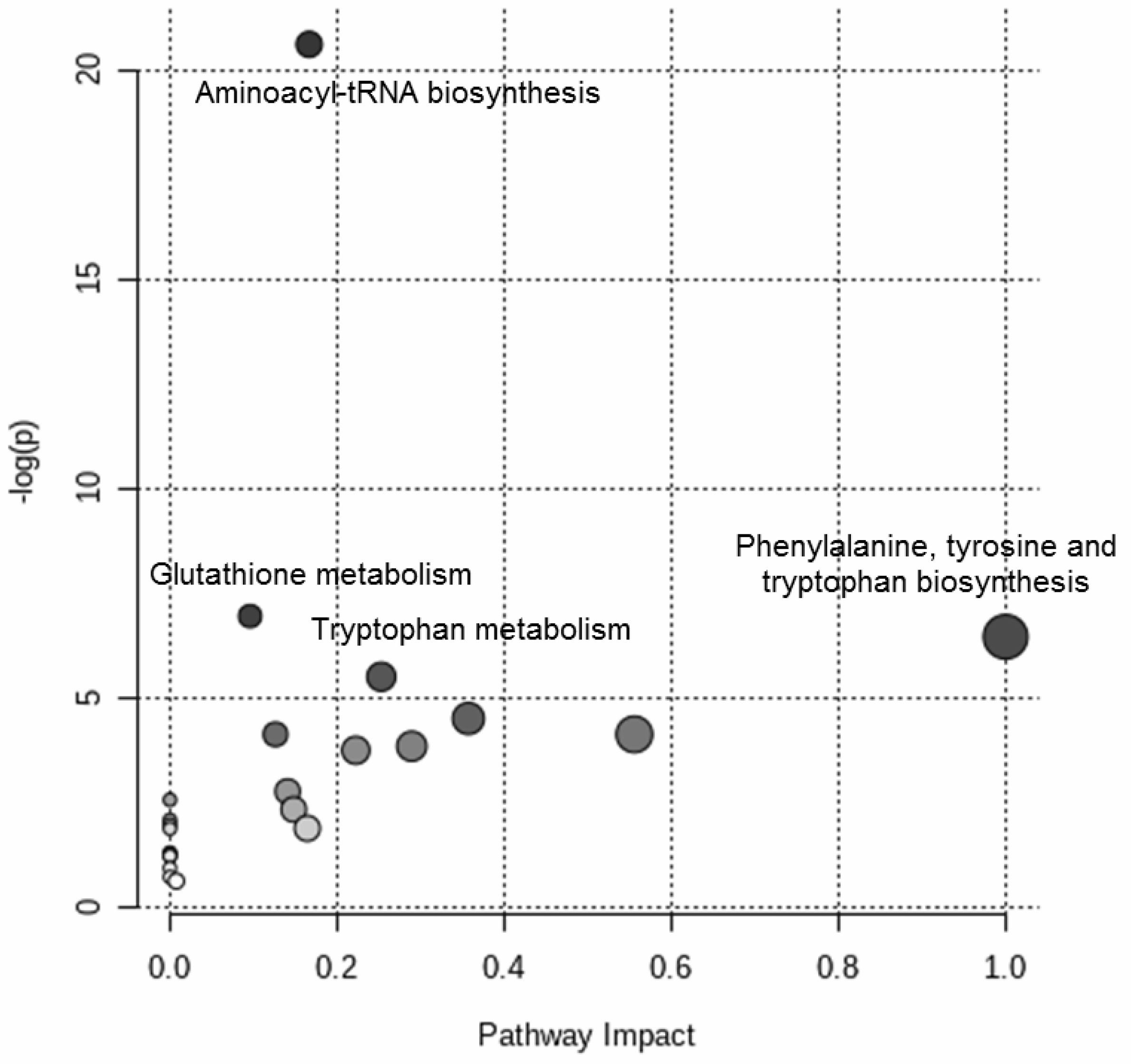
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Sample Collection
4.3. Metabolomic Analysis
4.4. Statistical Data Analysis
4.5. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fleischmann-Struzek, C.; Goldfarb, D.M.; Schlattmann, P.; Schlapbach, L.J.; Reinhart, K.; Kissoon, N. The global burden of paediatric and neonatal sepsis: A systematic review. Lancet Respir. Med. 2018, 6, 223–230. [Google Scholar] [CrossRef]
- Shane, A.L.; Sánchez, P.J.; Stoll, B.J. Neonatal sepsis. Lancet 2017, 390, 1770–1780. [Google Scholar] [CrossRef]
- Strunk, T.; Inder, T.; Wang, X.; Burgner, D.; Mallard, C.; Levy, O. Infection-induced inflammation and cerebral injury in preterm infants. Lancet Infect. Dis. 2014, 14, 751–762. [Google Scholar] [CrossRef]
- Bhutta, Z.A.; Black, R.E. Global maternal, newborn, and child health-so near and yet so far. N. Engl. J. Med. 2013, 369, 2226–2235. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Speer, C.P.; Glaser, K. Beyond sepsis: Staphylococcus epidermidis is an underestimated but significant contributor to neonatal morbidity. Virulence 2018, 9, 621–633. [Google Scholar] [CrossRef]
- Vogel, L. Sepsis kills one million newborns a year: WHO. CMAJ 2017, 189, E1272. [Google Scholar] [CrossRef]
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, regional, and national causes of under-5 mortality in 2000-15: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef]
- Khan, A.M.; Morris, S.K.; Bhutta, Z.A. Neonatal and Perinatal Infections. Pediatr. Clin. North Am. 2017, 64, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Polin, R.A. Newborn CoFa. Management of neonates with suspected or proven early-onset bacterial sepsis. Pediatrics 2012, 129, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Speer, C.P. Late-onset neonatal sepsis: Recent developments. Arch. Dis. Child Fetal. Neonatal Ed. 2015, 100, F257–F263. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Tiwari, S.; Jain, U. Potential biomarkers for effective screening of neonatal sepsis infections: An overview. Microb. Pathog. 2017, 107, 234–242. [Google Scholar] [CrossRef]
- Weston, E.J.; Pondo, T.; Lewis, M.M.; Martell-Cleary, P.; Morin, C.; Jewell, B.; Daily, P.; Apostol, M.; Petit, S.; Farley, M.; et al. The burden of invasive early-onset neonatal sepsis in the United States, 2005–2008. Pediatr. Infect. Dis. J. 2011, 30, 937–941. [Google Scholar] [CrossRef]
- Dong, H.; Cao, H.; Zheng, H. Pathogenic bacteria distributions and drug resistance analysis in 96 cases of neonatal sepsis. BMC Pediatr. 2017, 17, 44. [Google Scholar] [CrossRef]
- Bhandari, V.; Wang, C.; Rinder, C.; Rinder, H. Hematologic profile of sepsis in neonates: Neutrophil CD64 as a diagnostic marker. Pediatrics 2008, 121, 129–134. [Google Scholar] [CrossRef]
- Benitz, W.E. Adjunct laboratory tests in the diagnosis of early-onset neonatal sepsis. Clin. Perinatol. 2010, 37, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Al-Zahrani, A.K.; Ghonaim, M.M.; Hussein, Y.M.; Eed, E.M.; Khalifa, A.S.; Dorgham, L.S. Evaluation of recent methods versus conventional methods for diagnosis of early-onset neonatal sepsis. J. Infect. Dev. Ctries. 2015, 9, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Shukla, S.; Rahman, N.; Gunzler, D.; Abughali, N. Clinical Indicators of Late-Onset Sepsis Workup in Very Low-Birth-Weight Infants in the Neonatal Intensive Care Unit. Am. J. Perinatol. 2016, 33, 856–860. [Google Scholar]
- Carraro, S.; Giordano, G.; Reniero, F.; Perilongo, G.; Baraldi, E. Metabolomics: A new frontier for research in pediatrics. J. Pediatr. 2009, 154, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Li, H.; Xie, A.; Liu, D.; Rao, W.; Lan, L.; Li, X.; Li, F.; Xiao, K.; Wang, H.; et al. Dynamic changes in amino acid concentration profiles in patients with sepsis. PLoS ONE 2015, 10, e0121933. [Google Scholar]
- Kauppi, A.M.; Edin, A.; Ziegler, I.; Mölling, P.; Sjöstedt, A.; Gylfe, A.; Strålin, K.; Johansson, A. Metabolites in Blood for Prediction of Bacteremic Sepsis in the Emergency Room. PLoS ONE 2016, 11, e0147670. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, M.; Cambiaghi, A.; Brunelli, L.; Giordano, S.; Caironi, P.; Guatteri, L.; Raimondi, F.; Gattinoni, L.; Latini, R.; Masson, S.; et al. Mortality prediction in patients with severe septic shock: A pilot study using a target metabolomics approach. Sci. Rep. 2016, 6, 20391. [Google Scholar] [CrossRef]
- Liu, Z.; Yin, P.; Amathieu, R.; Savarin, P.; Xu, G. Application of LC-MS-based metabolomics method in differentiating septic survivors from non-survivors. Anal. Bioanal. Chem. 2016, 408, 7641–7649. [Google Scholar] [CrossRef]
- Garcia-Simon, M.; Morales, J.M.; Modesto-Alapont, V.; Gonzalez-Marrachelli, V.; Vento-Rehues, R.; Jorda-Miñana, A.; Blanquer-Olivas, J.; Monleon, D. Prognosis Biomarkers of Severe Sepsis and Septic Shock by 1H NMR Urine Metabolomics in the Intensive Care Unit. PLoS ONE 2015, 10, e0140993. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.J.; Tsalik, E.L.; van Velkinburgh, J.C.; Glickman, S.W.; Rice, B.J.; Wang, C.; Chen, B.; Carin, L.; Suarez, A.; Mohney, R.P.; et al. An integrated clinico-metabolomic model improves prediction of death in sepsis. Sci. Transl. Med. 2013, 5, 195ra195. [Google Scholar] [CrossRef]
- Seymour, C.W.; Yende, S.; Scott, M.J.; Pribis, J.; Mohney, R.P.; Bell, L.N.; Chen, Y.F.; Zuckerbraun, B.S.; Bigbee, W.L.; Yealy, D.M.; et al. Metabolomics in pneumonia and sepsis: An analysis of the GenIMS cohort study. Intensive Care Med. 2013, 39, 1423–1434. [Google Scholar] [CrossRef] [PubMed]
- Evangelatos, N.; Bauer, P.; Reumann, M.; Satyamoorthy, K.; Lehrach, H.; Brand, A. Metabolomics in Sepsis and Its Impact on Public Health. Public Health Genom. 2017, 20, 274–285. [Google Scholar] [CrossRef]
- Su, L.; Huang, Y.; Zhu, Y.; Xia, L.; Wang, R.; Xiao, K.; Wang, H.; Yan, P.; Wen, B.; Cao, L.; et al. Discrimination of sepsis stage metabolic profiles with an LC/MS-MS-based metabolomics approach. BMJ Open Respir. Res. 2014, 1, e000056. [Google Scholar] [CrossRef] [PubMed]
- Report on the Expert Meeting on Neonatal and Paediatric Sepsis. EMA London 8 June 2010. Available online: https://www.ema.europa.eu/en/events/expert-meeting-neonatal-paediatric-sepsis (accessed on 20 November 2020).
- Stocchero, M.; Locci, E.; d’Aloja, E.; Nioi, M.; Baraldi, E.; Giordano, G. PLS2 in Metabolomics. Metabolites 2019, 9, 51. [Google Scholar] [CrossRef]
- Viant, M.R.; Kurland, I.J.; Jones, M.R.; Dunn, W.B. How close are we to complete annotation of metabolomes? Curr. Opin. Chem. Biol. 2017, 36, 64–69. [Google Scholar] [CrossRef]
- Bohár, Z.; Toldi, J.; Fülöp, F.; Vécsei, L. Changing the face of kynurenines and neurotoxicity: Therapeutic considerations. Int. J. Mol. Sci. 2015, 16, 9772–9793. [Google Scholar] [CrossRef]
- Wlodarska, M.M.; Luo, C.; Kolde, R.; d’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic acid produced by commensal Peptostreptococcus species suppresses inflammation. Cell Host Microbe. 2017, 22, 25–37.e6. [Google Scholar] [CrossRef] [PubMed]
- Hendrikx, T.; Schnabl, S. Indoles: Metabolites produced by intestinal bacteria capable of controlling liver disease manifestation. J. Intern. Med. 2019, 286, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Embleton, N.D.; Marrs, E.C.L.; Smith, D.P.; Fofanova, T.; Nelson, A.; Skeath, T.; Perry, J.D.; Petrosino, J.F.; Berrington, E.; et al. Longitudinal development of the gut microbiome and metabolome in preterm neonates with late onset sepsis and healthy controls. Microbiome 2017, 5, 75. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Cernada, M.; Neu, J.; Pérez-Martínez, G.; Gormaz, M.; Vento, M. Factors influencing gastrointestinal tract and microbiota immune interaction in preterm infants. Pediatr. Res. 2015, 77, 726–731. [Google Scholar] [CrossRef]
- Madan, J.C.; Cowper Salari, R.; Saxena, D.; Davidson, L.; O’Toole, G.A.; Moore, J.H.; Sogin, M.L.; Foster, J.A.; Edwards, W.H.; Palumbo, P.; et al. Gut microbialcolonisation in premature neonates predicts neonatal sepsis. Arch. Dis. Child Fetal. Neonatal Ed. 2012, 97, F456–F462. [Google Scholar] [CrossRef]
- Lyons, J.; Rauh-Pfeiffer, A.; Ming-Yu, Y.; Lu, X.M.; Zurakowski, D.; Curley, M.; Collier, S.; Duggan, C.; Nurko, S.; Thompson, J.; et al. Cysteine metabolism and whole blood glutathione synthesis in septic pediatric patients. Crit. Care Med. 2001, 29, 870–877. [Google Scholar] [CrossRef]
- Mickiewicz, B.; Vogel, H.J.; Wong, H.R.; Winston, B.W. Metabolomics as a novel approach for early diagnosis of pediatric septic shock and its mortality. Am. J. Respir Crit. Care Med. 2013, 187, 967–976. [Google Scholar] [CrossRef]
- Mickiewicz, B.; Thompson, G.C.; Blackwood, J.; Jenne, C.N.; Winston, B.W.; Vogel, H.J.; Joffe, A.R.; Alberta Sepsis Network. Development of metabolic and inflammatory mediator biomarker phenotyping for early diagnosis and triage of pediatric sepsis. Crit. Care 2015, 19, 320. [Google Scholar] [CrossRef]
- Ambroggio, L.; Florin, T.A.; Shah, S.S.; Ruddy, R.; Yeomans, L.; Trexel, J.; Stringer, K.A. Emerging Biomarkers of Illness Severity: Urinary Metabolites Associated with Sepsis and Necrotizing Methicillin-Resistant Staphylococcus aureus Pneumonia. Pharmacotherapy 2017, 37, 1033–1042. [Google Scholar] [CrossRef]
- Fanos, V.; Caboni, P.; Corsello, G.; Stronati, M.; Gazzolo, D.; Noto, A.; Lussu, M.; Dessì, A.; Giuffrè, M.; Lacerenza, S.; et al. Urinary (1)H-NMR and GC-MS metabolomics predicts early and late onset neonatal sepsis. Early Hum. Dev. 2014, 90, S78–S83. [Google Scholar] [CrossRef]
- Sarafidis, K.; ChrysovalantouChatziioannou, A.; Thomaidou, A.; Gika, H.; Mikros, E.; Benaki, D.; Diamanti, E.; Agakidis, C.; Raikos, N.; Drossou, V.; et al. Urine metabolomics in neonates with late-onset sepsis in a case-control study. Sci. Rep. 2017, 7, 45506. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, F.; Montella, S.; Stocchero, M.; Pirillo, P.; Bozzetto, S.; Giordano, G.G.; Poeta, M.; Baraldi, E. Effects of pidotimod and bifidobacteria mixture on clinical symptoms and urinary metabolomic profile of children with recurrent respiratory infections: A randomized placebo-controlled trial. Pulm. Pharmacol. Ther. 2019, 58, 101818. [Google Scholar] [CrossRef] [PubMed]
- Stocchero, M. Relevant and irrelevant predictors in PLS2. J. Chemometr. 2020, 34, e3237. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptive Variable | EOS Group (n = 15) | Controls (n = 15) | p-Value |
|---|---|---|---|
| Gestational age [days] | 207 (17) | 213 (16) | 0.34 |
| Birth weight [g] | 1269 (358) | 1300 (354) | 0.82 |
| Male sex (%) | 7 (47) | 5 (33) | 0.71 |
| Apgar score 1 [min] | 7.0 [1.5] | 7 [1] | 0.32 |
| Apgar score 5 [min] | 8.0 [1.5] | 8 [1] | 0.11 |
| Cesarean section | 14 (93) | 15 (100) | >0.99 |
| Prenatal steroids | 13 (87) | 13 (87) | >0.99 |
| Small for gestational age | 2 (13) | 6 (40) | 0.21 |
| Positive maternal vaginal swab | 3 (20) | 0 (0) | 0.22 |
| Premature rupture of membranes >18 h | 4 (27) | 2 (13) | 0.65 |
| Inotropes | 0 (0) | 1 (7) | >0.99 |
| C-reactive protein <2.9 mg/L–at birth | 12 (80) | 15 (100) | 0.22 |
| White blood count-day 0 [K/μL] | 4.9 [2.8] | 8.2 [6.2] | 0.07 |
| Platelet count-day 0 [K/μL] | 200 [83] | 245 [70] | 0.43 |
| m/z | Type | HMDB ID | Level | Annotation | AUC (CI 95%) |
|---|---|---|---|---|---|
| 175.0244 | CTRL > SEPSIS | HMDB00044 | 2 | Ascorbic acid | 0.66–1.00 |
| 328.0445 | CTRL > SEPSIS | HMDB00058 | 2 | Cyclic AMP | 0.53–1.00 |
| 132.0772 | CTRL > SEPSIS | HMDB00064 | 2 | Creatine | 0.62–1.00 |
| 130.0868 | CTRL > SEPSIS | HMDB00070 | 2 | Pipecolic acid | 0.58–1.00 |
| 152.0575 | CTRL > SEPSIS | HMDB00132 | 1 | Guanine | 0.72–1.00 |
| 464.3013 | CTRL > SEPSIS | HMDB00138 | 1 | Glycocholic acid | 0.66–1.00 |
| 137.0464 | CTRL > SEPSIS | HMDB00157 | 2 | Hypoxanthine | 0.75–1.00 |
| 180.0655 | CTRL > SEPSIS | HMDB00158 | 1 | L-Tyrosine | 0.60–1.00 |
| 164.0708 | CTRL > SEPSIS | HMDB00159 | 1 | L-Phenylalanine | 0.76–1.00 |
| 90.0555 | CTRL > SEPSIS | HMDB00161 | 2 | L-Alanine | 0.64–1.00 |
| 116.0711 | CTRL > SEPSIS | HMDB00162 | 2 | L-Proline | 0.65–1.00 |
| 154.0618 | CTRL > SEPSIS | HMDB00177 | 1 | L-Histidine | 0.60–1.00 |
| 147.1132 | CTRL > SEPSIS | HMDB00182 | 1 | L-Lysine | 0.54–1.00 |
| 239.016 | CTRL > SEPSIS | HMDB00192 | 1 | L-Cystine | 0.58–1.00 |
| 165.0552 | CTRL > SEPSIS | HMDB00205 | 2 | Phenylpyruvic acid | 0.56–1.00 |
| 187.1084 | CTRL > SEPSIS | HMDB00206 | 1 | N6-Acetyl-L-lysine | 0.75–1.00 |
| 335.068 | CTRL > SEPSIS | HMDB00229 | 2 | Nicotinamide ribotide | 0.76–1.00 |
| 126.0224 | CTRL > SEPSIS | HMDB00251 | 1 | Taurine | 0.70–1.00 |
| 283.0677 | CTRL > SEPSIS | HMDB00299 | 2 | Xanthosine | 0.70–1.00 |
| 160.0607 | CTRL > SEPSIS | HMDB00510 | 2 | Aminoadipic acid | 0.57–1.00 |
| 290.1607 | CTRL > SEPSIS | HMDB00552 | 2 | 3-Methylglutarylcarnitine | 0.38–0.95 |
| 120.0123 | CTRL > SEPSIS | HMDB00574 | 1 | L-Cysteine | 0.61–1.00 |
| 135.0309 | CTRL > SEPSIS | HMDB00613 | 2 | Erythronic acid | 0.80–1.00 |
| 269.0601 | CTRL > SEPSIS | HMDB00676 | 2 | L-Homocystine | 0.35–0.94 |
| 609.2639 | SEPSIS > CTRL | HMDB00683 | 2 | Harderoporphyrin | 0.62–1.00 |
| 209.0928 | CTRL > SEPSIS | HMDB00684 | 1 | L-Kynurenine | 0.61–1.00 |
| 130.0867 | CTRL > SEPSIS | HMDB00687 | 1 | L-Leucine | 0.65–1.00 |
| 246.1706 | SEPSIS > CTRL | HMDB00688 | 2 | Isovalerylcarnitine | 0.62–1.00 |
| 130.0506 | CTRL > SEPSIS | HMDB00725 | 1 | Hydroxyproline | 0.65–1.00 |
| 188.0712 | CTRL > SEPSIS | HMDB00734 | 2 | Indoleacrylic acid | 0.64–1.00 |
| 192.0662 | CTRL > SEPSIS | HMDB00763 | 2 | 5-Hydroxyindoleacetic acid | 0.59–1.00 |
| 153.0413 | CTRL > SEPSIS | HMDB00786 | 2 | Oxypurinol | 0.83–1.00 |
| 382.0995 | CTRL > SEPSIS | HMDB00912 | 2 | Succinyladenosine | 0.61–1.00 |
| 158.0815 | CTRL > SEPSIS | HMDB00927 | 2 | Valerylglycine | 0.66–1.00 |
| 203.0818 | CTRL > SEPSIS | HMDB00929 | 1 | L-Tryptophan | 0.70–1.00 |
| 385.1296 | CTRL > SEPSIS | HMDB00939 | 2 | S-Adenosylhomocysteine | 0.45–0.95 |
| 305.0977 | SEPSIS > CTRL | HMDB01067 | 2 | N-Acetylaspartylglutamic acid | 0.50–0.97 |
| 153.0415 | CTRL > SEPSIS | HMDB01182 | 2 | 6,8-Dihydroxypurine | 0.85–1.00 |
| 219.1111 | SEPSIS > CTRL | HMDB01238 | 2 | N-Acetylserotonin | 0.58–1.00 |
| 136.076 | CTRL > SEPSIS | HMDB01250 | 2 | N-Acetylarylamine | 0.62–1.00 |
| 298.1126 | SEPSIS > CTRL | HMDB01563 | 2 | 1-Methylguanosine | 0.67–1.00 |
| 150.0556 | CTRL > SEPSIS | HMDB01859 | 2 | Acetaminophen | 0.70–1.00 |
| 123.0446 | CTRL > SEPSIS | HMDB01870 | 2 | Benzoic acid | 0.60–1.00 |
| 346.1228 | SEPSIS > CTRL | HMDB01913 | 2 | Omeprazole | 0.47–0.98 |
| 86.0605 | CTRL > SEPSIS | HMDB02039 | 2 | 2-Pyrrolidinone | 0.65–1.00 |
| 329.175 | CTRL > SEPSIS | HMDB02121 | 2 | Carnosol | 0.65–1.00 |
| 147.0444 | CTRL > SEPSIS | HMDB02359 | 2 | Phenylpropiolic acid | 0.50–1.00 |
| 175.0606 | SEPSIS > CTRL | HMDB03070 | 2 | Shikimic acid | 0.67–1.00 |
| 337.1276 | SEPSIS > CTRL | HMDB03409 | 2 | Berberine | 0.54–1.00 |
| 179.0559 | SEPSIS > CTRL | HMDB03466 | 2 | L-Gulonolactone | 0.49–1.00 |
| 301.1803 | CTRL > SEPSIS | HMDB03955 | 2 | 19-Hydroxyandrost-4-ene-3,17-dione | 0.65–1.00 |
| 232.028 | CTRL > SEPSIS | HMDB04148 | 2 | Dopamine 4-sulfate | 0.63–1.00 |
| 257.0772 | CTRL > SEPSIS | HMDB04813 | 2 | 3-Methyluridine | 0.52–1.00 |
| 296.1395 | SEPSIS > CTRL | HMDB05037 | 2 | Sumatriptan | 0.50–0.99 |
| 671.5588 | CTRL > SEPSIS | HMDB05233 | 2 | DG(20:1(11Z)/20:4(5Z,8Z,11Z,14Z)/0:0)[iso2] | 0.60–1.00 |
| 290.1356 | CTRL > SEPSIS | HMDB05765 | 2 | Ophthalmic acid | 0.41–0.96 |
| 296.0998 | CTRL > SEPSIS | HMDB05862 | 2 | 2-Methylguanosine | 0.53–1.00 |
| 73.0301 | CTRL > SEPSIS | HMDB06112 | 2 | Malondialdehyde | 0.59–1.00 |
| 275.1128 | SEPSIS > CTRL | 3 | 0.80–1.00 | ||
| 819.2381 | SEPSIS > CTRL | 3 | 0.52–1.00 |
| Name | HMDB ID | Type | AUC (CI 95%) |
|---|---|---|---|
| glycine | HMDB0000123 | SEPSIS > CONTROL | 0.59–0.96 |
| tyrosine | HMDB0000158 | SEPSIS > CONTROL | 0.48–0.89 |
| phenylalanine | HMDB0000159 | SEPSIS > CONTROL | 0.56–1.00 |
| alanine | HMDB0000161 | SEPSIS > CONTROL | 0.50–0.92 |
| proline | HMDB0000162 | SEPSIS > CONTROL | 0.52–0.95 |
| asparagine | HMDB0000168 | SEPSIS > CONTROL | 0.49–0.88 |
| lysine | HMDB0000182 | SEPSIS > CONTROL | 0.48–0.89 |
| serine | HMDB0000187 | SEPSIS > CONTROL | 0.59–0.99 |
| cystine | HMDB0000192 | SEPSIS > CONTROL | 0.52–0.92 |
| ornithine | HMDB0000214 | SEPSIS > CONTROL | 0.59–0.97 |
| serotonin | HMDB0000259 | SEPSIS > CONTROL | 0.49–0.88 |
| sarcosine | HMDB0000271 | SEPSIS > CONTROL | 0.66–1.00 |
| tryptamine | HMDB0000303 | SEPSIS > CONTROL | 0.51–0.91 |
| tyramine | HMDB0000306 | SEPSIS > CONTROL | 0.51–0.91 |
| aminoadipic acid | HMDB0000510 | SEPSIS > CONTROL | 0.52–0.93 |
| kynurenine | HMDB0000684 | SEPSIS > CONTROL | 0.49–0.88 |
| methionine | HMDB0000696 | SEPSIS > CONTROL | 0.49–0.922 |
| kynurenic acid | HMDB0000715 | SEPSIS > CONTROL | 0.51–0.92 |
| 5-HIAA | HMDB0000763 | SEPSIS > CONTROL | 0.48–0.90 |
| xanthurenic acid | HMDB0000881 | SEPSIS > CONTROL | 0.49–0.88 |
| valine | HMDB0000883 | SEPSIS > CONTROL | 0.49–0.92 |
| citrulline | HMDB0000904 | SEPSIS > CONTROL | 0.52–0.92 |
| spermidine | HMDB0001257 | SEPSIS > CONTROL | 0.48–0.89 |
| N1-AcetylSPD | HMDB0001276 | SEPSIS > CONTROL | 0.49–0.89 |
| ADMA | HMDB0001539 | SEPSIS > CONTROL | 0.49–0.92 |
| cadaverine | HMDB0002322 | SEPSIS > CONTROL | 0.64–0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mardegan, V.; Giordano, G.; Stocchero, M.; Pirillo, P.; Poloniato, G.; Donadel, E.; Salvadori, S.; Giaquinto, C.; Priante, E.; Baraldi, E. Untargeted and Targeted Metabolomic Profiling of Preterm Newborns with EarlyOnset Sepsis: A Case-Control Study. Metabolites 2021, 11, 115. https://doi.org/10.3390/metabo11020115
Mardegan V, Giordano G, Stocchero M, Pirillo P, Poloniato G, Donadel E, Salvadori S, Giaquinto C, Priante E, Baraldi E. Untargeted and Targeted Metabolomic Profiling of Preterm Newborns with EarlyOnset Sepsis: A Case-Control Study. Metabolites. 2021; 11(2):115. https://doi.org/10.3390/metabo11020115
Chicago/Turabian StyleMardegan, Veronica, Giuseppe Giordano, Matteo Stocchero, Paola Pirillo, Gabriele Poloniato, Enrica Donadel, Sabrina Salvadori, Carlo Giaquinto, Elena Priante, and Eugenio Baraldi. 2021. "Untargeted and Targeted Metabolomic Profiling of Preterm Newborns with EarlyOnset Sepsis: A Case-Control Study" Metabolites 11, no. 2: 115. https://doi.org/10.3390/metabo11020115
APA StyleMardegan, V., Giordano, G., Stocchero, M., Pirillo, P., Poloniato, G., Donadel, E., Salvadori, S., Giaquinto, C., Priante, E., & Baraldi, E. (2021). Untargeted and Targeted Metabolomic Profiling of Preterm Newborns with EarlyOnset Sepsis: A Case-Control Study. Metabolites, 11(2), 115. https://doi.org/10.3390/metabo11020115