Abstract
Benign prostatic hyperplasia (BPH) is a common disease in aging men. Panicum dichotomiflorum (PD) is an annual grass species of Poaceae that is distributed worldwide. The present study examined whether PD has a protective effect against BPH. BPH was generated in rats by daily subcutaneous administration of testosterone for four weeks. During this period, the rats were also given daily oral gavages of an extract of PD (150 mg/kg). After the final treatment, all animals were euthanized and their prostates were collected and weighed. In BPH model rats, the prostate weight and levels of dihydrotestosterone (DHT) and 5α-reductase expression were inhibited following treatment with PD extract. Testosterone-induced increases in prostate gland epithelial thickness and expression of cyclin D1 and proliferating cell nuclear antigen (PCNA) were markedly suppressed in PD-treated rats, whereas cleaved caspase-3 levels were increased. PD administration also decreased the expression of transforming growth factor (TGF)-β and vascular endothelial growth factor (VEGF), the phosphorylation of Akt, and inflammatory cytokines levels. Taken together, these results show that PD extract protects against testosterone-induced BPH progression by alleviating prostate cell growth and reducing levels of growth factors and inflammatory cytokines, indicating that PD extract may have potential in protecting against BPH.
1. Introduction
Benign prostatic hyperplasia (BPH) is one of the most frequent diseases in aging men. BPH is characterized by hyperplasia of smooth muscle and stromal cells in the transitional area of the prostate. This hyperplasia compresses the urethra and can cause lower urinary tract symptoms (LUTS) [1,2] such as nocturia, frequent urination, reduced urination flow, and feelings of residual urine, thereby impacting quality of life (QOL) [3]. Currently, there are no effective ways to prevent BPH.
Although the pathophysiology of BPH has not been fully elucidated, some mechanisms have been suggested [4]. Testosterone from the testis and adrenal gland plays a major role in prostate growth, and the prostatic 5α-reductase enzyme (especially type 2), which converts testosterone to dihydrotestosterone (DHT), has been shown to play an important role in BPH pathogenesis. DHT binds to androgen receptor (AR) with higher affinity than does testosterone [5]. AR is a type of nuclear receptor that binds with accessory proteins to function as a transcription factor, resulting in the transcription of target genes and ultimately in the synthesis of encoded proteins [5,6]. DHT also regulates the expression and receptor activation levels of certain growth factors, including transforming growth factor-beta1 (TGF-β1), vascular endothelial growth factor (VEGF), insulin-like growth factor 1 (IGF-1), and epidermal growth factor (EGF) [7]. These growth factors modulate the interaction between stromal and epithelial cells to alter cell proliferation and apoptosis, ultimately causing BPH progression [6]. The progression of BPH is closely related to inflammation, with several studies demonstrating the involvement of inflammatory cytokines, including interleukins (ILs) and tumor necrosis factor-α (TNF-α) [8]. Inflammation can cause proliferation within prostatic tissue through various mechanisms, including oxidative stress [9].
Although preventing the development of BPH is critical, there are no medication approved to prevent BPH. Current pharmacological therapies seek to decrease LUTS, reduce disease progression, and limit complications. The drugs approved for BPH include 5α-reductase inhibitors (5-ARIs), alpha1-adrenergic antagonists (α1-blockers), phosphodiesterase 5 inhibitors (PDE5Is), and their combined therapies such as the use of an α1-blocker with a 5-ARI [10]. The 5-ARIs inhibit the conversion of testosterone to DHT to reduce the level of DHT and thus inhibit prostate enlargement and BPH progression [11]. Although 5-ARI is currently used in the clinic, the agent has shown adverse effects on libido and erectile function [11,12]. The α1-blockers relax prostatic smooth muscle, thereby helping to alleviate LUTS without altering prostate weight [13]. PDE5Is relax the bladder detrusor and urinary smooth muscle to help alleviate LUTS via cyclic guanosine monophosphate (cGMP) [1]. In BPH patients, single-drug therapy often fails to result in satisfactory efficacy, and may have low patient compliance due to the risk of side effects and the slow onset of clinical symptom improvement. These have led researchers to investigate alternative methods with fewer side effects for treating BPH.
Panicum dichotomiflorum (PD), also known as fall panicum, is an annual grass species of Poaceae that is distributed worldwide in tropical and temperate regions [14]. The seeds of PD are edible, and the stems and leaves are used as a paper-making material as well as a cutting agent and animal feed component in South Korea [15]. A previous report described the repeated column chromatography-based isolation of four flavonoid derivatives and one lignan from the aerial structures of PD [16]. Spectroscopic analysis identified them as the flavonoid derivatives, tricin, luteolin, luteolin-4′-O-β-D-glucopyranoside and luteolin-7-O-β-D-glucopyranoside, and the lignan, (-)-pinoresinol [16]. Among the compounds isolated from PD, pinoresinol has been reported to exhibit multiple pharmacological activities, including neuroprotection, anti-inflammation, and anti-tumor activity [17,18,19]. Tricin was found to show anti-inflammatory effects in lipopolysaccharide-stimulated hPBMCs and carrageenan-induced rats and to exhibit potent antiangiogenic activity in vitro [20,21]. Luteolin has been reported to induce apoptosis and decrease angiogenesis and carcinogenesis in animal models, diminish tumor growth in vivo, and render tumor cells sensitive to some anticancer drugs; thus, this flavonoid appears to exhibit cancer chemopreventive activity, and thus may hold chemotherapeutic potential [22]. Moreover, luteolin and its derivative, luteolin-4′-O-glucoside, exhibited strong efficacy in treating hyperuricemia and gout in the clinic [23], and showed antioxidative potential in RAW 264.7 cells [24]. Studies on plants of the genus Panicum found that P. turgidum and P. repens contain the steroid, saponin, as well as phenolic compounds such as rutin, caffeic acid, and protocatechuic acid [25,26,27,28]. Bioactivity studies showed that P. miliare has antihyperlipidemic efficacy and that P. maximum has antidiabetic and anti-inflammatory effects [29,30,31]. However, the anti-BPH effects of these plants remain unknown. Here, we performed an exploratory study to examine whether an extract of PD protects against BHP in a rodent model of testosterone-induced BPH.
2. Materials and Methods
2.1. Preparation of PD Extract
The PD extract (leaf and stem) used in the present study was obtained from the Natural Product Central Bank at the Korea Research Institute of Bioscience and Biotechnology (Daejeon, Republic of Korea).
2.2. UPLC-QTOF/MS Analysis of PD Extract
Analysis of the PD extract was conducted using a Waters ACQUITY UPLC system equipped with a XEVO-G2 XS QTOF mass detector (Waters, Milford, MA, USA), which featured an electrospray ionization (ESI) source. The ESI source was operated in both positive and negative modes, with a capillary voltage of 2.3 kV and a cone voltage of 40 V. An ACQUITY UPLC BEH C18 Column (Waters) was used, with two mobile phases: A, 0.1% formic acid in water; and B, 0.1% formic acid in acetonitrile. A gradient elution method was employed, commencing with 5% B for 1 min, followed by a linear gradient from 5% to 100% B over the subsequent 19 min. The mobile phase was then held at 100% B for 2.5 min, followed by a rapid decrease to the initial condition of 5% B over 0.5 min, and a subsequent 2.5 min hold at 5% B for an equilibration period to stabilize the column. The flow rate was maintained at 0.4 mL/min, and a 1 μL injection volume was utilized. The PD extract was prepared at a concentration of 3 mg/mL in methanol. Data were recorded using a Waters MassLynx v4.2 SCN1033 in centroid mode.
2.3. Experimental Animals
Seven-week-old male SD rats (150–220 g; Orient Bio, Seongnam, Republic of Korea) were used. During the experimental period, the rats were given a standard rodent diet with access to water ad libitum. The Animal Experimental Ethics Committee of Chungnam National University approved the animal experiments, which followed ethical guidelines for experimental animal studies (Daejeon, Republic of Korea; Approval No. CNU-00711).
2.4. BPH Induction and Drug Administration
Rats were randomly distributed to the following 4 groups (6–8 animals per group): (1) the normal control (NC) group, which was administered oral gavage of PBS and subcutaneous (s.c.) injection of corn oil; (2) the BPH group, which was administered oral gavage of PBS and s.c. injection of testosterone (3 mg/kg; Tokyo Chemical Industry, Tokyo, Japan); (3) the finasteride group (Fina), which received oral gavage of finasteride (10 mg/kg, Sigma, St. Louis, MO, USA) and s.c. injection of testosterone. The Fina group was considered as positive control.; and (4) the PD group, which received oral gavage of PD extract (150 mg/kg) and s.c. injection of testosterone. All treatments were given daily for 4 weeks with administration volumes of 5 mL/kg for oral gavage (PBS, finasteride, or PD extract) and 3 mL/kg for s.c. injection (corn oil or testosterone). In the present study, testosterone was administrated without castration. The vehicle was corn oil or PBS for testosterone or finasteride/PD extract, respectively. The animals were fasted overnight for 12 h after the last treatment and euthanized with CO2 gas. Blood samples were drawn from the caudal vena cava. Serum was collected from whole blood by centrifugation. Prostates were carefully isolated and weighed. The percent inhibition of hyperplasia was calculated from the relative prostate weight, as follows: 100 − [(treated group − normal control group)/(positive control group − normal control group)] × 100. For each prostate, a tissue sample was obtained from the ventral lobe. One part of this sample was fixed in 10% formalin, and the remainder was cut into several pieces and immediately frozen at −70 °C.
2.5. Determination of DHT and Testosterone
The levels of androgen hormone in serum were examined using ELISA kits (Cat. No. E-EL-0031 and 55-TESMS-E01; ALPCO Diagnostics, Salem, NH, USA) and all testing procedures followed the manufacturer’s manual.
2.6. Histological Examination
Hematoxylin and eosin (H&E) staining was performed as described previously [32]. Epithelial thickness (µm) was measured in the ventral lobes of prostate glands as described previously [33]. For each prostate, 30 randomly selected regions were evaluated. The pathologist who selected regions for viewing was blinded to the group allocation.
2.7. Proliferative Cell Nuclear Antigen (PCNA) Staining
PCNA staining was used to examine proliferating cells. Five regions of ventral lobe from the prostate of each animal were randomly assigned for evaluation. The slides were incubated overnight with anti-PCNA (Cat. No. ab29; Abcam, Cambridge, UK) followed by incubation with a biotinylated secondary antibody. The results were visualized with diaminobenzidine (DAB; Vector Laboratories, Newark, CA, USA) staining and hematoxylin counter-staining. PCNA-positive cells were expressed as a percentage of observed cells on the field.
2.8. Western Blot
Prostatic tissue samples were homogenized in radioimmunoprecipitation assay (RIPA) buffer (Cell Signaling Technology, Danvers, MA, USA) with protease inhibitor (Roche, Basel, Switzerland) and phosphatase inhibitor (Roche). Proteins (15 µg each) were separated by SDS-PAGE. Gels were transferred to nitrocellulose membranes (350 mA) for 2 h and incubated overnight with antibodies as follows: anti-cyclin D1, anti-caspase-3, anti-phospho-Akt (p-Akt), anti-Akt, and anti-β-actin (Cat. No. 2922, 9662, 9271, 9272, and 4970; Cell Signaling Technology, Danvers, MA, USA), and anti-B-cell lymphoma 2 (Cat. No. sc-70411; Bcl-2; Santa Cruz, Dallas, TX, USA). The membranes were incubated at room temperature with an appropriate secondary antibody, and the results were detected using a chemiluminescence detection kit and quantified.
2.9. Quantitative Real-Time PCR (RT-qPCR)
Equal amounts (1.4 µg) of RNA were extracted from prostatic tissue using TRIzol reagent and were subjected to reverse transcription (RT) using a commercial RT kit (Toyobo, Osaka, Japan). RT-qPCR was performed using SYBR Green Master Mix (Thermo Scientific, Waltham, MA, USA) and the following primers: 5α-reductase type2, forward (f): 5′-ATTTGTGTGGCAGAGAGAGG-3′ and reverse (r): 5′-TTGATTGACTGCCTGGATGG-3′; vascular endothelial growth factor (VEGF), f: 5′-GAGGAAAGGGAAAGGGTCAAAA-3′ and r: 5′-CACAGTGAACGCTCCAGGATT-3′; transforming growth factor-beta1 (TGF-β1), f: 5′-AGGGCTCAACACCTGCAC-3′ and r: 5′-GGGCCCCAGACAGAAGTT-3′; TNF-α, f: 5′- GTCTGTGCCTCAGCCTCTTC-3′ and r: 5′-CCCATTTGGGAACTTCGCCT-3′; IL-6, f: 5′-TAGTCCTTCCTACCCCAACT -3′ and r: 5′-TTGGTCCTTAGCCACTCCTT-3′; IL-8, f: 5′-CATTAATATTTAACGATGTGGATGCGTTTCA-3′ and r: 5′-GCCTACCATCTTTAAACTGCACAAT-3′; and GAPDH, f: 5′-ACAGCAACAGGGTGGTGGAC-3′ and r: 5′-TTTGAGGGTGCAGCGAACTT-3′. RT-qPCR data were processed using an Applied Biosystems PCR System and the associated computer program (Applied Biosystems, Waltham, MA, USA), and are presented as fold-change relative to the endogenous normalized control (GAPDH), as determined using the 2−ΔΔCt method [34].
2.10. Statistical Analysis
Data were represented as means ± standard deviations (SDs). One-way ANOVA with Dunnett’s post hoc test was applied for multiple comparisons between the BPH group and other groups. A p-value < 0.05 was considered statistically significant. Statistical analyses were performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA) software.
3. Results
3.1. Analysis of PD
To explore the potential medicinal properties of PD extract from a chemical perspective, we conducted a phytochemical analysis of the extract using UPLC-UV-QTOF/MS. This analysis allowed us to tentatively identify the predominant components present in the PD extract, as depicted in Figure 1 and summarized in Table 1. In the UV chromatogram at 254 nm, we observed the presence of tetrahydroxyflavonoid-C-hexosyl-O-deoxyhexoside (1), which is an isoorientin-O-rhamnoside isomer, with remarkable prominence. Furthermore, in the MS chromatograms in both ionization modes, we noted the heightened visibility of dichotomin (2), a steroidal saponin.
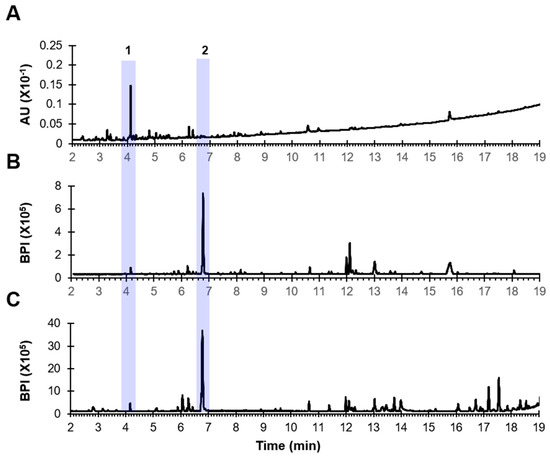
Figure 1.
UPLC-QTOF/MS chromatograms of the PD extract. (A) UV chromatogram at 254 nm. (B) MS chromatogram in negative mode. (C) MS chromatogram in positive mode.

Table 1.
Tentative identification of compounds using UPLC-QTOF/MS.
3.2. PD Extract Reduces Prostate Weight in BPH Rats
To examine the protective effect of PD extract against BPH, we assessed the prostate weight in testosterone-induced BPH rats. BPH rats exhibited marked increases in prostate weight (absolute and relative) compared to NC rats. The differences in weight were notably lower in the Fina and PD groups (Table 2). The percent inhibitions of prostate weight in the Fina and PD group animals were 63.98% and 30.28%, respectively (Table 2).
Table 2.
Effect of PD on prostatic weight (absolute and relative) and inhibitory action.
3.3. PD Extract Decreases Androgen and 5α-Reductase Levels in BPH Rats
Testosterone and DHT play important roles in the development of BPH [35]. The serum DHT concentration was much higher in the BPH group than in the NC group, and the serum DHT concentrations in the Fina and PD groups were lower than that in BPH animals (Figure 2A). The same pattern was seen for the prostate testosterone level (Figure 2B) and the relative expression of 5α-reductase (Figure 2C).
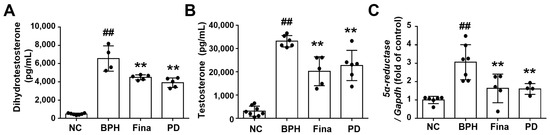
Figure 2.
Effects of PD extract on testosterone, DHT, and 5α-reductase. The serum levels of DHT (A) and testosterone (B) were examined by ELISA. (C) The relative mRNA levels of 5α-reductase in the prostate by RT-qPCR analysis. NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. ## p < 0.01 compared with the NC group; ** p < 0.01 compared with the BPH group.
3.4. PD Extract Decreases the Thickness of Glandular Epithelium in BPH Rats
Histopathological examination showed normal prostate morphology in NC animals, but revealed hyperplasia, increased glandular epithelial thickness, and reduced glandular luminal space in the prostates of BPH rats (Figure 3). The glandular epithelial thicknesses in the Fina and PD groups were lower than that of the BPH group and similar to that of the NC group (Figure 3).
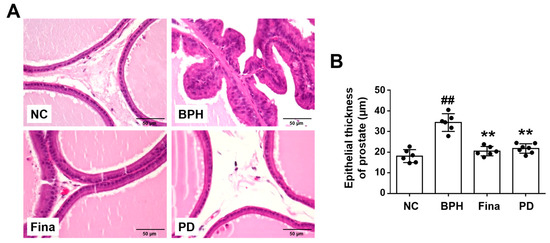
Figure 3.
Effects of PD extract on histological features in the prostate. (A) Representative images of H&E-stained prostatic tissue (×400). (B) Thickness of the glandular epithelium in the prostate. NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. ## p < 0.01 compared with the NC group; ** p < 0.01 compared with the BPH group.
3.5. PD Extract Ameliorates Testosterone-Induced Glandular Epithelial Cell Proliferation
The number of PCNA-positive cells was notably increased in BPH rats compared to NC rats, whereas the number of such cells was lower in the Fina and PD-administrated animals compared to NC rats (Figure 4A,B). Cyclin D1 protein expression was notably up-regulated in the BPH group, but this change was weaker in the Fina and PD groups (Figure 4C). These data indicate that PD extract attenuates BPH progression in part by exerting anti-proliferative activity.
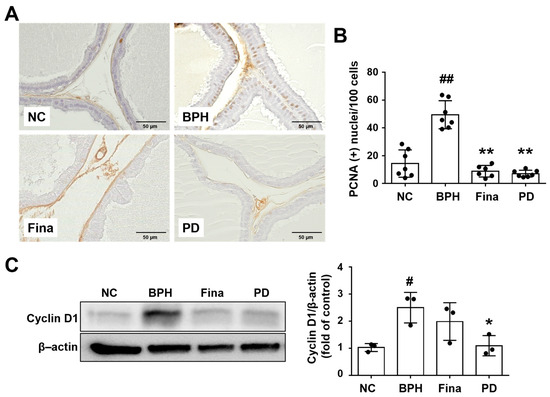
Figure 4.
Effects of PD extract on glandular epithelial cell proliferation. (A) Representative prostatic images of immunohistochemistry for PCNA (×400). (B) Percent of PCNA-stained cells in prostatic tissue (×400). (C) Cyclin D1 protein expression level was determined using Western blot analysis. NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. # p < 0.05 and ## p < 0.01 compared with the NC group; * p < 0.05 and ** p < 0.01 compared with the BPH group.
3.6. PD Extract Activates Apoptosis of Prostatic Tissue in BPH Rats
Given that a key aspect of BPH pathogenesis is an imbalance between cell proliferation and death [36], we used Western blot analysis to examine whether PD extract affects apoptotic proteins in prostatic tissues. Increased expression of Bcl-2 was observed in prostates of BPH rats, but this increase was not observed in rats in the Fina and PD groups (Figure 5A). In addition, the levels of the apoptosis marker, cleaved caspase-3, were higher in the Fina and PD groups than in the BPH group (Figure 5B).
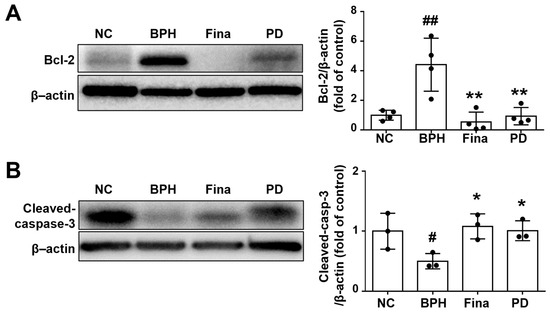
Figure 5.
Effects of PD extract on prostatic cell apoptosis. Expression levels of Bcl-2 (A) and cleaved caspase-3 (B) were evaluated by Western blot. NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. # p < 0.05 and ## p < 0.01 compared with the NC group; * p < 0.05 and ** p < 0.01 compared with the BPH group.
3.7. PD Extract Inhibits the Testosterone-Triggered Induction of Growth Factors and Akt Activation
In addition to directly affecting cell cycle regulation, DHT alters the expression of several growth factors [6,7]. Accordingly, we performed RT-qPCR to assess the mRNA expression of growth factors in prostatic tissues. The relative levels of TGF-β1 and VEGF were increased in the BPH group compared to the NC group, but these increases were diminished in the Fina and PD groups compared to the BPH group (Figure 6A,B).
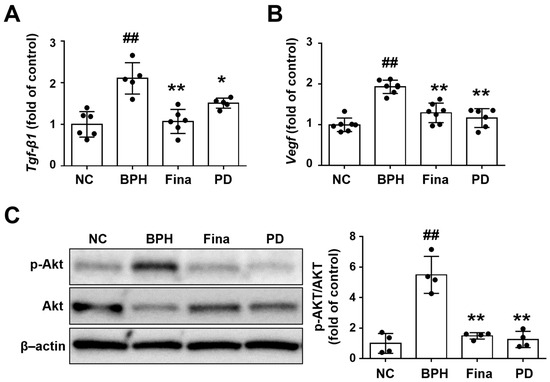
Figure 6.
Effects of PD extract on growth factors and Akt signaling in rat prostatic tissues. Relative mRNA levels of Tgf-β1 (A) and Vegf (B) determined by RT-qPCR in prostatic tissue. (C) Western blot analysis was used to examine the phosphorylation levels of phospho-Akt and Akt in lung tissue samples. NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. ## p < 0.01 compared with the NC group; * p < 0.05 and ** p < 0.01 compared with the BPH group.
The Akt pathway is generally considered to be important for growth factor signaling and cell cycle regulation [37,38]. Akt phosphorylation was markedly increased in BPH rats compared to NC rats, but this increase was only mild in the Fina and PD groups (Figure 6C).
3.8. PD Extract Suppresses Inflammatory Cytokines in BPH Rats
On histopathological examination, the NC group showed normal prostatic tissue morphology, the BPH group showed infiltration of inflammatory cells in prostatic interstitial lesions, and the Fina and PD groups showed only a few inflammatory cells (Figure 7A). Furthermore, the relative expression levels of TNF-α, IL-6, and IL-8 were increased in the BPH group, and the changes were notably reduced in the Fina and PD groups (Figure 7B).
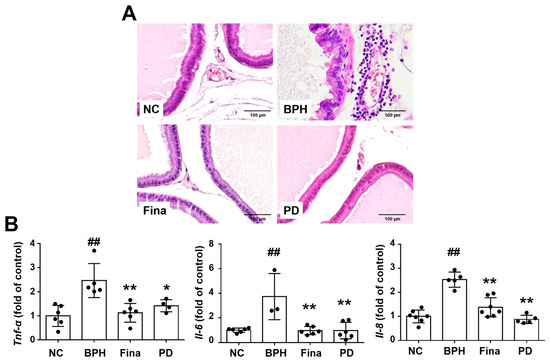
Figure 7.
Effects of PD extract on inflammation. (A) Representative prostatic images showing penetration of inflammatory cells in the BPH group (×200). (B) Measurement of mRNA expression of Tnf-α, Il-6, and Il-8 in the prostate by RT-qPCR. NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. ## p < 0.01 compared with the NC group; * p < 0.05 and ** p < 0.01 compared with the BPH group.
3.9. PD Extract Does Not Affect Liver Function or Histology
PD is the predominant grass species in hay and may cause hepatotoxicosis in horses and sheep [39,40]. To determine whether PD extract induced hepatotoxicity in our experimental setting, we examined the serum levels of aspartate transaminase (AST) and alanine aminotransferase (ALT) as well as liver weight. The relative liver weight and serum levels of AST and ALT did not differ between the NC group and any treatment group (Figure 8A–C).
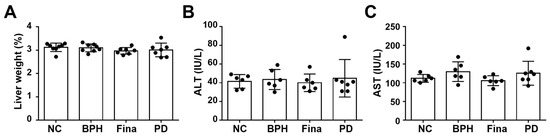
Figure 8.
Effects of PD extract on liver function and histology. Relative liver weights (A) and serum levels of ALT (B) and AST (C). NC, corn oil-injected and PBS-administrated rats; BPH, testosterone- and PBS-administrated rats; Fina, testosterone- and finasteride-administrated rats; PD, testosterone- and PD-administrated rats. There was no statistical difference between any group.
4. Discussion
In this study, we assessed whether PD extract has a protective effect against BPH in a rat model of testosterone-induced BPH. The testosterone-induced increases in relative prostate weight and serum DHT and prostatic 5α-reductase concentrations were decreased in PD-treated BPH rats. Moreover, treatment of BPH rats with PD extract notably suppressed the enhanced proliferation of prostatic glandular epithelial cells, ameliorated the increases in expression of growth factors and inflammatory cytokines, and activated apoptotic signaling. These results indicate that PD extract inhibits the progression of testosterone-induced BPH.
DHT, which is an androgen hormone related to prostate growth, is synthesized from testosterone with catalysis by 5α-reductase in the prostate [41], and has important functions in the pathogenesis and progression of BPH [35]. DHT levels in the prostate can be reduced by 5-ARI (e.g., finasteride), which is currently used for BPH treatment in the clinic [42]. Here, we showed that finasteride (positive control) decreased the serum level of testosterone and DHT. These findings are consistent with those of a previous study [43,44]. In addition, PD decreased the levels of testosterone and DHT in the serum and prostatic mRNA expression of 5α-reductase.
An imbalance between cell proliferation and apoptotic processes in the prostate is considered to be a critical component of BPH pathogenesis [36]. Apoptosis is an intrinsic cell death pathway that is thought to involve Bcl-2, related pro-survival factors, and familial pro-apoptotic proteins (e.g., Bax and Bak) [45]. The Bax/Bcl-2 ratio and caspase-3 activity level are considered to be molecular markers of apoptosis [46]. Bcl-2 inhibits apoptosis by supporting mitochondrial membrane integrity, and by preventing the oligomerization of Bax and Bak and subsequent mitochondrial release of apoptogenic molecules. Bcl-2 further inhibits apoptosis by binding to and inactivating other pro-apoptotic molecules, including Bax protein [47]. Meanwhile, Bax facilitates the release of mitochondrial cytochrome c and the downstream activation of caspase-3 [48], which consists of large and small subunits and cleaves substrates to enact cell death [49]. In the present study, we used PNCA staining to evaluate proliferation and Western blot experiments of apoptosis-related proteins to evaluate cell death. Consistent with previous reports, PD notably decreased the PCNA-positive cell number and down-regulated Bcl-2 expression while increasing the levels of cleaved caspase-3 in the prostate. These results indicated that PD extract evoked prostate epithelial cell apoptosis in the present BPH animal model to improve the imbalance between cell cycle progression and apoptosis. BPH is defined histologically as a disease process characterized by stromal and glandular epithelial cell hyperplasia of the prostate [50,51]. Although PD reduced the positive cell number of PCNA in glandular epithelial cells, further study is needed to determine whether PD treatment could decrease the proliferation of prostatic stromal cells.
Growth factors play critical roles in the interaction between stromal and epithelial cells, and alteration of this cross-talk contributes to the development of BPH [6]. Various growth factors are involved in BHP development, including TGF-β, VEGF, FGF, and IGF [52,53]. They are primarily secreted by stromal cells, and act in paracrine and autocrine manners to support the homeostasis of prostate cells. Among them, TGF-β1 was reported to induce fibroblasts to differentiate into myofibroblasts and promote stromal cell remodeling [54]. TGF-β1 is mainly expressed in epithelial cells, and it is present at higher levels in BPH than in normal prostatic tissue [54]. It may affect extracellular matrix (ECM) deposition and inflammatory signaling [55]. VEGF, which has been associated with angiogenesis [56], is secreted by epithelial cells in response to androgens, and contributes to the hyperplasic growth seen in BPH [57]. VEGF downstream signaling is critical to tumorigenesis, and may be induced by hypoxia inducible factor 1-a (HIF-1a) and signal transducer/activator of transcription-3 (STAT3) [37]. VEGF can activate cell survival and mitogenic pathways. Consistent with the above findings, we observed that the relative mRNA expression levels of TGF-β1 and VEGF were increased in the BPH group, and that treatment with PD extract reversed these effects.
The signaling pathways downstream of growth factors share the PI3K/Akt pathway as a common intracellular mediator [37]. The PI3K/Akt signaling cascade has been associated with proliferation, metabolism, growth, and migration; it is known to promote cell survival and apoptosis resistance via several mechanisms, including through the Bcl-2 family members, Bad and Bax [58]. Consistent with this, we herein found that Akt phosphorylation was increased in testosterone-induced BPH rats and that this effect was reversed by PD extract.
BPH progression is linked to the presence of an inflammatory environment. Several studies found that, in BPH patients, the prostate stroma produces various chemokines, the most prominent being CXC chemokines [59]. The inflammation associated with BPH is immunologically characterized by abundant T-cells and inflammatory cytokine expression [60]. Prostate inflammation involves inflammatory cell infiltration and localized tissue effects; neutrophil infiltration is seen under acute inflammation, whereas chronic inflammation involves T-cells. Localized tissue effects may be partly governed by Toll-like receptors (TLRs) on prostatic epithelial cells [61]. TLR activation stimulates the secretion of local inflammatory mediators, including IL-1, IL-6, and IL-8 [61], which recruit further inflammatory cells and stimulate local inflammatory responses [60]. Human BPH is characterized by general increases in pro-inflammatory interleukin expression. IL-6 is an acute-acting factor that is released by stromal and epithelial cells under acute inflammation. Epidemiologically, an elevated concentration of IL-6 has been reported in BPH patients younger than 65 years [62]. Consistent with this, prostates of BPH patients exhibit increased levels of IL-6 compared to those of normal individuals [63]. IL-8 is secreted by prostate epithelial cells; it contributes to leukocyte chemotaxis and is elevated in BPH tissue samples, especially in those of men with BPH and chronic prostatitis [64]. TNF-α is increased in the prostate tissue of BPH patients, potentially due to the inflammatory reaction caused by hyperplasia of prostate tissue and the destruction of tissue structure [65]. Consistent with this, we herein observed cell infiltration in the prostate and upregulation of TNF-α, IL-6, and IL-8 in BPH rats. However, these changes were reversed by treatment with PD extract.
In the present study, the predominant components of PD extract were differed from those in published reports [16]. These discrepancies may be due to differences in analysis conditions or the region of PD origin. In addition, the pharmacological molecules present in the PD extract and their underlying mechanisms were not determined in this study. Because several flavonoids from plant extract could leads to many artefacts, these should be elucidated in the further study. Additionally, we evaluated the protective effect of PD extract in a testosterone-induced BPH rat model. While rodent model is often used in BPH research and have some value, their general limitation is a difference in anatomy of the prostate between rodents and humans. Furthermore, anti-androgen targeting does not guarantee efficacy in the clinic. Further investigations are needed to determine the suitability of PD extract for clinical applications.
In sum, the present results demonstrate that treatment with PD extract markedly alleviates testosterone-induced prostate hyperplasia and histopathological changes in BPH rats. PD extract lowered the levels of serum DHT and prostate 5α-reductase, ameliorated the ability of testosterone to disrupt the balance between cell proliferation and apoptosis, and increased growth factor expression and inflammation. These results suggest that PD extract may have potential as a preventive treatment for BPH.
Author Contributions
Conceptualization: H.-J.K.; methodology: E.B.B. and E.-J.H.; formal analysis: E.B.B., E.-J.H., J.-H.K., M.K., and J.A.; investigation: E.B.B., and E.-J.H.: writing—original draft preparation: E.B.B.; writing—review and editing: E.B.B. and H.-J.K.; supervision: H.-J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the research fund of Chungnam National University.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of Chungnam National University (Approval No. CNU-00711) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data generated and analyzed in this study are mentioned in this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Roehrborn, C.G. Pathology of benign prostatic hyperplasia. Int. J. Impot. Res. 2008, 20 (Suppl. S3), S11–S18. [Google Scholar] [CrossRef]
- Foo, K.T. Pathophysiology of clinical benign prostatic hyperplasia. Asian J. Urol. 2017, 4, 152–157. [Google Scholar] [CrossRef]
- Zhang, A.Y.; Xu, X. Prevalence, Burden, and Treatment of Lower Urinary Tract Symptoms in Men Aged 50 and Older: A Systematic Review of the Literature. SAGE Open Nurs. 2018, 4, 2377960818811773. [Google Scholar] [CrossRef] [PubMed]
- Madersbacher, S.; Sampson, N.; Culig, Z. Pathophysiology of Benign Prostatic Hyperplasia and Benign Prostatic Enlargement: A Mini-Review. Gerontology 2019, 65, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Smock, S.L.; Castleberry, T.A.; Owen, T.A. Molecular cloning and functional characterization of the canine androgen receptor. Mol. Cell Biochem. 2001, 226, 129–140. [Google Scholar] [CrossRef] [PubMed]
- La Vignera, S.; Condorelli, R.A.; Russo, G.I.; Morgia, G.; Calogero, A.E. Endocrine control of benign prostatic hyperplasia. Andrology 2016, 4, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Carson, C., 3rd; Rittmaster, R. The role of dihydrotestosterone in benign prostatic hyperplasia. Urology 2003, 61, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Chughtai, B.; Lee, R.; Te, A.; Kaplan, S. Role of inflammation in benign prostatic hyperplasia. Rev. Urol. 2011, 13, 147–150. [Google Scholar] [PubMed]
- Naber, K.G.; Weidner, W. Chronic prostatitis-an infectious disease? J. Antimicrob. Chemother. 2000, 46, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Larson, J.A.; Andriole, G.L. Management of Benign Prostatic Hyperplasia. Annu. Rev. Med. 2016, 67, 137–151. [Google Scholar] [CrossRef]
- Drobnis, E.Z.; Nangia, A.K. 5alpha-Reductase Inhibitors (5ARIs) and Male Reproduction. Adv. Exp. Med. Biol. 2017, 1034, 59–61. [Google Scholar] [CrossRef]
- Tarter, T.H.; Vaughan, E.D., Jr. Inhibitors of 5alpha-reductase in the treatment of benign prostatic hyperplasia. Curr. Pharm. Des. 2006, 12, 775–783. [Google Scholar] [CrossRef]
- Lowe, F. Alpha-1-adrenoceptor blockade in the treatment of benign prostatic hyperplasia. Prostate Cancer Prostatic. Dis. 1999, 2, 110–119. [Google Scholar] [CrossRef]
- Burrows, G.E.; Tyrl, R.J. Toxic Plants of North America, 2nd ed.; Wiley-Blackwell: Hoboken, HJ, USA, 2013. [Google Scholar]
- Yoon, S.Y. Korean Resources Plant Encyclopedia; Academic Book: Seoul, Republic of Korea, 1995. [Google Scholar]
- Bae, J.J. Phytochemical Constituents of Panicum dichotomiflorum Michaux. Korean J. Pharmacogn. 2017, 48, 285–288. [Google Scholar]
- In, S.J.; Seo, K.H.; Song, N.Y.; Lee, D.S.; Kim, Y.C.; Baek, N.I. Lignans and neolignans from the stems of Vibrunum erosum and their neuroprotective and anti-inflammatory activity. Arch. Pharm. Res. 2015, 38, 26–34. [Google Scholar] [CrossRef]
- Ahmad, I.; Waheed, A.; Tahir, N.B.; Rais, A.K. Anti-inflammatory constituents from Perovskia atriplicifolia. Pharm. Biol. 2015, 53, 1628–1631. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Biedma, A.; Sanchez-Quesada, C.; Beltran, G.; Delgado-Rodriguez, M.; Gaforio, J.J. Phytoestrogen (+)-pinoresinol exerts antitumor activity in breast cancer cells with different oestrogen receptor statuses. BMC Complement. Altern. Med. 2016, 16, 350. [Google Scholar] [CrossRef] [PubMed]
- Shalini, V.; Jayalekshmi, A.; Helen, A. Mechanism of anti-inflammatory effect of tricin, a flavonoid isolated from Njavara rice bran in LPS induced hPBMCs and carrageenan induced rats. Mol. Immunol. 2015, 66, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Kwon, H.J.; Jung, H.J. Tricin, 4’,5,7-trihydroxy-3’,5’-dimethoxyflavone, exhibits potent antiangiogenic activity in vitro. Int. J. Oncol. 2016, 49, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, P.G.; Liang, W.Q.; Hu, Y.J.; Xu, P.; Zhou, J.; Pu, J.B.; Zhang, H.J. Luteolin-4’-O-glucoside and its aglycone, two major flavones of Gnaphalium affine D. Don, resist hyperuricemia and acute gouty arthritis activity in animal models. Phytomedicine 2018, 41, 54–61. [Google Scholar] [CrossRef]
- Song, Y.S.; Park, C.M. Luteolin and luteolin-7-O-glucoside strengthen antioxidative potential through the modulation of Nrf2/MAPK mediated HO-1 signaling cascade in RAW 264.7 cells. Food Chem. Toxicol. 2014, 65, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.A.; Ali, Z.; Wang, Y.H.; El-Amier, Y.A.; Khan, S.I.; Khan, I.A. Cytotoxic steroidal saponins from Panicum turgidum Forssk. Steroids 2017, 125, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Temraz, A.; Hozaien, H.E.; El-Tantawy, W.H.; El-Gindi, O.D.; Taha, K.F. Cholestane and spirostane-type glycosides from the roots and rhizomes of Panicum repens L. Phytochem. Lett. 2014, 10, 173–178. [Google Scholar] [CrossRef]
- Farag, M.A.; El Fishawy, A.M.; El-Toumy, S.A.; Amer, K.F.; Mansour, A.M.; Taha, H.E. Antihepatotoxic Effect and Metabolite Profiling of Panicum turgidum Extract via UPLC-qTOF-MS. Pharmacogn. Mag. 2016, 12, S446–S453. [Google Scholar] [CrossRef] [PubMed]
- Zakia, A.A.; Qiub, L.; Alib, Z.; Khanb, S.I.; Khanb, I.A. Anti-inflammatory Steroidal Saponins from Panicum turgidum. J. Agric. Basic Sci. 2016, 1, 1–6. [Google Scholar]
- Radha, R.; Vijayalakshmi, P. Hypolipidemic potential of Panicum miliare on selected cardiovascular subjects. Anc. Sci. Life 2007, 26, 29–32. [Google Scholar]
- Antia, B.S.; Okokon, J.E.; Umoh, E.E.; Udobang, J.A. Antidiabetic activity of ethanolic leaf extract of Panicum maximum. Int. J. Drug Dev. Res. 2010, 2, 488–492. [Google Scholar]
- Kanife, U.; Odesanmi, O.; Doherty, V. Phytochemical composition and antifungal properties of leaf, stem and florets of Panicum maximum Jacq. (Poaceae). Int. J. Biol. 2012, 4, 64. [Google Scholar] [CrossRef]
- Ub Wijerathne, C.; Park, H.S.; Jeong, H.Y.; Song, J.W.; Moon, O.S.; Seo, Y.W.; Won, Y.S.; Son, H.Y.; Lim, J.H.; Yeon, S.H.; et al. Quisqualis indica Improves Benign Prostatic Hyperplasia by Regulating Prostate Cell Proliferation and Apoptosis. Biol. Pharm. Bull. 2017, 40, 2125–2133. [Google Scholar] [CrossRef]
- Rho, J.; Seo, C.S.; Park, H.S.; Wijerathne, C.U.; Jeong, H.Y.; Moon, O.S.; Seo, Y.W.; Son, H.Y.; Won, Y.S.; Kwun, H.J. Ulmus macrocarpa Hance improves benign prostatic hyperplasia by regulating prostatic cell apoptosis. J. Ethnopharmacol. 2019, 233, 115–122. [Google Scholar] [CrossRef]
- Park, H.S.; Wijerathne, C.U.B.; Jeong, H.Y.; Seo, C.S.; Ha, H.; Kwun, H.J. Gastroprotective effects of Hwanglyeonhaedok-tang against Helicobacter pylori-induced gastric cell injury. J. Ethnopharmacol. 2018, 216, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Andriole, G.; Bruchovsky, N.; Chung, L.W.; Matsumoto, A.M.; Rittmaster, R.; Roehrborn, C.; Russell, D.; Tindall, D. Dihydrotestosterone and the prostate: The scientific rationale for 5alpha-reductase inhibitors in the treatment of benign prostatic hyperplasia. J. Urol. 2004, 172, 1399–1403. [Google Scholar] [CrossRef] [PubMed]
- Pawlicki, B.; Zielinski, H.; Dabrowski, M. Role of apoptosis and chronic prostatitis in the pathogenesis of benign prostatic hyperplasia. Pol. Merkur. Lek. 2004, 17, 307–310. [Google Scholar]
- Reynolds, A.R.; Kyprianou, N. Growth factor signalling in prostatic growth: Significance in tumour development and therapeutic targeting. Br. J. Pharmacol. 2006, 147 (Suppl. S2), S144–S152. [Google Scholar] [CrossRef] [PubMed]
- Gershtein, E.S.; Scherbakov, A.M.; Shatskaya, V.A.; Kushlinsky, N.E.; Krasil’nikov, M.A. Phosphatidylinositol 3-kinase/AKT signalling pathway components in human breast cancer: Clinicopathological correlations. Anticancer Res. 2007, 27, 1777–1782. [Google Scholar] [PubMed]
- Sillman, S.J.; Lee, S.T.; Claborn, J.; Boruch, J.; Harris, S.P. Fall panicum (Panicum dichotomiflorum) toxicosis in three juvenile goats. J. Vet. Diagn. Investig. 2019, 31, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Divers, T.J.; Freckleton, M.L.; McKenzie, H.C.; Mitchell, E.; Cullen, J.M.; McDonough, S.P. Fall panicum (Panicum dichotomiflorum) hepatotoxicosis in horses and sheep. J. Vet. Intern. Med. 2006, 20, 1414–1421. [Google Scholar] [CrossRef] [PubMed]
- Vickman, R.E.; Franco, O.E.; Moline, D.C.; Vander Griend, D.J.; Thumbikat, P.; Hayward, S.W. The role of the androgen receptor in prostate development and benign prostatic hyperplasia: A review. Asian J. Urol. 2020, 7, 191–202. [Google Scholar] [CrossRef]
- Wurzel, R.; Ray, P.; Major-Walker, K.; Shannon, J.; Rittmaster, R. The effect of dutasteride on intraprostatic dihydrotestosterone concentrations in men with benign prostatic hyperplasia. Prostate Cancer Prostatic. Dis. 2007, 10, 149–154. [Google Scholar] [CrossRef]
- Lee, G.; Shin, J.; Choi, H.; Jo, A.; Pan, S.; Bae, D.; Lee, Y.; Choi, C. Cynanchum wilfordii Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia by Regulating 5alpha-Reductase and Androgen Receptor Activities in a Rat Model. Nutrients 2017, 9, 1070. [Google Scholar] [CrossRef]
- Park, H.S.; Seo, C.S.; Wijerathne, C.U.; Jeong, H.Y.; Moon, O.S.; Seo, Y.W.; Won, Y.S.; Son, H.Y.; Lim, J.H.; Kwun, H.J. Effect of Veratrum maackii on Testosterone Propionate-Induced Benign Prostatic Hyperplasia in Rats. Biol. Pharm. Bull. 2019, 42, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Vela-Navarrete, R.; Escribano-Burgos, M.; Farre, A.L.; Garcia-Cardoso, J.; Manzarbeitia, F.; Carrasco, C. Serenoa repens treatment modifies bax/bcl-2 index expression and caspase-3 activity in prostatic tissue from patients with benign prostatic hyperplasia. J. Urol. 2005, 173, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Tzifi, F.; Economopoulou, C.; Gourgiotis, D.; Ardavanis, A.; Papageorgiou, S.; Scorilas, A. The Role of BCL2 Family of Apoptosis Regulator Proteins in Acute and Chronic Leukemias. Adv. Hematol. 2012, 2012, 524308. [Google Scholar] [CrossRef]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol. 2013, 14, 32. [Google Scholar] [CrossRef]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genes Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef]
- Djavan, B.; Margreiter, M.; Dianat, S.S. An algorithm for medical management in male lower urinary tract symptoms. Curr. Opin. Urol. 2011, 21, 5–12. [Google Scholar] [CrossRef]
- Liu, R.F.; Fu, G.; Li, J.; Yang, Y.F.; Wang, X.G.; Bai, P.D.; Chen, Y.D. Roles of autophagy in androgen-induced benign prostatic hyperplasia in castrated rats. Exp. Ther. Med. 2018, 15, 2703–2710. [Google Scholar] [CrossRef]
- Culig, Z.; Hobisch, A.; Cronauer, M.V.; Radmayr, C.; Hittmair, A.; Zhang, J.; Thurnher, M.; Bartsch, G.; Klocker, H. Regulation of prostatic growth and function by peptide growth factors. Prostate 1996, 28, 392–405. [Google Scholar] [CrossRef]
- Lucia, M.S.; Lambert, J.R. Growth factors in benign prostatic hyperplasia: Basic science implications. Curr. Urol. Rep. 2008, 9, 272–278. [Google Scholar] [CrossRef]
- Sampson, N.; Zenzmaier, C.; Heitz, M.; Hermann, M.; Plas, E.; Schafer, G.; Klocker, H.; Berger, P. Stromal insulin-like growth factor binding protein 3 (IGFBP3) is elevated in the diseased human prostate and promotes ex vivo fibroblast-to-myofibroblast differentiation. Endocrinology 2013, 154, 2586–2599. [Google Scholar] [CrossRef]
- Descazeaud, A.; Weinbreck, N.; Robert, G.; Vacherot, F.; Abbou, C.C.; Labrousse, F.; Allory, Y.; Rubin, M.A.; de la Taille, A. Transforming growth factor beta-receptor II protein expression in benign prostatic hyperplasia is associated with prostate volume and inflammation. BJU Int. 2011, 108, E23–E28. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.C.; Wang, Y.; Zhang, S.W.; Luo, D.K.; Chang, D.G.; Wu, X.Q.; Tang, M.; He, Z.M. Angiogenesis and regulatory factors in rats with BPH induced by testosterone. Zhonghua Nan Ke Xue 2005, 11, 413–418. [Google Scholar] [PubMed]
- Shah, A.; Shah, A.A.; Nandakumar, K.; Lobo, R. Mechanistic targets for BPH and prostate cancer-a review. Rev. Environ. Health 2020, 36, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Wozney, J.L.; Antonarakis, E.S. Growth factor and signaling pathways and their relevance to prostate cancer therapeutics. Cancer Metast. Rev. 2014, 33, 581–594. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Macoska, J.A. Chemokines and BPH/LUTS. Differentiation 2011, 82, 253–260. [Google Scholar] [CrossRef]
- McLaren, I.D.; Jerde, T.J.; Bushman, W. Role of interleukins, IGF and stem cells in BPH. Differentiation 2011, 82, 237–243. [Google Scholar] [CrossRef]
- Penna, G.; Fibbi, B.; Amuchastegui, S.; Cossetti, C.; Aquilano, F.; Laverny, G.; Gacci, M.; Crescioli, C.; Maggi, M.; Adorini, L. Human Benign Prostatic Hyperplasia Stromal Cells As Inducers and Targets of Chronic Immuno-Mediated Inflammation. J. Immunol. 2009, 182, 4056–4064. [Google Scholar] [CrossRef]
- Schenk, J.M.; Kristal, A.R.; Neuhouser, M.L.; Tangen, C.M.; White, E.; Lin, D.W.; Kratz, M.; Thompson, I.M. Biomarkers of systemic inflammation and risk of incident, symptomatic benign prostatic hyperplasia: Results from the prostate cancer prevention trial. Am. J. Epidemiol. 2010, 171, 571–582. [Google Scholar] [CrossRef]
- Mechergui, Y.B.; Ben Jemaa, A.; Mezigh, C.; Fraile, B.; Ben Rais, N.; Paniagua, R.; Royuela, M.; Oueslati, R. The Profile of Prostate Epithelial Cytokines and its Impact on Sera Prostate Specific Antigen Levels. Inflammation 2009, 32, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.R.; Li, Q.J.; Han, P.; Li, X.; Zeng, H.; Zhu, Y.C.; Wei, Q. Evaluation of Interleukin-8 in Expressed Prostatic Secretion as a Reliable Biomarker of Inflammation in Benign Prostatic Hyperplasia. Urology 2009, 74, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Z.; Yang, L.; Zhao, H.; Yue, C.; Li, J.; Han, L. Expression of IL-6 and TNF-α in benign prostatic hyperplasia combined with histological inflammation. Discuss. Clin. Cases 2016, 3, 11–15. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).