The Urine Metabolome of Young Autistic Children Correlates with Their Clinical Profile Severity

 ,
, 
 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
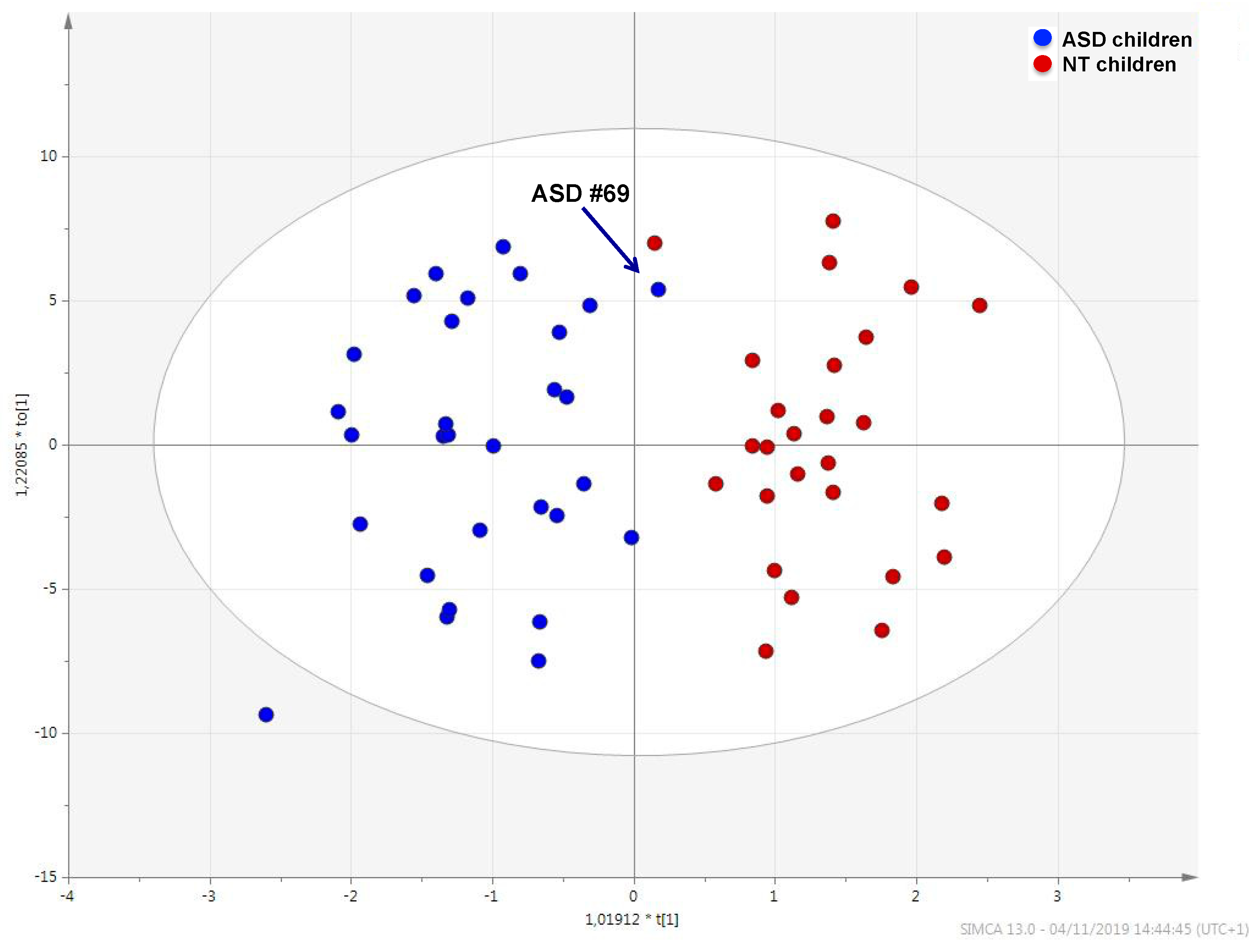
2.1. Urine Metabolic Profile in ASD Children and NT Children
2.2. Association between ASD Core Symptoms and the Urine Metabolome
2.3. Association between Repetitive, Problematic Abnormal Behaviors and the Urine Metabolome
3. Discussion
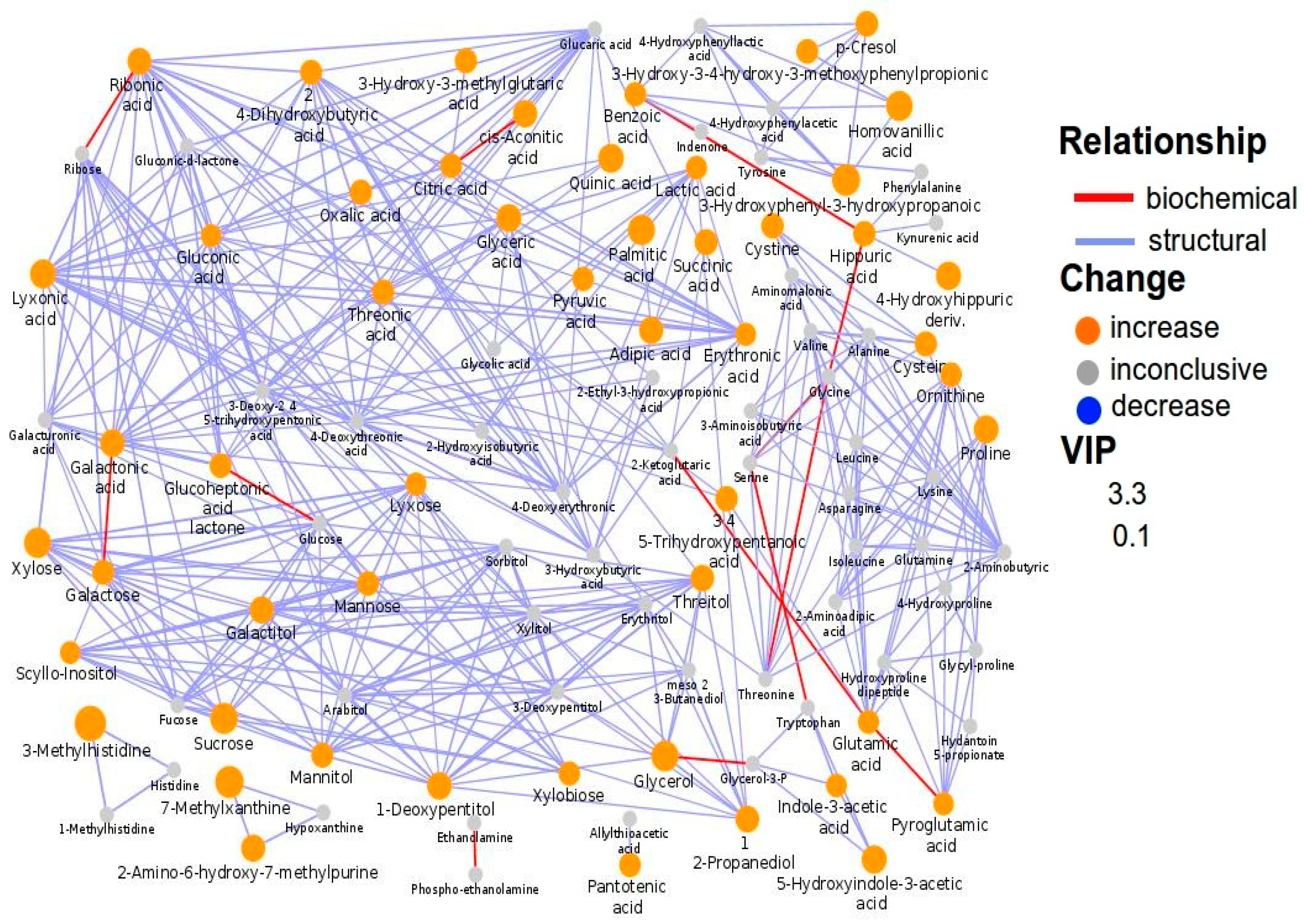
3.1. The Metabolic Profile of Autistic Children
3.2. The Urine Metabolome Reflects Autism Core Symptoms Severity
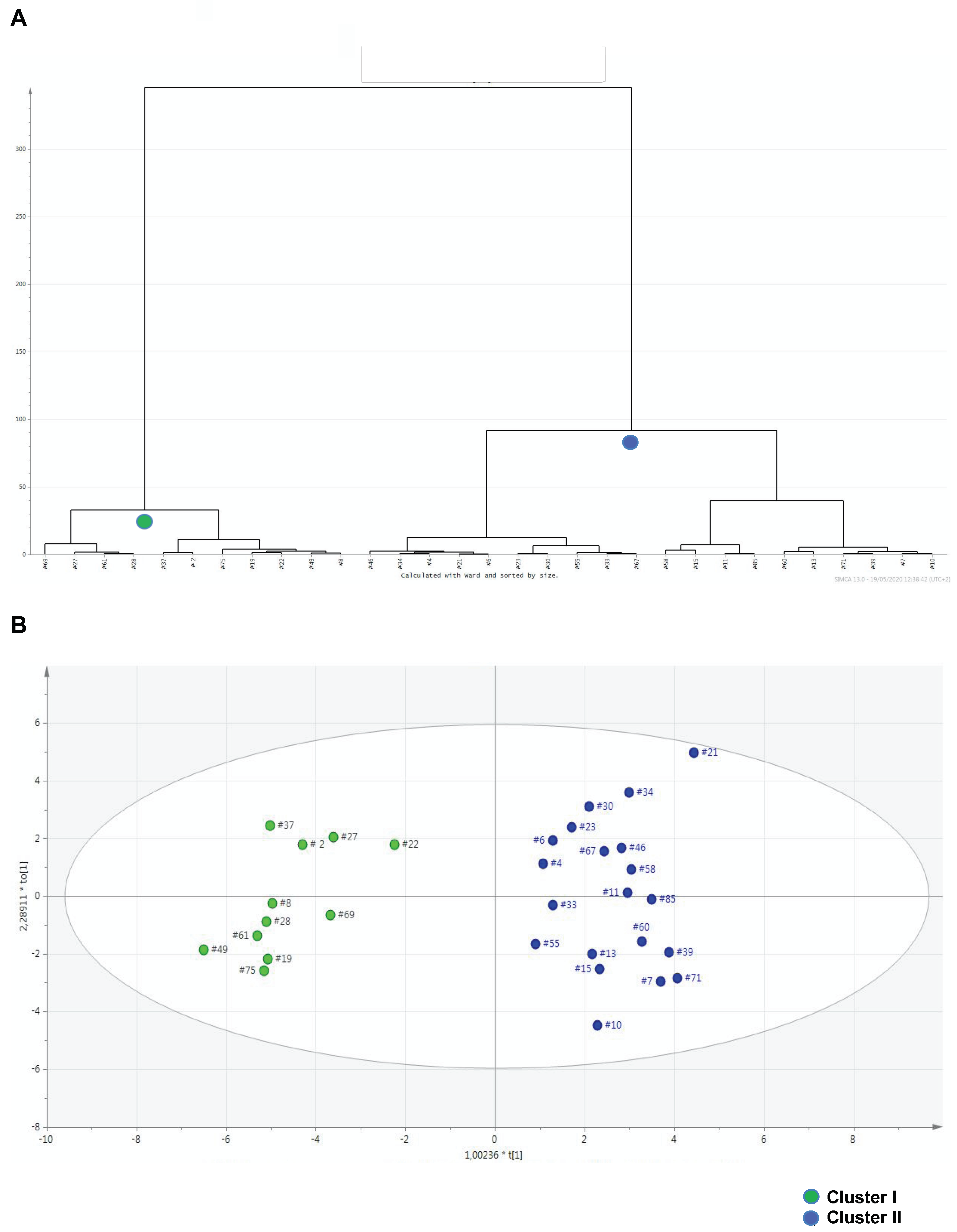
3.3. Metabolomics Contributes to Identifying Clinical Thresholds Discriminating Severe from Moderate Behavioral Impairments
3.4. Limitations of the Study
4. Materials and Methods
4.1. Participants
4.2. Primary Behavioral Outcome Measures in Autistic Children
4.3. Sample Collection, Storage, and Preparation
4.4. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4.5. Data Processing and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lyall, K.; Croen, L.; Daniels, J.; Fallin, M.D.; Ladd-Acosta, C.; Lee, B.K.; Park, B.Y.; Snyder, N.W.; Schendel, D.; Volk, H.; et al. The Changing Epidemiology of Autism Spectrum Disorders. Annu. Rev. Public Heal. 2017, 38, 81–102. [Google Scholar] [CrossRef]
- Lord, C.; Brugha, T.S.; Charman, T.; Cusack, J.; Dumas, G.; Frazier, T.; Jones, E.J.H.; Jones, R.M.; Pickles, A.; State, M.W.; et al. Autism spectrum disorder. Nat. Rev. Dis. Primers 2020, 6, 5. [Google Scholar] [PubMed]
- Maenner, M.J.; Shaw, K.A.; Baio, J.; Washington, A.; Patrick, M.; DiRienzo, M.; Christensen, D.L.; Wiggins, L.D.; Pettygrove, S.; Andrews, J.G.; et al. Prevalence of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2016. MMWR Surveill Summ. 2020, 69, 1–12. [Google Scholar] [PubMed]
- Leigh, J.P.; Du, J. Brief Report: Forecasting the Economic Burden of Autism in 2015 and 2025 in the United States. J. Autism Dev. Disord. 2015, 45, 4135–4139. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Yip, B.H.K.; Windham, G.C.; Sourander, A.; Francis, R.; Yoffe, R.; Glasson, E.; Mahjani, B.; Suominen, A.; Leonard, H.; et al. Association of Genetic and Environmental Factors With Autism in a 5-Country Cohort. JAMA Psychiatry 2019, 76, 1035–1043. [Google Scholar] [CrossRef]
- Svoboda, E. Could the gut microbiome be linked to autism? Nat. Cell Biol. 2020, 577, S14–S15. [Google Scholar] [CrossRef]
- Zamboni, N.; Saghatelian, A.; Patti, G.J. Defining the Metabolome: Size, Flux, and Regulation. Mol. Cell 2015, 58, 699–706. [Google Scholar] [CrossRef]
- Mussap, M.; Noto, A.; Fanos, V. Metabolomics of autism spectrum disorders: Early insights regarding mammalian-microbial cometabolites. Expert Rev. Mol. Diagn. 2016, 16, 869–881. [Google Scholar] [CrossRef]
- Salerno, C.; Crifò, C. Urinary Methylxanthine and Autistic Disorder: Absence of Previously Reported Correlation. Nucleosides Nucleotides Nucleic Acids 2004, 23, 1249–1251. [Google Scholar] [CrossRef]
- Camandola, S.; Plick, N.; Mattson, M.P. Impact of Coffee and Cacao Purine Metabolites on Neuroplasticity and Neurodegenerative Disease. Neurochem. Res. 2018, 44, 214–227. [Google Scholar] [CrossRef]
- Cova, I.; Leta, V.; Mariani, C.; Pantoni, L.; Pomati, S. Exploring cocoa properties: Is theobromine a cognitive modulator? Psychopharmacology 2019, 236, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Schreck, K.A.; Williams, K. Food preferences and factors influencing food selectivity for children with autism spectrum disorders. Res. Dev. Disabil. 2006, 27, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Diolordi, L.; Del Balzo, V.; Bernabei, P.; Vitiello, V.; Donini, L. Eating habits and dietary patterns in children with autism. Eat. Weight. Disord. Stud. Anorexia Bulim. Obes. 2014, 19, 295–301. [Google Scholar] [CrossRef]
- Yeldandi, A.V.; Yeldandi, V.; Kumar, S.; Murthy, C.N.; Wang, X.; Keith, A.; Rao, M.S.; Reddy, J.K. Molecular evolution of the urate oxidase-encoding gene in hominoid primates: Nonsense mutations. Gene 1991, 109, 281–284. [Google Scholar] [CrossRef]
- Yeldandi, A.; Patel, Y.; Liao, M.; Kao, F.; Rao, M.; Reddy, J.; Le Beau, M. Localization of the human urate oxidase gene (UOX) to 1p22. Cytogenet. Genome Res. 1992, 61, 121–122. [Google Scholar] [CrossRef]
- Page, T.; Coleman, M. Purine metabolism abnormalities in a hyperuricosuric subclass of autism. Biochim. Biophys. Acta Bioenerg. 2000, 1500, 291–296. [Google Scholar] [CrossRef]
- Ming, X.; Stein, T.P.; Barnes, V.; Rhodes, N.; Guo, L. Metabolic Perturbance in Autism Spectrum Disorders: A Metabolomics Study. J. Proteome Res. 2012, 11, 5856–5862. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, C. A metabolome study on 90 autism spectrum disorder patients by mass spectrometry. Med Mass Spectrom. 2017, 1, 14–19. [Google Scholar]
- Glinton, K.E.; Elsea, S.H. Untargeted Metabolomics for Autism Spectrum Disorders: Current Status and Future Directions. Front. Psychiatry 2019, 10, 647. [Google Scholar] [CrossRef]
- Sautin, Y.Y.; Johnson, R.J. Uric Acid: The Oxidant-Antioxidant Paradox. Nucleosides Nucleotides Nucleic Acids 2008, 27, 608–619. [Google Scholar] [CrossRef]
- Proctor, P.H. Uric acid: Neuroprotective or neurotoxic? Stroke 2008, 39, e88. [Google Scholar] [PubMed]
- Dawson, J.; Quinn, T.; Lees, K.; Walters, M. The continued yin and yang of uric acid. Stroke 2007, 39, 9. [Google Scholar] [CrossRef][Green Version]
- Giustarini, D.; Dalle-Donne, I.; Milzani, A.D.G.; Rossi, R. Oxidative stress induces a reversible flux of cysteine from tissues to blood in vivo in the rat. FEBS J. 2009, 276, 4946–4958. [Google Scholar] [CrossRef] [PubMed]
- Bridges, R.J.; Natale, N.R.; Patel, S.A. System xc- cystine/glutamate antiporter: An update on molecular pharmacology and roles within the CNS. Br. J. Pharmacol. 2011, 165, 20–34. [Google Scholar] [CrossRef]
- Lewerenz, J.; Hewett, S.J.; Huang, Y.; Lambros, M.; Gout, P.W.; Kalivas, P.W.; Massie, A.; Smolders, I.; Methner, A.; Pergande, M.; et al. The cystine/glutamate antiporter system x(c)(-) in health and disease: From molecular mechanisms to novel therapeutic opportunities. Antioxid. Redox Signal. 2013, 1, 522–525. [Google Scholar]
- Ghasemitarei, M.; Yusupov, M.; Razzokov, J.; Shokri, B.; Bogaerts, A. Effect of oxidative stress on cystine transportation by xC− antiporter. Arch. Biochem. Biophys. 2019, 674, 108114. [Google Scholar] [CrossRef]
- Castora, F.J. Mitochondrial function and abnormalities implicated in the pathogenesis of ASD. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2019, 92, 83–108. [Google Scholar] [CrossRef]
- McBean, G.J. Cysteine, Glutathione, and Thiol Redox Balance in Astrocytes. Antioxidants 2017, 6, 62. [Google Scholar] [CrossRef]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef]
- Seaquist, E.R.; Gruetter, R. Identification of a high concentration ofscyllo-inositol in the brain of a healthy human subject using1H- and13C-NMR. Magn. Reson. Med. 1998, 39, 313–316. [Google Scholar] [CrossRef]
- Michaelis, T.; Helms, G.; Merboldt, K.-D.; Hänicke, W.; Bruhn, H.; Frahm, J. Identification ofscyllo-inositol in proton NMR spectra of human brainin vivo. NMR Biomed. 1993, 6, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Nozadze, M.; Mikautadze, E.; Lepsveridze, E.; Mikeladze, E.; Kuchiashvili, N.; Kiguradze, T.; Kikvidze, M.; Solomonia, R. Anticonvulsant activities of myo-inositol and scyllo-inositol on pentylenetetrazol induced seizures. Seizure 2011, 20, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Ceppa, F.; Mancini, A.; Tuohy, K. Current evidence linking diet to gut microbiota and brain development and function. Int. J. Food Sci. Nutr. 2018, 70, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pero, R.W.; Lund, H. Dietary quinic acid supplied as the nutritional supplement AIO + AC-11® leads to induction of micromolar levels of nicotinamide and tryptophan in the urine. Phytother. Res. 2011, 25, 851–857. [Google Scholar]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef]
- Raimondi, S.; Anighoro, A.; Quartieri, A.; Amaretti, A.; Tomás-Barberán, F.A.; Rastelli, G.; Rossi, M. Role of bifidobacteria in the hydrolysis of chlorogenic acid. Microbiologyopen 2015, 4, 41–52. [Google Scholar] [CrossRef]
- Pero, R.W.; Lund, H.; Leanderson, T. Antioxidant metabolism induced by quinic acid. increased urinary excretion of tryptophan and nicotinamide. Phytother. Res. 2009, 23, 335–346. [Google Scholar] [CrossRef]
- Gao, K.; Mu, C.-L.; Farzi, A.; Zhu, W.-Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef]
- Savitz, J. The kynurenine pathway: A finger in every pie. Mol. Psychiatry 2020, 25, 131–147. [Google Scholar] [CrossRef]
- Guillemin, G.J. Quinolinic acid, the inescapable neurotoxin. FEBS J. 2012, 279, 1356–1365. [Google Scholar] [CrossRef]
- Wirthgen, E.; Hoeflich, A.; Rebl, A.; Günther, J. Kynurenic Acid: The Janus-Faced Role of an Immunomodulatory Tryptophan Metabolite and Its Link to Pathological Conditions. Front. Immunol. 2018, 8, 8. [Google Scholar] [CrossRef]
- Mandić, A.D.; Woting, A.; Jaenicke, T.; Sander, A.; Sabrowski, W.; Rolle-Kampcyk, U.; Von Bergen, M.; Blaut, M. Clostridium ramosum regulates enterochromaffin cell development and serotonin release. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous Bacteria from the Gut Microbiota Regulate Host Serotonin Biosynthesis. Cell 2015, 163, 258. [Google Scholar] [CrossRef]
- O’Mahony, S.; Clarke, G.; Borre, Y.; Dinan, T.; Cryan, J. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Muller, C.L.; Anacker, A.M.; Veenstra-VanderWeele, J. The serotonin system in autism spectrum disorder: From biomarker to animal models. Neuroscience 2016, 321, 24–41. [Google Scholar] [CrossRef]
- Myint, T.; Fraser, G.E.; Lindsted, K.D.; Knutsen, S.F.; Hubbard, R.W.; Bennett, H.W. Urinary 1-Methylhistidine Is a Marker of Meat Consumption in Black and in White California Seventh-day Adventists. Am. J. Epidemiol. 2000, 152, 752–755. [Google Scholar] [CrossRef]
- Liu, Y.-W.; Liong, M.-T.; Chung, Y.-C.E.; Huang, H.-Y.; Peng, W.-S.; Cheng, Y.-F.; Lin, Y.-S.; Wu, Y.-Y.; Tsai, Y.-C. Effects of Lactobacillus plantarum PS128 on Children with Autism Spectrum Disorder in Taiwan: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2019, 11, 820. [Google Scholar] [CrossRef]
- Buzzini, P.; Romano, S.; Turchetti, B.; Vaughan, A.; Pagnoni, U.M.; Davoli, P. Production of volatile organic sulfur compounds (VOSCs) by basidiomycetous yeasts. FEMS Yeast Res. 2005, 5, 379–385. [Google Scholar] [CrossRef]
- Sourabié, A.M.; Spinnler, H.-E.; Bourdat-Deschamps, M.; Tallon, R.; Landaud, S.; Bonnarme, P. S-methyl thioesters are produced from fatty acids and branched-chain amino acids by brevibacteria: Focus on l-leucine catabolic pathway and identification of acyl–CoA intermediates. Appl. Microbiol. Biotechnol. 2011, 93, 1673–1683. [Google Scholar] [CrossRef]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal Microbiota and Metabolome of Children with Autism and Pervasive Developmental Disorder Not Otherwise Specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef]
- Coretti, L.; Paparo, L.; Riccio, M.P.; Amato, F.; Cuomo, M.; Natale, A.; Borrelli, L.; Corrado, G.; De Caro, C.; Comegna, M.; et al. Gut Microbiota Features in Young Children With Autism Spectrum Disorders. Front. Microbiol. 2018, 9, 3146. [Google Scholar] [CrossRef]
- Fattorusso, A.; Di Genova, L.; Dell’Isola, G.B.; Mencaroni, E.; Esposito, S. Autism Spectrum Disorders and the Gut Microbiota. Nutrients 2019, 11, 521. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nat. Cell Biol. 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Gotham, K.O.; Pickles, A.; Lord, C. Trajectories of Autism Severity in Children Using Standardized ADOS Scores. Pediatrics 2012, 130, e1278–e1284. [Google Scholar] [CrossRef] [PubMed]
- Elsden, S.R.; Hilton, M.G.; Waller, J.M. The end products of the metabolism of aromatic amino acids by clostridia. Arch. Microbiol. 1976, 107, 283–288. [Google Scholar] [CrossRef]
- Schweiger, G.; Buckel, W. Identification of acrylate, the product of the dehydration of (R)-lactate catalyzed by cell-free extracts from Clostridium propionicum. FEBS Lett. 1985, 185, 253–256. [Google Scholar]
- Shaw, W. Increased urinary excretion of a 3-(3-hydroxyphenyl)-3-hydroxypropionic acid (HPHPA), an abnormal phenylalanine metabolite ofClostridiaspp. in the gastrointestinal tract, in urine samples from patients with autism and schizophrenia. Nutr. Neurosci. 2010, 13, 135–143. [Google Scholar] [CrossRef]
- Selmer, T.; Andrei, P.I. p-Hydroxyphenylacetate Decarboxylase from Clostridium Difficile. A Novel Glycyl Radical Enzyme Catalysing the Formation of P-Cresol. Eur. J. Biochem. 2001, 268, 1363–1372. [Google Scholar]
- Persico, A.M.; Napolioni, V. Urinary p-cresol in autism spectrum disorder. Neurotoxicol. Teratol. 2013, 36, 82–90. [Google Scholar] [CrossRef]
- Gabriele, S.; Sacco, R.; Cerullo, S.; Neri, C.; Urbani, A.; Tripi, G.; Malvy, J.; Barthelemy, C.; Bonnet-Brihault, F.; Persico, A.M. Urinary p-cresol is elevated in young French children with autism spectrum disorder: A replication study. Biomarkers 2014, 19, 463–470. [Google Scholar]
- Pascucci, T.; Colamartino, M.; Fiori, E.; Sacco, R.; Coviello, A.; Ventura, R.; Puglisi-Allegra, S.; Turriziani, L.; Persico, A.M. P-cresol Alters Brain Dopamine Metabolism and Exacerbates Autism-Like Behaviors in the BTBR Mouse. Brain Sci. 2020, 10, 233. [Google Scholar] [CrossRef]
- Gabriele, S.; Sacco, R.; Altieri, L.; Neri, C.; Urbani, A.; Bravaccio, C.; Riccio, M.P.; Iovene, M.R.; Bombace, F.; De Magistris, L.; et al. Slow intestinal transit contributes to elevate urinaryp-cresol level in Italian autistic children. Autism Res. 2015, 9, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Yuille, S.; Reichardt, N.; Panda, S.; Dunbar, H.; Mulder, I.E. Human gut bacteria as potent class I histone deacetylase inhibitors in vitro through production of butyric acid and valeric acid. PLoS ONE 2018, 13, e0201073. [Google Scholar] [CrossRef]
- Puig-Alcaraz, C.; Fuentes-Albero, M.; Cauli, O. Relationship between adipic acid concentration and the core symptoms of autism spectrum disorders. Psychiatry Res. 2016, 242, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Clark-Taylor, T.; Clark-Taylor, B.E. Is autism a disorder of fatty acid metabolism? Possible dysfunction of mitochondrial beta-oxidation by long chain acyl-CoA dehydrogenase. Med. Hypotheses 2004, 62, 970–975. [Google Scholar] [PubMed]
- Wu, J.-Y.; Roberts, E. Properties of Brain L-glutamate decarboxylase: Inhibition Studies. J. Neurochem. 1974, 23, 759–767. [Google Scholar] [CrossRef]
- Schousboe, A.; Wu, J.-Y.; Roberts, E. Subunit structure and kinetic properties of 4-aminobutyrate-2-ketoglutarate transaminase purified from mouse brain. J. Neurochem. 1974, 23, 1189–1195. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, D.; Wang, F.; Liu, S.; Zhao, S.; Ling, E.A.; Hao, A. Saturated fatty acids activate microglia via Toll-like receptor 4/NFkappaB signalling. Br. J. Nutr. 2012, 107, 229–241. [Google Scholar]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef]
- Fatima, S.; Hu, X.; Gong, R.-H.; Huang, C.; Chen, M.; Wong, H.L.X.; Bian, Z.; Kwan, H.Y. Palmitic acid is an intracellular signaling molecule involved in disease development. Cell. Mol. Life Sci. 2019, 76, 2547–2557. [Google Scholar] [CrossRef]
- Jung, S.; Choi, M.; Choi, K.; Bin Kwon, E.; Kang, M.; Kim, D.-E.; Jeong, H.; Kim, J.; Kim, J.H.; Kim, M.O.; et al. Inactivation of human DGAT2 by oxidative stress on cysteine residues. PLoS ONE 2017, 12, e0181076. [Google Scholar] [CrossRef]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef] [PubMed]
- Kordowska-Wiater, M. Production of arabitol by yeasts: Current status and future prospects. J. Appl. Microbiol. 2015, 119, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Kałużna-Czaplińska, J.; Błaszczyk, S. The level of arabinitol in autistic children after probiotic therapy. Nutrition 2012, 28, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Christensson, B.; Sigmundsdottir, G.; Larsson, L. D-arabinitol—A marker for invasive candidiasis. Med. Mycol. 1999, 37, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, P.; Del Chierico, F.; Putignani, L. Gut Microbiota Profiling: Metabolomics Based Approach to Unravel Compounds Affecting Human Health. Front. Microbiol. 2016, 7, 1144. [Google Scholar] [CrossRef]
- Larsen, P.E.; Dai, Y. Metabolome of human gut microbiome is predictive of host dysbiosis. GigaScience 2015, 4, 42. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; (DSM-5); American Psychiatric Association: Washington, DC, USA, 2013; p. 5. [Google Scholar]
- Lord, C.; Rutter, M.; DiLavore, P.C.; Risi, S.; Gotham, K.; Bishop, S. Autism Diagnostic Observation Schedule, 2nd ed.; (ADOS-2) Manual (Part I): Modules 1-4; WPS Publishing, Western Psychological Services: Torrance, CA, USA, 2012. [Google Scholar]
- Hus, V.; Gotham, K.; Lord, C. Standardizing ADOS Domain Scores: Separating Severity of Social Affect and Restricted and Repetitive Behaviors. J. Autism Dev. Disord. 2014, 44, 2400–2412. [Google Scholar] [CrossRef]
- Fulceri, F.; Narzisi, A.; Apicella, F.; Balboni, G.; Baldini, S.; Brocchini, J.; Domenici, I.; Cerullo, S.; Igliozzi, R.; Cosenza, A.; et al. Application of the Repetitive Behavior Scale-Revised—Italian version—In preschoolers with autism spectrum disorder. Res. Dev. Disabil. 2016, 48, 43–52. [Google Scholar] [CrossRef]
- Aman, M.; Singh, N. (Eds.) The Aberrant Behavior Checklist—Community. Supplementary Manual; Slosson Education Publications, Inc.: New York, NY, USA, 1994. [Google Scholar]
- Aman, M.; Singh, N. (Eds.) The Aberrant Behavior Checklist Manual, 2nd ed.; Slosson Education Publications, Inc.: New York, NY, USA, 2017. [Google Scholar]
- Lam, K.S.; Aman, M.G. The Repetitive Behavior Scale-Revised: Independent Validation in Individuals with Autism Spectrum Disorders. J. Autism Dev. Disord. 2007, 37, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Palmas, F.; Mussap, M.; Fattuoni, C. Urine metabolome analysis by gas chromatography–mass spectrometry (GC–MS): Standardization and optimization of protocols for urea removal and short-term sample storage. Clin. Chim. Acta 2018, 485, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Oberacher, H.; Whitley, G.; Berger, B. Evaluation of the sensitivity of the ‘Wiley registry of tandem mass spectral data, MSforID’ with MS/MS data of the ‘NIST/NIH/EPA mass spectral library’. J. Mass Spectrom. 2013, 48, 487–496. [Google Scholar] [CrossRef]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. GMD@CSB.DB: The Golm Metabolome Database. Bioinformatics 2004, 21, 1635–1638. [Google Scholar] [CrossRef]
- Dudzik, D.; Barbas-Bernardos, C.; Garcia, A.; Barbas, C. Quality assurance procedures for mass spectrometry untargeted metabolomics. a review. J. Pharm. Biomed. Anal. 2018, 147, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Beger, R.D.; Dunn, W.B.; Bandukwala, A.; Bethan, B.; Broadhurst, D.; Clish, C.B.; Dasari, S.; Derr, L.; Evans, A.; Fischer, S.; et al. Towards quality assurance and quality control in untargeted metabolomics studies. Metabolomics 2019, 15, 1–5. [Google Scholar] [CrossRef]
- Broadhurst, D.; Goodacre, R.; Reinke, S.N.; Kuligowski, J.; Wilson, I.D.; Lewis, M.R.; Dunn, W.B. Guidelines and considerations for the use of system suitability and quality control samples in mass spectrometry assays applied in untargeted clinical metabolomic studies. Metabolomics 2018, 14, 72. [Google Scholar] [CrossRef]
- Edmands, W.M.B.; Ferrari, P.; Scalbert, A. Normalization to Specific Gravity Prior to Analysis Improves Information Recovery from High Resolution Mass Spectrometry Metabolomic Profiles of Human Urine. Anal. Chem. 2014, 86, 10925–10931. [Google Scholar] [CrossRef]
- Whiteley, P.; Waring, R.; Williams, L.; Klovrza, L.; Nolan, F.; Smith, S.; Farrow, M.; Dodou, K.; Lough, W.J.; Shattock, P. Spot urinary creatinine excretion in pervasive developmental disorders. Pediatr. Int. 2006, 48, 292–297. [Google Scholar] [CrossRef]
- Misra, B. Data normalization strategies in metabolomics: Current challenges, approaches, and tools. Eur. J. Mass Spectrom. 2020, 26, 165–174. [Google Scholar] [CrossRef]
- Berg, R.A.V.D.; Hoefsloot, H.C.J.; Westerhuis, J.A.; Smilde, A.K.; Van Der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 1–15. [Google Scholar] [CrossRef]
- Triba, M.N.; Le Moyec, L.; Amathieu, R.; Goossens, C.; Bouchemal, N.; Nahon, P.; Rutledge, D.N.; Savarin, P. PLS/OPLS models in metabolomics: The impact of permutation of dataset rows on the K-fold cross-validation quality parameters. Mol. BioSyst. 2015, 11, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Akarachantachote, N.; Chadcham, S.; Saithanu, K. Cutoff threshold of variable importance in projection for variable selection. Int. J. Pure Appl. Math. 2014, 94, 307–322. [Google Scholar] [CrossRef]
- Ideker, T.; Krogan, N.J. Differential network biology. Mol. Syst. Biol. 2012, 8, 565. [Google Scholar] [CrossRef]
- Barabási, A.L.; Oltvai, Z.N. Network biology: Understanding the cell’s functional organization. Nat. Rev. Genet. 2004, 5, 101–113. [Google Scholar]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef]
- Grapov, D.; Wanichthanarak, K.; Fiehn, O. MetaMapR: Pathway independent metabolomic network analysis incorporating unknowns. Bioinformatics 2015, 31, 2757–2760. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–245. [Google Scholar]
- Wishart, D. 256. Note: An Algorithm for Hierarchical Classifications. Biometrics 1969, 25, 165. [Google Scholar] [CrossRef]
- Carbone, P.S.; Campbell, K.; Wilkes, J.; Stoddard, G.J.; Huynh, K.; Young, P.C.; Gabrielsen, T.P. Primary Care Autism Screening and Later Autism Diagnosis. Pediatrics 2020, 146, e20192314. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Median (IQR) 1 | Range |
|---|---|---|
| Age (y) | 5 (3–6) | 2–11 |
| Gender (M/F) | 23/8 | === |
| Family type (n simplex/n multiplex) | 25/6 | === |
| Gastrointestinal disease (n GI/n total) | 6/31 | === |
| Food selectivity (n FS/n total) | 17/31 | === |
| Developmental Level | 50 (43–60) | 21–84 |
| ADOS-2 CSS 2 | 7.5 (6–9.2) | 3–10 |
| SCQ score 3 | 15 (11–22) | 2–42 |
| SRS score 4 | 78 (70–90) | 50–101 |
| RBS-R score 5 | 23 (8.5–35) | 2–94 |
| ABC-C score 6 | 40 (23–55) | 4–128 |
| Clinical Assessment | Score | ||
|---|---|---|---|
| Result | Median (IQR) 1 | Range | |
| Age (y) | 5.8 | 5 (3–6) | 2–11 |
| Gastrointestinal disease | No | === | === |
| Food selectivity | Yes | === | === |
| Developmental level | 43 | 50 (43–60) | 21–84 |
| ADOS-2 CSS 2 | 6 | 7.5 (6–9.2) | 3–10 |
| SCQ 3 | 11 | 15 (11–22) | 2–42 |
| RSR 4 | 71 | 78 (70–90) | 50–101 |
| RBS-R 5 | 15 | 23 (8.5–35) | 2–94 |
| ABC-C 6 | 18 | 40 (23–55) | 4–128 |
| Metabolite | Two-Tailed Mann Whitney U test | VIP * | % Difference ASD vs. NT | |
|---|---|---|---|---|
| p | z-Score | |||
| 7-Methylxanthine | 0.012 | 2.48 | 3.30 | −61% |
| Scylloinositol | 0.011 | 2.52 | 2.43 | −35% |
| Uric acid | 0.002 | −3.02 | 2.41 | −50% |
| Aminomalonic acid | 0.034 | 2.10 | 1.73 | −52% |
| Quinic acid | 0.002 | 3.11 | 1.72 | +263% |
| Hippuric acid | 0.003 | −2.93 | 1.65 | +164% |
| Tryptophan | 0.024 | 2.25 | 1.44 | +100% |
| 1-Methylhistidine | 0.015 | −2.42 | 1.41 | +67% |
| Cystine | 0.018 | 2.36 | 1.37 | +101% |
| Indole-3-acetic acid | 0.036 | 2.10 | 1.20 | +61% |
| Allyl thioacetic acid | 0.014 | −2.46 | 1.12 | +28% |
| Leucine | 0.006 | 2.76 | 0.93 | +49% |
| Lactic acid | 0.004 | −2.87 | 0.88 | +67% |
| Metabolite | Two-Tailed Mann Whitney U Test | VIP * | % Difference ASD vs. NT | |
|---|---|---|---|---|
| p | z-Score | |||
| 2-Hydroxyacrylic acid | 0.002 | 3.09 | 2.27 | +321% |
| Sucrose | 0.009 | 2.58 | 1.89 | +234% |
| Allantoin | 0.003 | 3.00 | 1.85 | +143% |
| 3-Methylhistidine | 0.039 | 2.06 | 1.84 | +726% |
| Adipic acid | 0.005 | 2.83 | 1.80 | +389% |
| 3-(3-Hydroxyphenyl)-3-hydroxypropanoic acid | 0.008 | 2.64 | 1.74 | +142% |
| Xylose | 0.002 | 3.06 | 1.69 | +364% |
| 1-Deoxypentitol | 0.0002 | −3.68 | 1.68 | +294% |
| Glyceric acid | 0.016 | 2.41 | 1.65 | +265% |
| Palmitic acid | 0.005 | 2.80 | 1.63 | +263% |
| Hippuric acid | 0.044 | 2.00 | 1.60 | +196% |
| Homovanillic acid | 0.011 | 2.54 | 1.58 | +204% |
| 5-Hydroxyindoleacetic acid | 0.011 | 2.54 | 1.48 | +131% |
| Ribitol | 0.003 | 3.00 | 1.47 | +156% |
| Benzoic acid | 0.84 * | 1.73 * | 1.42 | +266% |
| Proline | 0.003 | 3.00 | 1.41 | +148% |
| p-Cresol | 0.048 | −1.97 | 1.40 | +105% |
| Quinolinic acid | 0.004 | 2.84 | 1.39 | +262% |
| Lactic acid | 0.009 | 2.59 | 1.26 | +88% |
| Oxalic acid | 0.035 | 2.11 | 1.22 | +138% |
| Mannose | 0.010 | 2.56 | 1.12 | +204% |
| Trihydroxypentanoic acid | 0.007 | 2.68 | 1.08 | +96% |
| Screening | Candidate Cut-Off Level | R2 X | R2 Y | Q2 | P |
|---|---|---|---|---|---|
| RBS-R | >35 | 0.496 | 0.933 | 0.557 | 1.00 |
| ABC-C | >50 | 0.487 | 0.822 | 0.465 | 0.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mussap, M.; Siracusano, M.; Noto, A.; Fattuoni, C.; Riccioni, A.; Rajula, H.S.R.; Fanos, V.; Curatolo, P.; Barberini, L.; Mazzone, L. The Urine Metabolome of Young Autistic Children Correlates with Their Clinical Profile Severity. Metabolites 2020, 10, 476. https://doi.org/10.3390/metabo10110476
Mussap M, Siracusano M, Noto A, Fattuoni C, Riccioni A, Rajula HSR, Fanos V, Curatolo P, Barberini L, Mazzone L. The Urine Metabolome of Young Autistic Children Correlates with Their Clinical Profile Severity. Metabolites. 2020; 10(11):476. https://doi.org/10.3390/metabo10110476
Chicago/Turabian StyleMussap, Michele, Martina Siracusano, Antonio Noto, Claudia Fattuoni, Assia Riccioni, Hema Sekhar Reddy Rajula, Vassilios Fanos, Paolo Curatolo, Luigi Barberini, and Luigi Mazzone. 2020. "The Urine Metabolome of Young Autistic Children Correlates with Their Clinical Profile Severity" Metabolites 10, no. 11: 476. https://doi.org/10.3390/metabo10110476
APA StyleMussap, M., Siracusano, M., Noto, A., Fattuoni, C., Riccioni, A., Rajula, H. S. R., Fanos, V., Curatolo, P., Barberini, L., & Mazzone, L. (2020). The Urine Metabolome of Young Autistic Children Correlates with Their Clinical Profile Severity. Metabolites, 10(11), 476. https://doi.org/10.3390/metabo10110476