Vibrio harveyi Infection Significantly Alters Amino Acid and Carbohydrate Metabolism in Whiteleg Shrimp, Litopenaeus vannamei

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. HR-MAS 1H-NMR Spectra of the Gills, Hepatopancreas, and Haemolymph of Whiteleg Shrimp
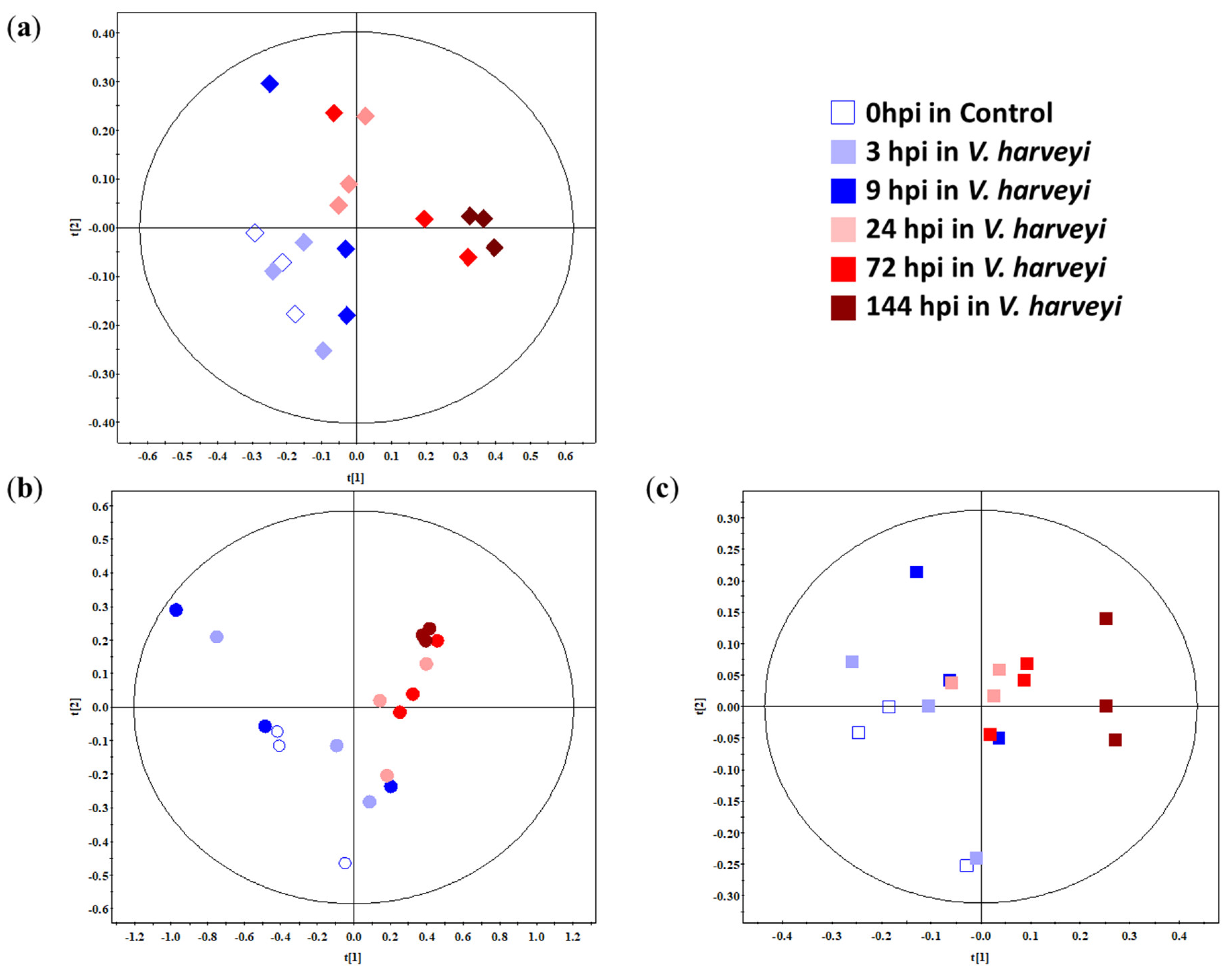
2.2. Multivariate Analysis of the Gills, Hepatopancreas, and Haemolymph of Whiteleg Shrimp
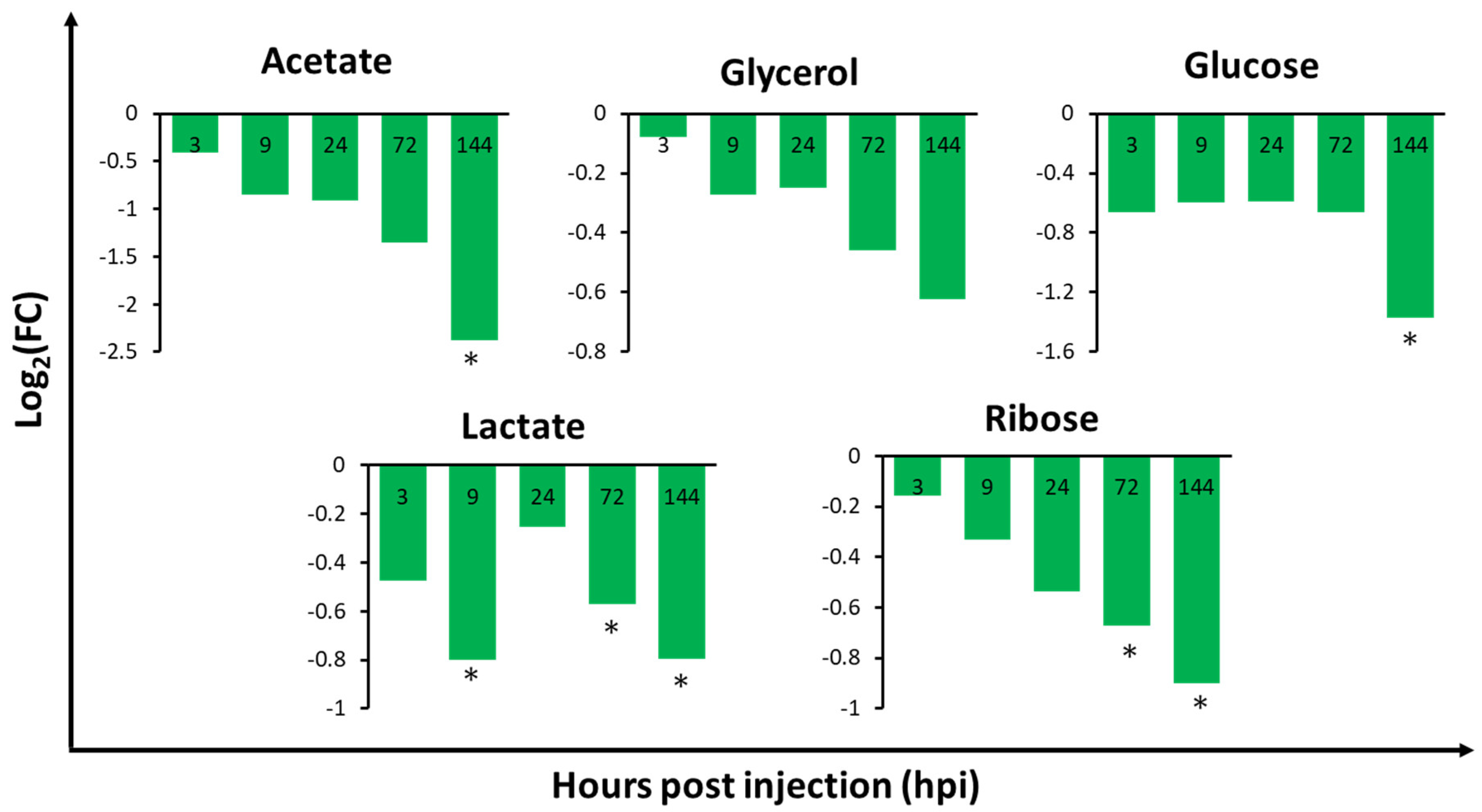
2.3. Relative Metabolite Concentrations in the Gills, Hepatopancreas, and Haemolymph of V. harveyi-Infected Whiteleg Shrimp
2.4. Metabolic Pathway Analysis of V. harveyi-Infected Whiteleg Shrimp
2.4.1. Metabolic Pathways Related to Carbohydrate Metabolism in the Tissues of Infected Whiteleg Shrimp
2.4.2. Metabolic Pathways Related to Amino Acid Metabolism in the Tissues of Infected Whiteleg Shrimp
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Bacterial Challenge
4.3. Sample Preparation
4.4. 1H-NMR Measurements
4.5. Spectral Pre-Processing
4.6. Multivariate Statistical Analysis
4.7. Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fisheries, F. Aquaculture Department. In The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2018; Available online: http://www.fao.org/3/i9540en/I9540EN.pdf (accessed on 10 January 2020).
- Zheng, C.N.; Wang, W. Effects of Lactobacillus pentosus on the growth performance, digestive enzyme and disease resistance of white shrimp, Litopenaeus vannamei (Boone, 1931). Aquac. Res. 2017, 48, 2767–2777. [Google Scholar] [CrossRef]
- Gomez-Gil, B.; Tron-Mayen, L.; Roque, A.; Turnbull, J.F.; Inglis, V.; Guerra-Flores, A.L. Species of Vibrio isolated from hepatopancreas, haemolymph and digestive tract of a population of healthy juvenile Penaeus vannamei. Aquaculture 1998, 163, 1–9. [Google Scholar] [CrossRef]
- Austin, B.; Zhang, X.H. Vibrio harveyi: A significant pathogen of marine vertebrates and invertebrates. Lett. Appl. Microbiol. 2006, 43, 119–124. [Google Scholar] [CrossRef]
- Chatterjee, S.; Haldar, S. Vibrio related diseases in aquaculture and development of rapid and accurate identification methods. J. Marine Sci. Res. Dev. S 2012, 1. [Google Scholar] [CrossRef]
- Travers, M.-A.; Le Goïc, N.; Huchette, S.; Koken, M.; Paillard, C. Summer immune depression associated with increased susceptibility of the European abalone, Haliotis tuberculata to Vibrio harveyi infection. Fish. Shellfish Immun. 2008, 25, 800–880. [Google Scholar] [CrossRef]
- Talpur, A.D. Mentha piperita (Peppermint) as feed additive enhanced growth performance, survival, immune response and disease resistance of Asian seabass, Lates calcarifer (Bloch) against Vibrio harveyi infection. Aquaculture 2014, 420, 71–78. [Google Scholar] [CrossRef]
- Liu, X.; Ji, C.; Zhao, J.; Wang, Q.; Li, F.; Wu, H. Metabolic profiling of the tissue-specific responses in mussel Mytilus galloprovincialis towards Vibrio harveyi challenge. Fish. Shellfish Immun. 2014, 39, 372–377. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Miyazaki, T.; Limsuwan, C. Histopathology, biochemistry, and pathogenicity of Vibrio harveyi infecting black tiger prawn Penaeus monodon. J. Aquat. Anim. Health 1994, 6, 27–35. [Google Scholar] [CrossRef]
- Huang, H.-H.; Liu, X.-L.; Xiang, J.-H.; Wang, P. Immune response of Litopenaeus vannamei after infection with Vibrio harveyi. Aquaculture 2013, 406, 115–120. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant. Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Krastanov, A. Metabolomics—The state of art. Biotechnol. Biotechnol. Equip. 2010, 24, 1537–1543. [Google Scholar] [CrossRef]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692. [Google Scholar] [CrossRef]
- Ji, C.; Wu, H.; Wei, L.; Zhao, J.; Wang, Q.; Lu, H. Responses of Mytilus galloprovincialis to bacterial challenges by metabolomics and proteomics. Fish. Shellfish Immun. 2013, 35, 489–498. [Google Scholar] [CrossRef]
- Wu, H.; Ji, C.; Wei, L.; Zhao, J.; Lu, H. Proteomic and metabolomic responses in hepatopancreas of Mytilus galloprovincialis challenged by Micrococcus luteus and Vibrio anguillarum. J. Proteom. 2013, 94, 54–67. [Google Scholar] [CrossRef]
- Liu, X.; Ji, C.; Zhao, J.; Wu, H. Differential metabolic responses of clam Ruditapes philippinarum to Vibrio anguillarum and Vibrio splendidus challenges. Fish. Shellfish Immun. 2013, 35, 2001–2007. [Google Scholar] [CrossRef]
- Ye, Y.; Xia, M.; Mu, C.; Li, R.; Wang, C. Acute metabolic response of Portunus trituberculatus to Vibrio alginolyticus infection. Aquaculture 2016, 463, 201–208. [Google Scholar] [CrossRef]
- Dumas, M.-E.; Maibaum, E.C.; Teague, C.; Ueshima, H.; Zhou, B.; Lindon, J.C.; Nicholson, J.K.; Stamler, J.; Elliott, P.; Chan, Q. Assessment of analytical reproducibility of 1H NMR spectroscopy based metabonomics for large-scale epidemiological research: The INTERMAP Study. Anal. Chem. 2006, 78, 2199–2208. [Google Scholar] [CrossRef]
- Pongsomboon, S.; Wongpanya, R.; Tang, S.; Chalorsrikul, A.; Tassanakajon, A. Abundantly expressed transcripts in the lymphoid organ of the black tiger shrimp, Penaeus monodon, and their implication in immune function. Fish. Shellfish Immun. 2008, 25, 485–493. [Google Scholar] [CrossRef]
- Silveira, A.S.; Matos, G.M.; Falchetti, M.; Ribeiro, F.S.; Bressan, A.; Bachère, E.; Perazzolo, L.M.; Rosa, R.D. An immune-related gene expression atlas of the shrimp digestive system in response to two major pathogens brings insights into the involvement of hemocytes in gut immunity. Dev. Comp. Immunol. 2018, 79, 44–50. [Google Scholar] [CrossRef]
- Somboonwiwat, K.; Chaikeeratisak, V.; Wang, H.-C.; Lo, C.F.; Tassanakajon, A. Proteomic analysis of differentially expressed proteins in Penaeus monodon hemocytes after Vibrio harveyi infection. Proteome Sci. 2010, 8, 39. [Google Scholar] [CrossRef]
- Rattanama, P.; Thompson, J.R.; Kongkerd, N.; Srinitiwarawong, K.; Vuddhakul, V.; Mekalanos, J.J. Sigma E regulators control hemolytic activity and virulence in a shrimp pathogenic Vibrio harveyi. PLoS ONE 2012, 7, e32523. [Google Scholar] [CrossRef]
- Chang, Y.; Xing, J.; Tang, X.; Sheng, X.; Zhan, W. Haemocyanin content of shrimp (Fenneropenaeus chinensis) associated with white spot syndrome virus and Vibrio harveyi infection process. Fish. Shellfish Immun. 2016, 48, 185–189. [Google Scholar] [CrossRef]
- Li, T.; Li, E.; Suo, Y.; Xu, Z.; Jia, Y.; Qin, J.G.; Chen, L.; Gu, Z. Energy metabolism and metabolomics response of Pacific white shrimp Litopenaeus vannamei to sulfide toxicity. Aquat. Toxicol. 2017, 183, 28–37. [Google Scholar] [CrossRef]
- Zhou, H.; Li, Y.; Wei, L.; Zhang, Z.; Huang, H.; Diao, X.; Xiang, J. Sensitivity of larvae and adult and the immunologic characteristics of Litopenaeus vannamei under the acute hypoxia. J. Chem. 2014, 2014. [Google Scholar] [CrossRef]
- Huynh, T.-G.; Cheng, A.-C.; Chi, C.-C.; Chiu, K.-H.; Liu, C.-H. A synbiotic improves the immunity of white shrimp, Litopenaeus vannamei: Metabolomic analysis reveal compelling evidence. Fish. Shellfish Immun. 2018, 79, 284–293. [Google Scholar] [CrossRef]
- Liu, P.-f.; Liu, Q.-h.; Wu, Y.; Jie, H. A pilot metabolic profiling study in hepatopancreas of Litopenaeus vannamei with white spot syndrome virus based on 1H NMR spectroscopy. J. Invertebr. Pathol. 2015, 124, 51–56. [Google Scholar] [CrossRef]
- Soto-Rodriguez, S.; Simoes, N.; Jones, D.; Roque, A.; Gomez-Gil, B. Assessment of fluorescent-labeled bacteria for evaluation of in vivo uptake of bacteria (Vibrio spp.) by crustacean larvae. J. Microbiol. Meth. 2003, 52, 101–114. [Google Scholar] [CrossRef]
- Soonthornchai, W.; Rungrassamee, W.; Karoonuthaisiri, N.; Jarayabhand, P.; Klinbunga, S.; Söderhäll, K.; Jiravanichpaisal, P. Expression of immune-related genes in the digestive organ of shrimp, Penaeus monodon, after an oral infection by Vibrio harveyi. Dev. Comp. Immunol. 2010, 34, 19–28. [Google Scholar] [CrossRef]
- Mohajeri, J.; Afsharnasab, M.; Jalali, B.; Kakoolaki, S.; Sharifrohani, M.; Haghighi, A. Immunological and histopathological changes in Penaeus semisulcatus challenged with Vibrio harveyi. Iran. J. Fish. Sci. 2011, 10, 254–265. Available online: http://jifro.ir/article-1-173-en.html (accessed on 2 March 2020).
- Chien, C.-C.; Lin, T.-Y.; Chi, C.-C.; Liu, C.-H. Probiotic, Bacillus subtilis E20 alters the immunity of white shrimp, Litopenaeus vannamei via glutamine metabolism and hexosamine biosynthetic pathway. Fish. Shellfish Immun. 2020. [Google Scholar] [CrossRef]
- Henry, R.P.; Lucu, C.; Onken, H.; Weihrauch, D. Multiple functions of the crustacean gill: Osmotic/ionic regulation, acid-base balance, ammonia excretion, and bioaccumulation of toxic metals. Front. Physiol. 2012, 3, 431. [Google Scholar] [CrossRef]
- Martin, G.G.; Quintero, M.; Quigley, M.; Khosrovian, H. Elimination of sequestered material from the gills of decapod crustaceans. J. Crust. Biol. 2000, 20, 209–217. [Google Scholar] [CrossRef][Green Version]
- Fan, W.; Ye, Y.; Chen, Z.; Shao, Y.; Xie, X.; Zhang, W.; Liu, H.-p.; Li, C. Metabolic product response profiles of Cherax quadricarinatus towards white spot syndrome virus infection. Dev. Comp. Immunol. 2016, 61, 236–241. [Google Scholar] [CrossRef]
- Yoganandhan, K.; Thirupathi, S.; Hameed, A.S. Biochemical, physiological and hematological changes in white spot syndrome virus-infected shrimp, Penaeus indicus. Aquaculture 2003, 221, 1–11. [Google Scholar] [CrossRef]
- Gannon, A.T.; Demarco, V.G.; Morris, T.; Wheatly, M.G.; Kao, Y.-H. Oxygen uptake, critical oxygen tension, and available oxygen for three species of cave crayfishes. J. Crust. Biol. 1999, 19, 235–243. [Google Scholar] [CrossRef]
- Su, M.-A.; Huang, Y.-T.; Chen, I.-T.; Lee, D.-Y.; Hsieh, Y.-C.; Li, C.-Y.; Ng, T.H.; Liang, S.-Y.; Lin, S.-Y.; Huang, S.-W. An invertebrate Warburg effect: A shrimp virus achieves successful replication by altering the host metabolome via the PI3K-Akt-mTOR pathway. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, J.; He, Y.; Zhou, J.; Yan, J.; Jiang, M. A metabolic study in hepatopancreas of Litopenaeus vannamei response to white spot syndrome virus. Int. Aquat. Res. 2017, 9, 195–201. [Google Scholar] [CrossRef]
- Zala, D.; Schlattner, U.; Desvignes, T.; Bobe, J.; Roux, A.; Chavrier, P.; Boissan, M. The advantage of channeling nucleotides for very processive functions. F1000Research 2017, 6. [Google Scholar] [CrossRef]
- Leaño, E.M.; Lavilla-Pitogo, C.R.; Paner, M.G. Bacterial flora in the hepatopancreas of pond-reared Penaeus monodon juveniles with luminous vibriosis. Aquaculture 1998, 164, 367–374. [Google Scholar] [CrossRef]
- Khimmakthong, U.; Sukkarun, P. The spread of Vibrio parahaemolyticus in tissues of the Pacific white shrimp Litopenaeus vannamei analyzed by PCR and histopathology. Microb. Pathog. 2017, 113, 107–112. [Google Scholar] [CrossRef]
- Aguilera-Rivera, D.; Escalante-Herrera, K.; Gaxiola, G.; Prieto-Davó, A.; Rodríguez-Fuentes, G.; Guerra-Castro, E.; Hernández-López, J.; Chávez-Sánchez, M.C.; Rodríguez-Canul, R. Immune response of the Pacific white shrimp, Litopenaeus vannamei, previously reared in biofloc and after an infection assay with Vibrio harveyi. J. World. Aquac. Soc. 2019, 50, 119–136. [Google Scholar] [CrossRef]
- Shinji, J.; Wilder, M.N. Dynamics of free amino acids in the hemolymph of Pacific whiteleg shrimp Litopenaeus vannamei exposed to different types of stress. Fish. Sci. 2012, 78, 1187–1194. [Google Scholar] [CrossRef]
- Barton, B.A.; Schreck, C.B. Metabolic cost of acute physical stress in juvenile steelhead. Trans. Am. Fish. Soc. 1987, 116, 257–263. [Google Scholar] [CrossRef]
- Bishop, J.S.; Burton, R.S. Amino acid synthesis during hyperosmotic stress in Penaeus aztecus postlarvae. Comp. Biochem. Phys. A Physiol. 1993, 106, 49–56. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, A.-L.; Xian, J.-A. Variation of free amino acid and carbohydrate concentrations in white shrimp, Litopenaeus vannamei: Effects of continuous cold stress. Aquaculture 2011, 317, 182–186. [Google Scholar] [CrossRef]
- Iacobazzi, V.; Vittoria, I. Citrate-new functions for an old metabolite. Biol. Chem. 2014, 395, 387–399. [Google Scholar] [CrossRef]
- Su, X.; Li, X. The improvement of growth, digestive enzyme activity and disease resistance of white shrimp by the dietary citric acid. Aquac Int. 2014, 22, 1823–1835. [Google Scholar] [CrossRef]
- Mercier, P.; Lewis, M.J. Towards automatic metabolomics profiling of high-resolution one-dimensional proton NMR spectra. J. Biomol NMR. 2011, 49, 307–323. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | |||||||||
| Metabolism | Pathway | 3 hpi | 9 hpi | 24 hpi | 72 hpi | 144 hpi | Hit | Metabolite(s) | Impact |
| Carbohydrate metabolism | Pyruvate metabolism | - | - | - | 0.041 | 0.012 | 3/22 | Pyruvate, Lactate, Acetate | 0.24 |
| Carbohydrate metabolism | Glycolysis / Gluconeogenesis | - | - | - | - | 0.020 | 3/26 | Pyruvate, Lactate, Acetate | 0.13 |
| Amino acid metabolism | Tyrosine metabolism | - | - | - | 0.045 | - | 2/33 | Tyrosine, Pyruvate | 0.13 |
| (b) | |||||||||
| Metabolism | Pathway | 3 hpi | 9 hpi | 24 hpi | 72 hpi | 144 hpi | Hit | Metabolite(s) | Impact |
| Amino acid metabolism | Tyrosine metabolism | 0.009 | - | 0.004 | - | 0.0005 | 2/33 | Tyrosine, Fumarate | 0.19 |
| Amino acid metabolism | Cysteine and methionine metabolism | - | 0.045 | - | - | 0.0002 | 2/33 | Serine, Methionine | 0.14 |
| Amino acid metabolism | Phenylalanine, tyrosine and tryptophan biosynthesis | - | - | 0.012 | - | 0.005 | 2/4 | Tyrosine, Phenylalanine | 1.00 |
| Amino acid metabolism | Phenylalanine metabolism | - | - | 0.012 | - | 0.005 | 2/8 | Tyrosine, Phenylalanine | 0.38 |
| Amino acid metabolism | Alanine, aspartate and glutamate metabolism | - | - | 0.019 | 0.005 | 0.014 | 7/27 | Aspartate, Asparagine, Alanine, Fumarate, Glutamine, Glutamate, Succinate | 0.61 |
| Nucleotide metabolism | Pyrimidine metabolism | - | - | - | - | 0.003 | 2/41 | Glutamine, Uracil | 0.10 |
| Metabolism of other amino acids | D-Glutamine and D-glutamate metabolism | - | - | - | - | 0.034 | 2/6 | Glutamate, Glutamine | 1.00 |
| Carbohydrate metabolism | Glyoxylate and dicarboxylate metabolism | - | - | - | - | 0.049 | 4/32 | Serine, Glycine, Glutamate, Glutamine | 0.19 |
| (c) | |||||||||
| Metabolism | Pathway | 3 hpi | 9 hpi | 24 hpi | 72 hpi | 144 hpi | Hit | Metabolite(s) | Impact |
| Carbohydrate metabolism | Glyoxylate and dicarboxylate metabolism | 0.033 | 0.028 | 0.021 | - | 0.024 | 4/24 | Acetate, Citrate, Glycine, Glutamate | 0.17 |
| Carbohydrate metabolism | Citrate cycle (TCA cycle) | 0.04 | 0.034 | 0.02 | - | 0.024 | 3/20 | Citrate, Succinate, Pyruvate | 0.17 |
| Metabolism of other amino acids | Glutathione metabolism | 0.041 | - | - | - | - | 2/26 | Glutamate, Glycine | 0.13 |
| Amino acid metabolism | Glycine, serine and threonine metabolism | - | 0.011 | - | - | 0.036 | 5/33 | Betaine, Choline, Creatine, Glycine, Pyruvate | 0.29 |
| Carbohydrate metabolism | Pyruvate metabolism | - | - | - | - | 0.013 | 3/22 | Acetate, Lactate, Pyruvate | 0.24 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, S.; Kim, A.; Lee, W.; Kim, S.; Lee, S.; Yoon, D.; Bae, J.-S.; Park, C.-I.; Kim, S. Vibrio harveyi Infection Significantly Alters Amino Acid and Carbohydrate Metabolism in Whiteleg Shrimp, Litopenaeus vannamei. Metabolites 2020, 10, 265. https://doi.org/10.3390/metabo10060265
Ma S, Kim A, Lee W, Kim S, Lee S, Yoon D, Bae J-S, Park C-I, Kim S. Vibrio harveyi Infection Significantly Alters Amino Acid and Carbohydrate Metabolism in Whiteleg Shrimp, Litopenaeus vannamei. Metabolites. 2020; 10(6):265. https://doi.org/10.3390/metabo10060265
Chicago/Turabian StyleMa, Seohee, Ahran Kim, Wonho Lee, Seonghye Kim, Sujin Lee, Dahye Yoon, Jin-Sol Bae, Chan-Il Park, and Suhkmann Kim. 2020. "Vibrio harveyi Infection Significantly Alters Amino Acid and Carbohydrate Metabolism in Whiteleg Shrimp, Litopenaeus vannamei" Metabolites 10, no. 6: 265. https://doi.org/10.3390/metabo10060265
APA StyleMa, S., Kim, A., Lee, W., Kim, S., Lee, S., Yoon, D., Bae, J.-S., Park, C.-I., & Kim, S. (2020). Vibrio harveyi Infection Significantly Alters Amino Acid and Carbohydrate Metabolism in Whiteleg Shrimp, Litopenaeus vannamei. Metabolites, 10(6), 265. https://doi.org/10.3390/metabo10060265