Antioxidant and Hepatoprotective Effect of Cajanus cajan in N-Nitrosodiethylamine-Induced Liver Damage


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Collection and Preparation of Plant Extract
2.4. Experimental Design
2.5. Measurement of Body Weight
2.6. Collection of Blood and Liver Tissue
2.7. Measurement of Liver Weight
2.8. Preparation of Liver Homogenate
2.9. Measurement of ALT and AST Activity
2.10. Measurement of Albumin (ALB)
2.11. Measurement of Reduced Glutathione (GSH)
2.12. Measurement of Glutathione S-Transferase (GST) Activity
2.13. Measurement of Superoxide Dismutase (SOD) Activity
2.14. Measurement of Catalase (CAT) Activity
2.15. Measurement of Malondialdehyde (MDA)
2.16. Histological Studies
2.17. Statistical Analysis
3. Results
3.1. Effect of Cajanus cajan on Body and Liver Weight
3.2. Effect of Cajanus cajan on Plasma AST and ALT
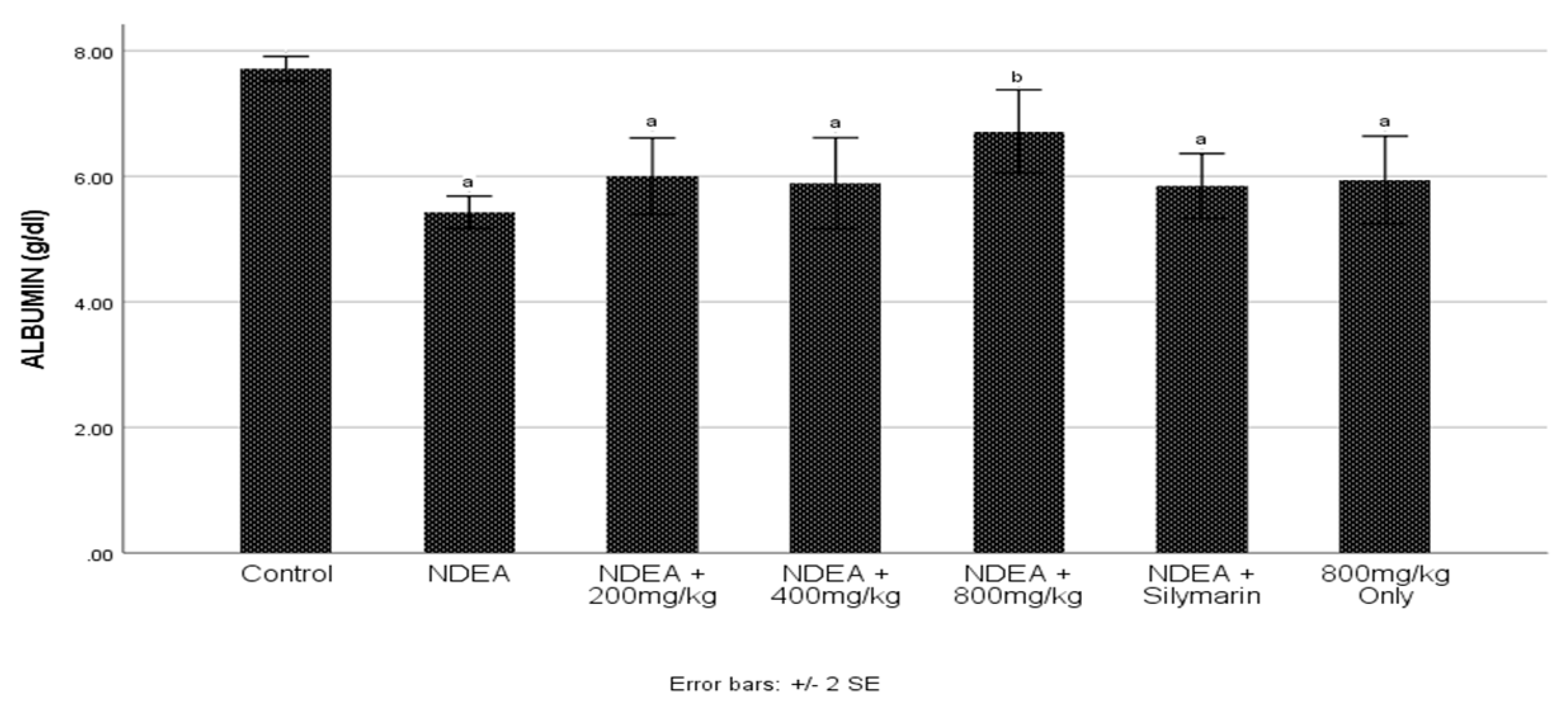
3.3. Effect of Cajanus cajan on Plasma Albumin
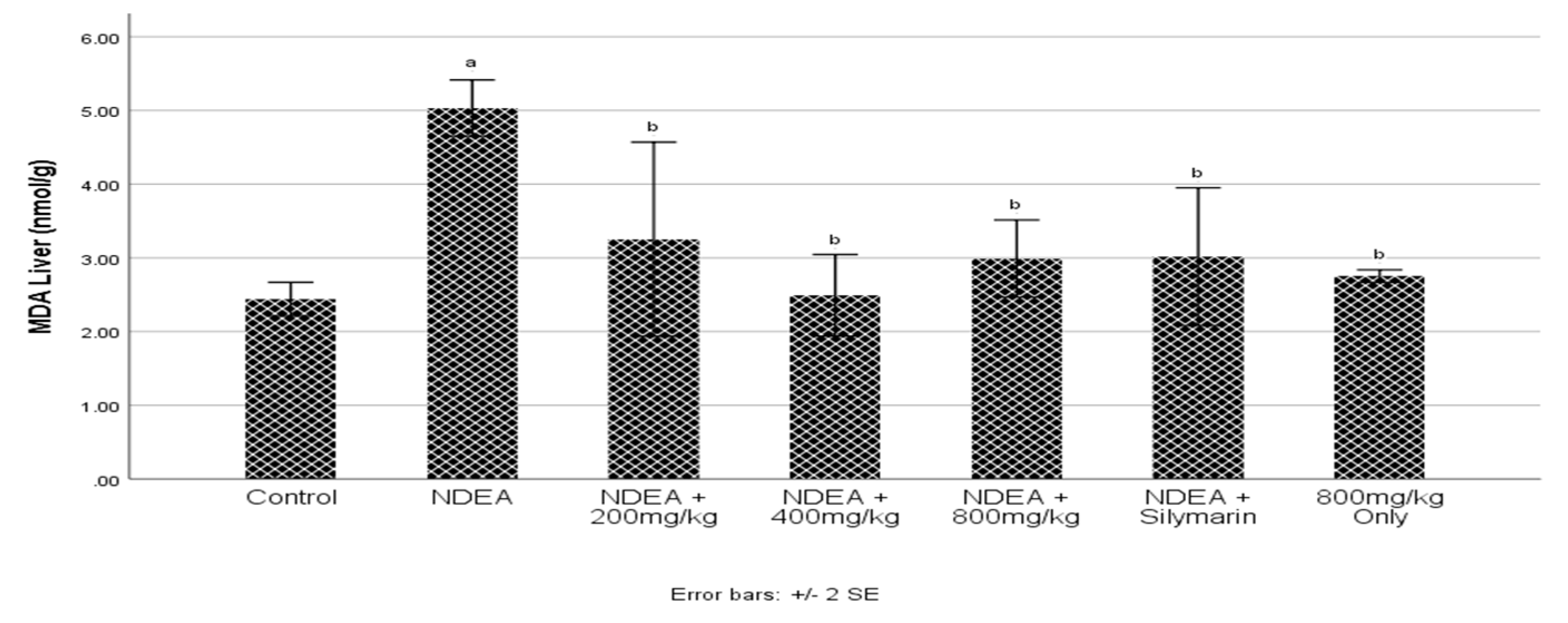
3.4. Effect of Cajanus cajan on Malondialdehyde (MDA)
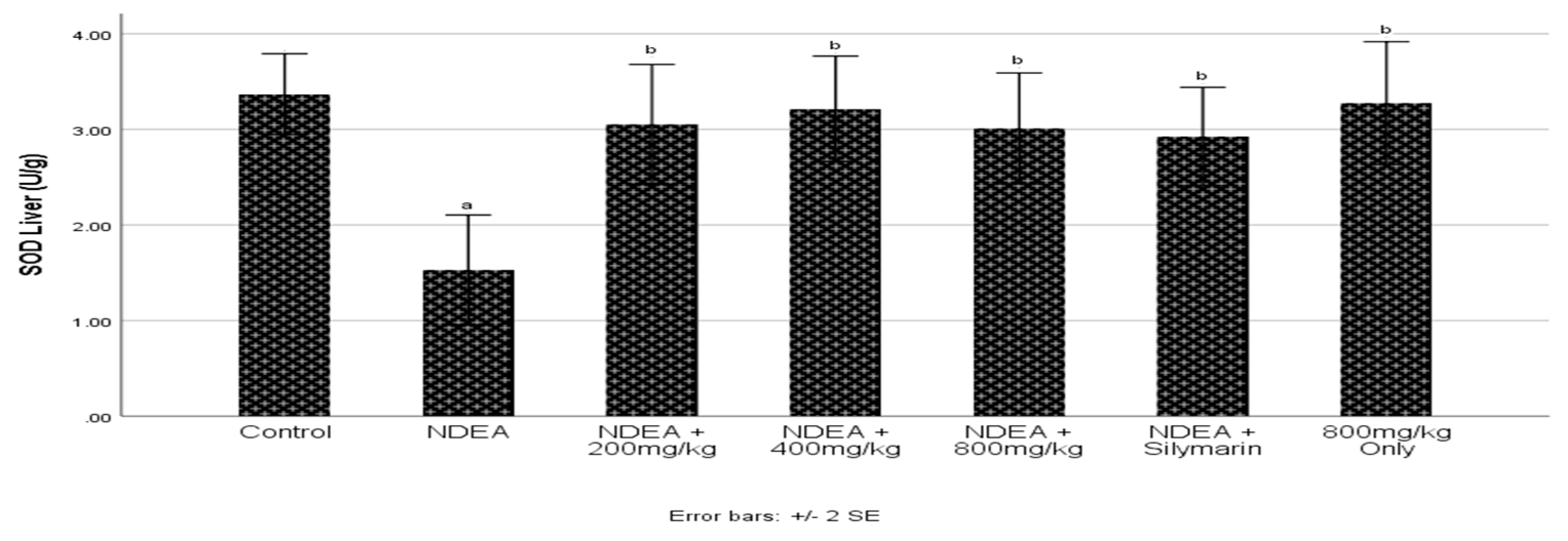
3.5. Effect of Cajanus cajan on Superoxide Dismutase (SOD) Activity
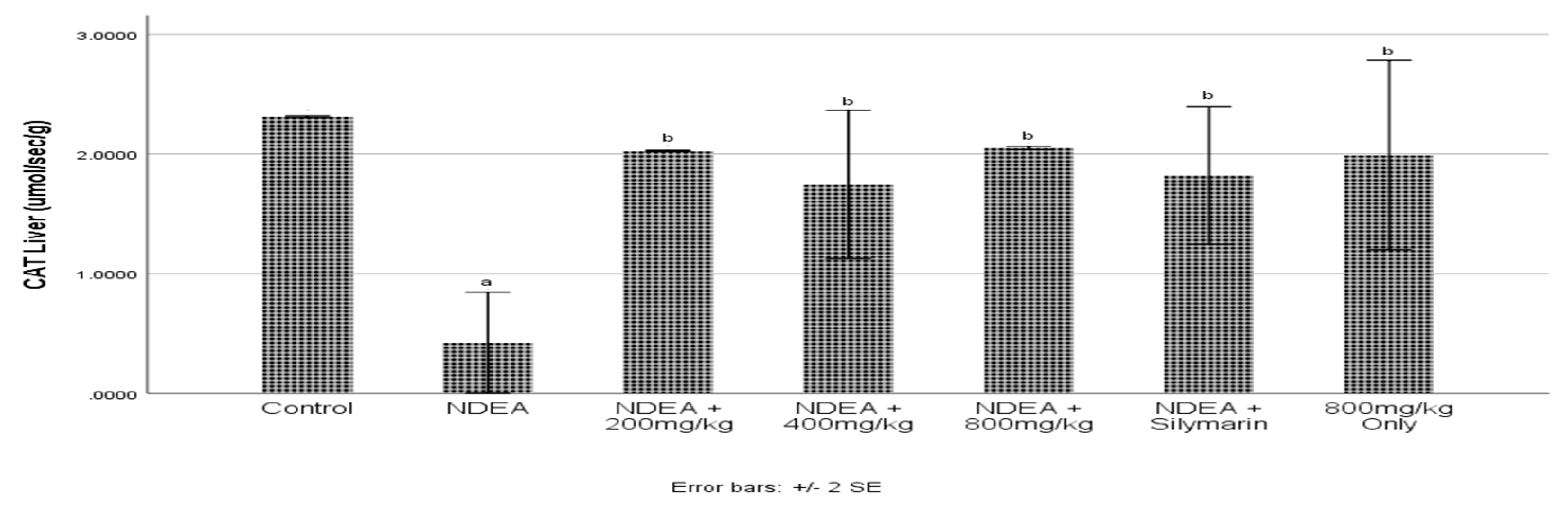
3.6. Effect of Cajanus cajan on Catalase Activity
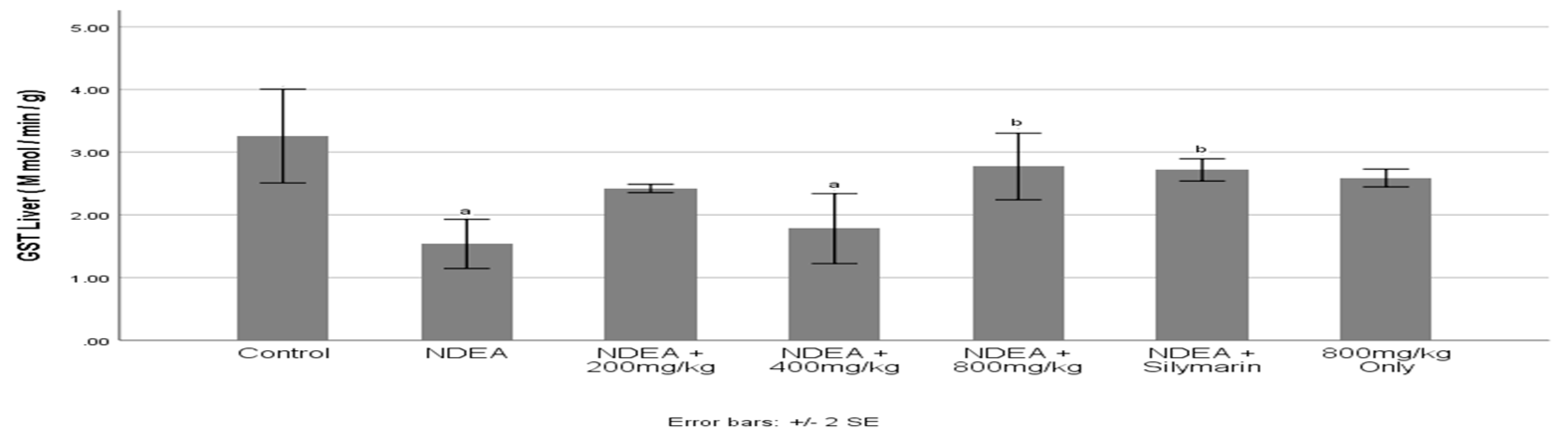
3.7. Effect of Cajanus cajan on Glutathione S-Transferase (GST) activity
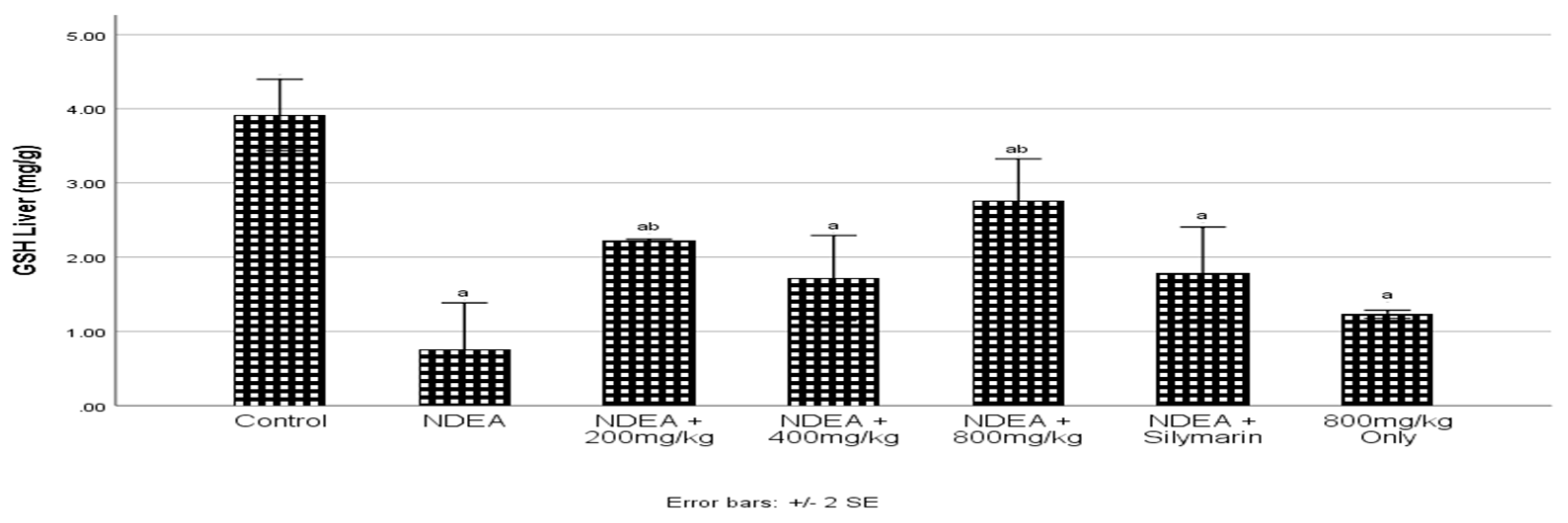
3.8. Effect of Cajanus cajan on Reduced Glutathione (GSH)
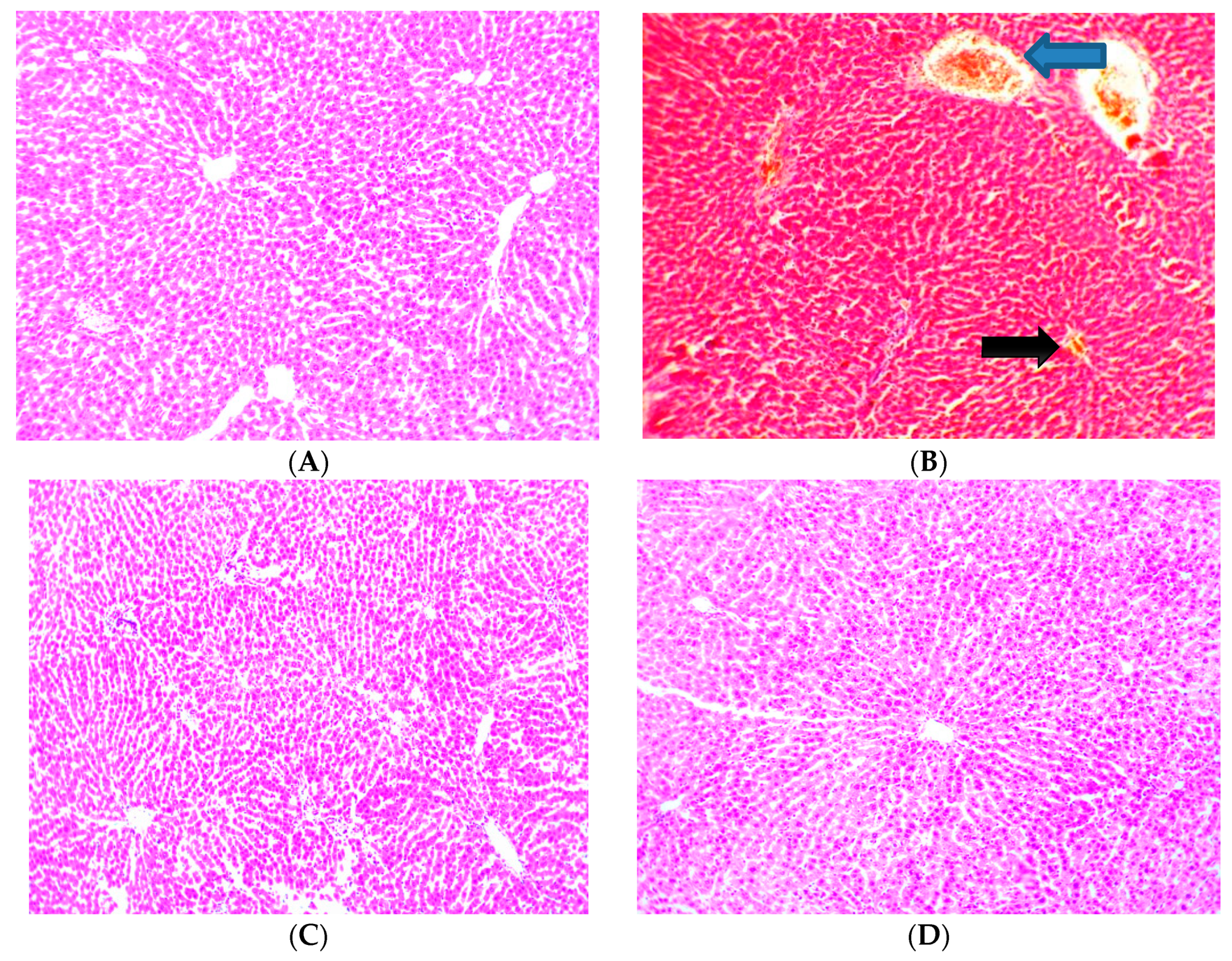
3.9. Effect of Cajanus cajan on Histological Features of the Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tricker, A.; Pfundstein, B.; Theobald, E.; Preussman, R.; Spiegenhalder, B. Mean daily intake of volatile N-nitrosamines from foods and beverages in West Germany. Food Chem. Toxicol. 1989, 29, 29–32. [Google Scholar]
- Shank, C. Toxicology of N-nitroso compounds. Toxicol. Appl. Pharm. 1975, 31, 729–732. [Google Scholar] [CrossRef]
- Lijinsky, W. N-Nitroso compounds in the diet. Mutat. Res. Genet. Toxicol. Environ. Mutagenes. 1999, 443, 129–138. [Google Scholar] [CrossRef]
- Bartsch, H.; Hietanen, E.; Malaveille, C. Carcinogenic nitrosamines: Free radical aspects of their action. Free Rad. Biol. Med. 1989, 7, 637–644. [Google Scholar] [CrossRef]
- Farber, J.; Gerson, R. Mechanisms of cell injury with hepatotoxic chemicals. Pharmacol. Rev. 1984, 36, 71–85. [Google Scholar]
- George, J.; Rao, K.R.; Stern, R.; Chandrakasan, G. Dimethylnitrosamine-induced liver injury in rats: The early deposition of collagen. Toxicology 2001, 156, 129–138. [Google Scholar] [CrossRef]
- Gey, K.F. Prospects for the prevention of free radical disease, regarding cancer and cardiovascular disease. Br. Med. Bull. 1993, 49, 679–699. [Google Scholar] [CrossRef]
- Arul, D.; Subramanian, P. Inhibitory effect of naringenin (citrus flavonone) on N-nitrosodiethylamine induced hepatocarcinogenesis in rats. Biochem. Biophys. Res. Commmun. 2013, 434, 203–209. [Google Scholar] [CrossRef]
- Singh, M.; Kaur, P.; Sandhir, R.; Kiran, R. Protective effects of vitamin E against atrazine-induced genotoxicity in rats. Mutat. Res. 2008, 654, 145–149. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell. Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Zu, Y.; Fu, Y.; Liu, W.; Hou, L.; Kong, Y. Simultaneous determination of four flavonoids in pigeon pea (Cajanus cajan) leaves using RP-LC. Chromatographia 2006, 63, 499–505. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, S.; Zu, Y.; Fu, Y.; Kong, Y.; Gao, Y. Negative pressure cavitation extraction and antioxidant activity of genistein and genistin from the roots of pigeon pea [Cajanus cajan (L.) Millsp.]. Sep. Purif. Technol. 2010, 74, 261–270. [Google Scholar] [CrossRef]
- Lu, L.M.; Zu, Y.; Fu, Y.; Zhang, S.; Yao, L.; Efferth, T. Cajanol, a novel anticancer agent from Pigeon pea roots, induces apoptosis in human breast cancer cells through a ROS-mediated mitochondrial pathway. Chem. Biol. Interact. 2010, 188, 151–160. [Google Scholar] [CrossRef]
- Ahsan, R.; Islam, M. In vitro antibacterial screening and toxicological study of some useful plants (Cajanus cajan). Eur. J. Sci. Res. 2009, 41, 227–232. [Google Scholar]
- Ito, N.; Imaida, K.; Hasegawa, R.; Tsuda, H. Rapid bioassay methods for carcinogens and modifiers of hepatocarcinogenesis. Crit. Rev. Toxicol. 1989, 19, 385–415. [Google Scholar]
- Iweala, E.; Ogidigo, J. Prostate Specific Antigen, Antioxidant and Hematological Parameters in Prostatic Rats Fed Solanum macrocarpon Leaves. Asian J. Biol. Sci. 2015, 8, 30–41. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, B.R.; Sarma, B.K.; Singh, B. Potential chemoprevention of N-nitrosodiethylamine-induced hepatocarcinogenesis by polyphenolics from Acacia nilotica bark. Chem. Biol. Int. 2009, 181, 20–28. [Google Scholar] [CrossRef]
- Prins, H.; Loos, J. Biochemical methods in red cell genetics. Eur. J. Pharm. 1969, 129, 122–125. [Google Scholar]
- Habig, W.H.; Pabst, J.; Fleischner, G.; Gatmaitan, Z.; Arias, M.; Jakoby, B. The identity of glutathione S-transferase with ligandin, a major binding protein of liver. Proc. Natl. Acad. Sci. USA 1974, 71, 3879–3882. [Google Scholar] [CrossRef]
- Stevens, M.; Obrosova, I.; Cao, X.; Van, C.; Greene, D. Effects of DLα-lipoic acid on peripheral nerve conduction, blood flow, energy metabolism, and oxidative stress in experimental diabetic neuropathy. Diabetes 2000, 49, 1006–1015. [Google Scholar] [CrossRef]
- Bock, P.; Kramer, R.; Pavelka, M. Peroxisomes and related particles. Cell Biol. Monog. 1980, 7, 44–74. [Google Scholar]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Bancroft, J.; Gamble, M. Theory and Practice of Histological Techniques. Eur. J. Sci. Res. 2002, 38, 302–310. [Google Scholar]
- Zimmerman, J. The adverse effects of drugs and other chemicals on the liver. Sep. Purif. Technol. 1978, 91, 121–133. [Google Scholar]
- Ahmad, A.; Maheshwari, V.; Ahmad, A.; Saleem, R.; Ahmad, R. Observation of esterase-like-albumin activity during N’-Nitrosodimethylamine induced hepatic fibrosis in a mammalian model. Maced. J. Med. Sci. 2012, 5, 55–61. [Google Scholar] [CrossRef]
- Miyao, H.; Ara, O.; Udayma, M.; Kinji, J.; Mahara, T. Kaikosaponin III and soyasaponin I, major triterpene saponins of Abrus cantoniensis, act on GOT and GPT influence on transamination elevation of rat liver cells concomitantly exposed to eclipse for one hour. Planta Med. 1998, 9, 65. [Google Scholar]
- Pawar, R.; Gopalakrishnan, C.; Bhutani, K. Dammarane triterpene saponin from Bacopa monniera as the superoxide inhibitor in polymorphonuclear cells. Planta Med. 2001, 67, 752–754. [Google Scholar] [CrossRef]
- Oluseyi, A.; Moshood, O. Hepatoprotective effect of Cajanus cajan on tissue defense system in D-galactosamine-induced hepatitis in rats. Turk. J. Biochem. 2011, 36, 237–241. [Google Scholar]
- Wahid, A.; Vijay, R.; Satish, B. Hepatoprotective activity of hydroalcoholic extract leaves of Alocassie indica. Ind. J. Exp. Biol. 2009, 47, 816–821. [Google Scholar]
- Zeashan, H.A.; Amresh, G.A.; Satyawan, S.B.; Venkateswara, Q. Hepatoprotective activity of Amaranthusspinosus in experimental animals. Food Chem. Toxicol. 2008, 46, 3417–3421. [Google Scholar] [CrossRef]
- Amresh, G.; Kant, R.; Zeashan, H.; Gupta, R.; Rao, V.; Singh, P. Gastro protective effects of ethanolic extract from Cissampelos pareira in experimental animals. J. Nat. Med. 2007, 61, 323–328. [Google Scholar] [CrossRef]
- Sankaran, M.; Vadivel, A.; Thangam, A. Curative effect of garlic on alcoholic liver disease patients. J. Biol. Sci. 2010, 3, 147–152. [Google Scholar]
- Meister, A.; Anderson, E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Bansal, A.K.; Trivedi, R.; Soni, G.; Bhatnagar, D. Hepatic and renal oxidative stress in acute toxicity of N-nitrosodiethylamine in rats. Ind. J. Exp. Biol. 2000, 38, 916–920. [Google Scholar]
- Kweon, S.; Park, K.A.; Choi, H. Chemopreventive effect of garlic powder diet in diethylnitrosamine-induced rat hepatocarcinogenesis. Life Sci. 2003, 73, 2515–2526. [Google Scholar] [CrossRef]
- Yu, P. Cellular defense against damage from reactive oxygen species. Physiol. Rev. 1994, 74, 136–162. [Google Scholar] [CrossRef]
- Fraschini, F.; Demartini, G.; Esposti, D. Pharmacology of silymarin. Clin. Drug. Investig. 2002, 22, 51–65. [Google Scholar] [CrossRef]
- Adaramoye, Y.; Adeyemi, U. Effect of kolaviron, a biflavonoid complex from Garcinia kola seeds, on ethanol-induced oxidative stress in liver of adults Wistar rats. J. Med. Food 2009, 12, 584–590. [Google Scholar] [CrossRef]
- Jeyabal, P.V.; Syed, M.B.; Venkataraman, M.; Sambandham, J.; Sakthisekaran, A. Apigenin inhibits oxidative stress-induced macromolecular damage in N-nitrosodiethylamine (NDEA)-induced hepatocellular carcinogenesis in Wistar albino rats. Mol. Carcinog. 2005, 44, 11–20. [Google Scholar] [CrossRef]
- Horie, T.; Awazu, S.; Itakura, Y. Identified diallyl polysul-fides from an aged garlic extract which protects the membranes from lipid peroxidation. Planta Med. 1992, 58, 468–469. [Google Scholar] [CrossRef]
- Junnila, M.; Rahko, A.; Sukura, A. Reduction of carbon tetrachloride induced hepatotoxic effects by oral administration of betaine in male Han-Wistar rats. Vet. Pathol. 2000, 37, 231–238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Treatment | Initial Body Weight (g) | Final Body Weight (g) | Liver Weight (g) | Relative Liver Weight (Liver/100 g b.w) |
|---|---|---|---|---|---|
| A | Control | 168 ± 12.90 | 198 ± 10.72 | 7.08 ± 0.24 | 3.58 ± 0.11 |
| B | NDEA | 160 ± 14.0 a | 170 ± 11.60 a | 9.93 ± 0.44 a | 5.98 ± 0.36 a |
| C | NDEA + 200 mg/kg | 166 ± 11.49 b | 182 ± 13.24 b | 8.16 ± 0.35 b | 4.48 ± 0.25 b |
| D | NDEA + 400 mg/kg | 164 ± 12.46 b | 185 ± 13.26 b | 7.55 ± 0.26 b | 4.19 ± 0.16 b |
| E | NDEA + 800 mg/kg | 163 ± 11.47 | 189 ± 13.29 b | 7.04 ± 0.22 b | 4.01 ± 0.13 b |
| F | NDEA + Silymarin | 161 ± 11.41 | 186 ± 15.08 b | 7.58 ± 0.11b | 3.91 ± 0.25 b |
| G | 800 mg/kg Only | 167 ± 11.06 | 195 ± 11.38 | 6.97± 0.35 | 3.57 ± 0.13 |
| Groups | Treatment | ALT (U/L) | AST (U/L) |
|---|---|---|---|
| A | Control | 69.73.38 ± 5.19 | 106.67 ± 5.47 |
| B | NDEA | 124.73 ± 22.06 a | 227.15 ± 25.5 a |
| C | NDEA + 200 mg/kg | 109.23 ± 8.39 b | 180.34 ± 8.25 b |
| D | NDEA + 400 mg/kg | 98.34 ± 6.81 b | 145.05 ± 6.75 b |
| E | NDEA + 800 mg/kg | 74.86 ± 5.63 b | 112.87 ± 5.80 b |
| F | NDEA + Silymarin | 90.59 ± 6.77 b | 132.12 ± 5.96 b |
| G | 800 mg/kg Only | 70.33 ± 5.22 | 109 ± 5.76 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iweala, E.E.J.; Evbakhavbokun, W.O.; Maduagwu, E.N. Antioxidant and Hepatoprotective Effect of Cajanus cajan in N-Nitrosodiethylamine-Induced Liver Damage. Sci. Pharm. 2019, 87, 24. https://doi.org/10.3390/scipharm87030024
Iweala EEJ, Evbakhavbokun WO, Maduagwu EN. Antioxidant and Hepatoprotective Effect of Cajanus cajan in N-Nitrosodiethylamine-Induced Liver Damage. Scientia Pharmaceutica. 2019; 87(3):24. https://doi.org/10.3390/scipharm87030024
Chicago/Turabian StyleIweala, Emeka Eze Joshua, Winifred Osa Evbakhavbokun, and Emmanuel Ndubisi Maduagwu. 2019. "Antioxidant and Hepatoprotective Effect of Cajanus cajan in N-Nitrosodiethylamine-Induced Liver Damage" Scientia Pharmaceutica 87, no. 3: 24. https://doi.org/10.3390/scipharm87030024
APA StyleIweala, E. E. J., Evbakhavbokun, W. O., & Maduagwu, E. N. (2019). Antioxidant and Hepatoprotective Effect of Cajanus cajan in N-Nitrosodiethylamine-Induced Liver Damage. Scientia Pharmaceutica, 87(3), 24. https://doi.org/10.3390/scipharm87030024