Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Strains Bacteria
2.3. Preparation of Denatured HEWL with Heat Treatment
2.4. Preparation of Denatured HEWL with a DTT Treatment
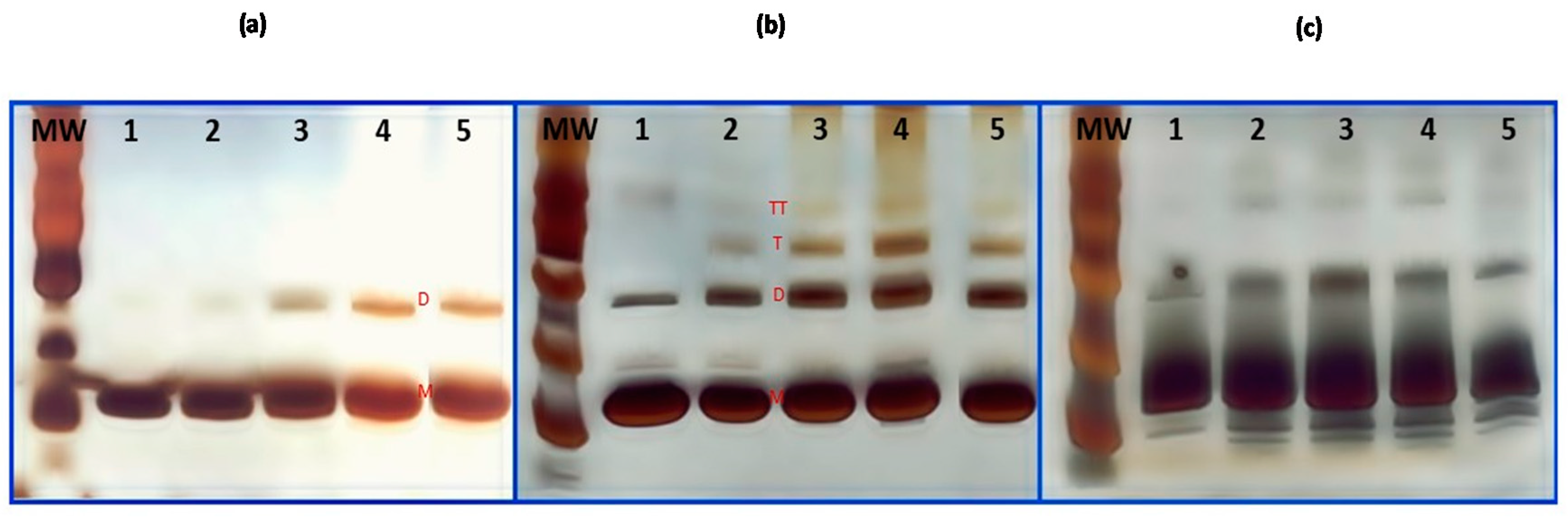
2.5. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis
2.6. HEWL Lytic Activity of the Assay
2.7. Antibacterial Assay
2.8. Fluorescence Microscopy
2.9. Electron Microscopy
2.10. Statistical Analysis
3. Results and Discussion
3.1. Denatured HEWL by Thermal and Chemical Treatments
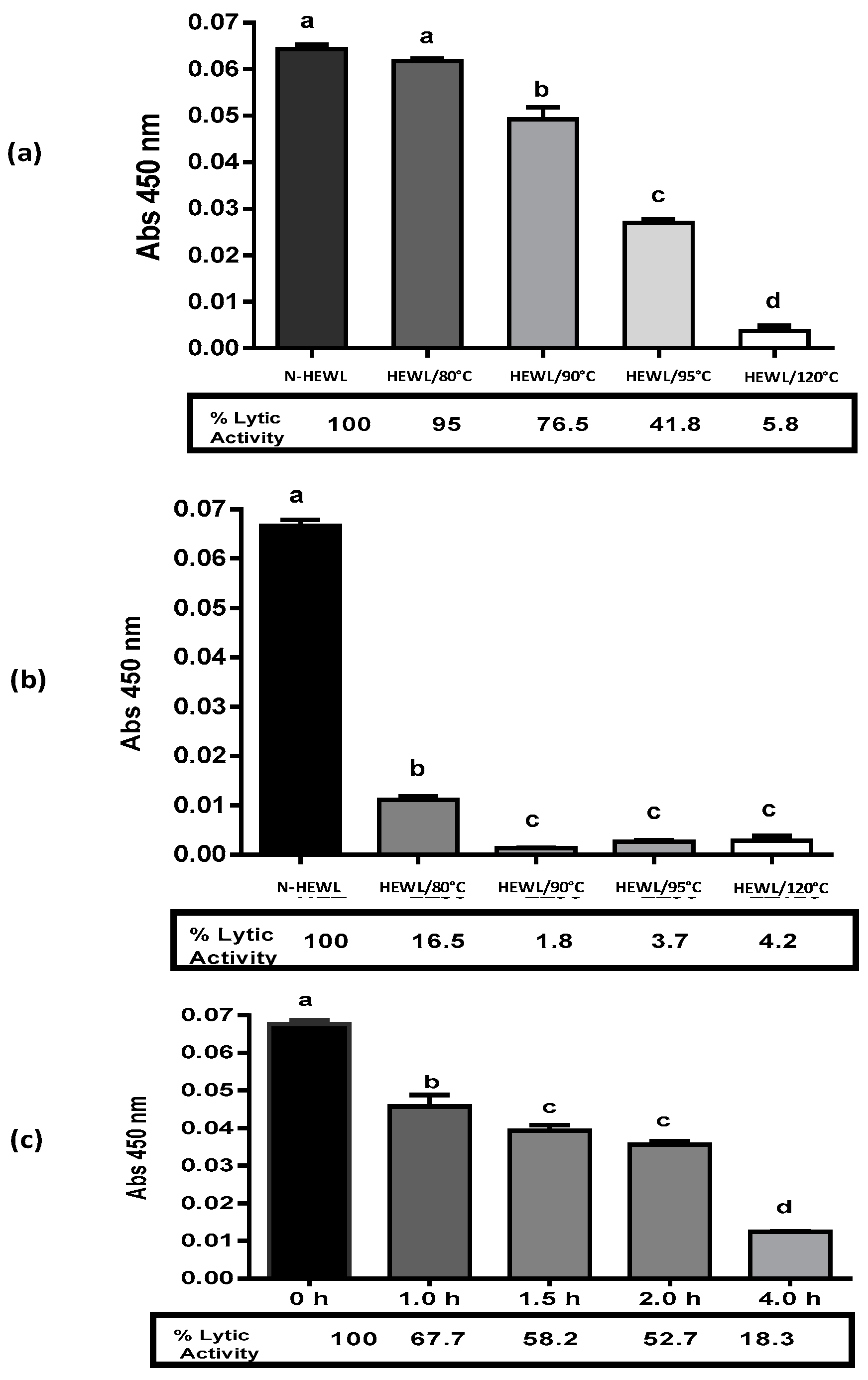
3.2. Denatured HEWL Lytic Enzymatic Activity
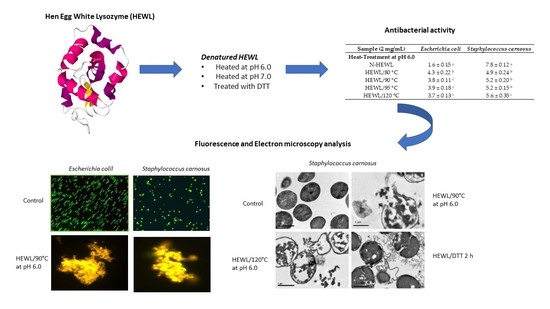
3.3. Antibacterial Activity of Modified HEWL
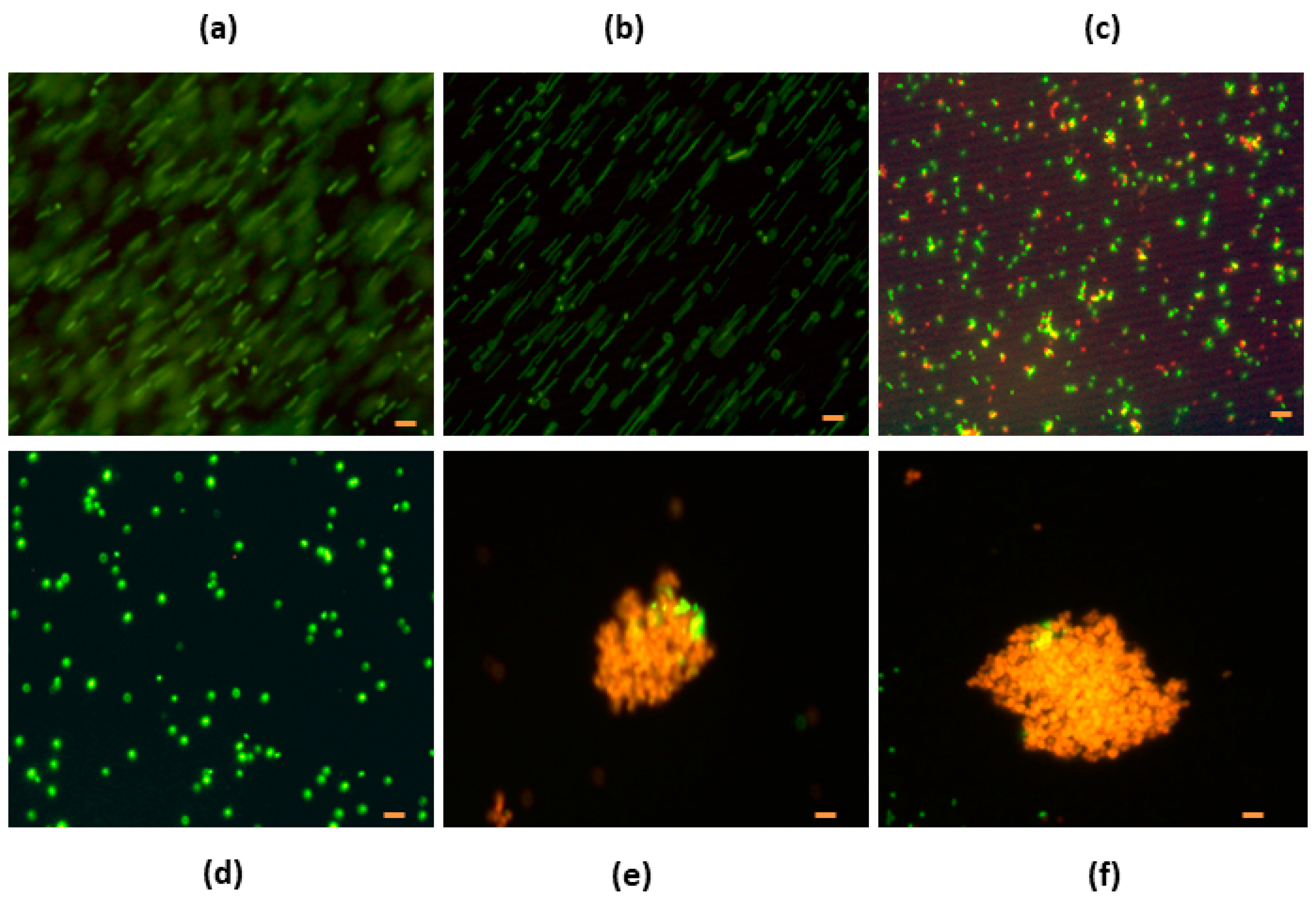
3.4. Fluorescent Analysis
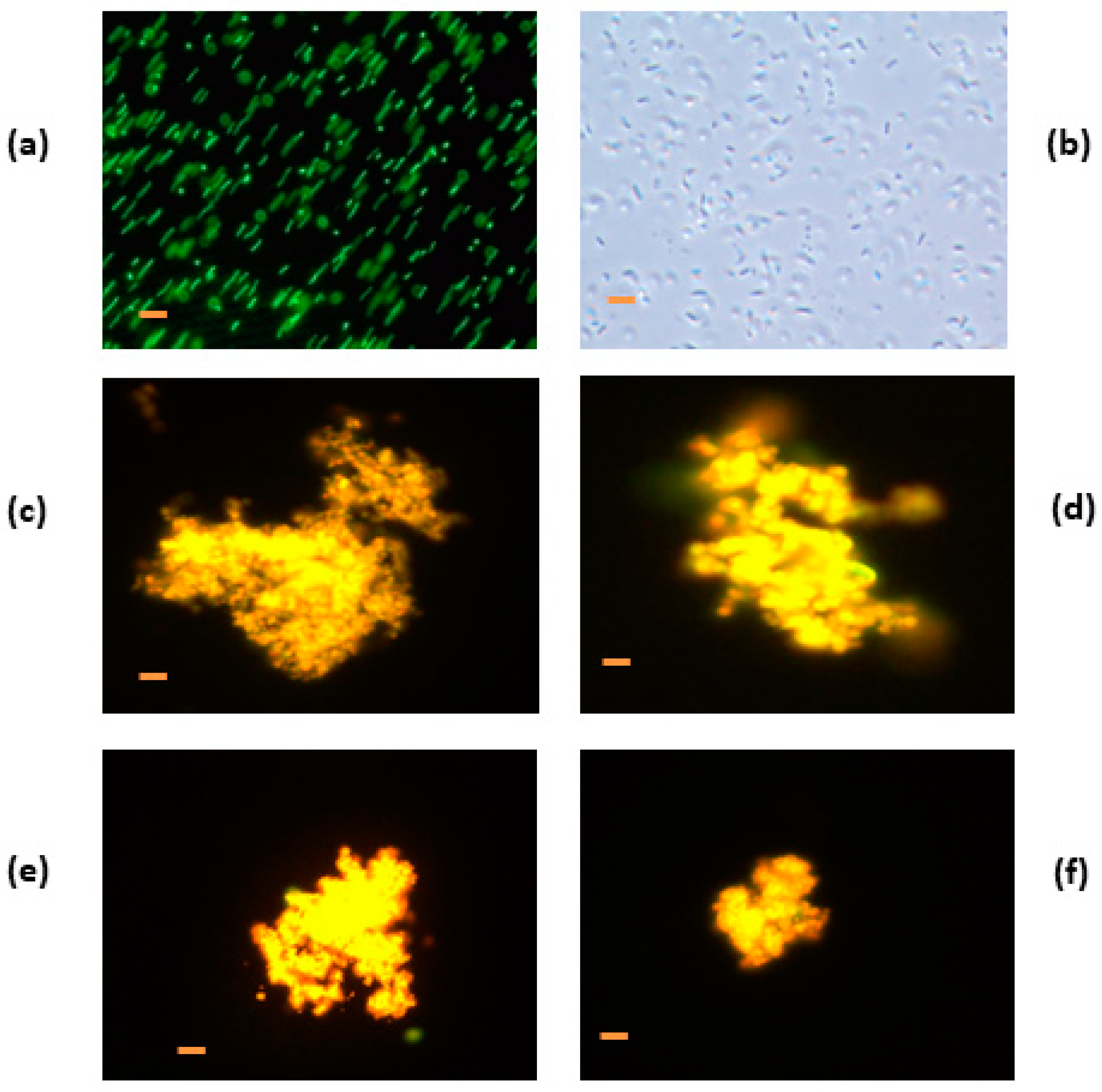
3.4.1. Denatured HEWL at pH 6.0
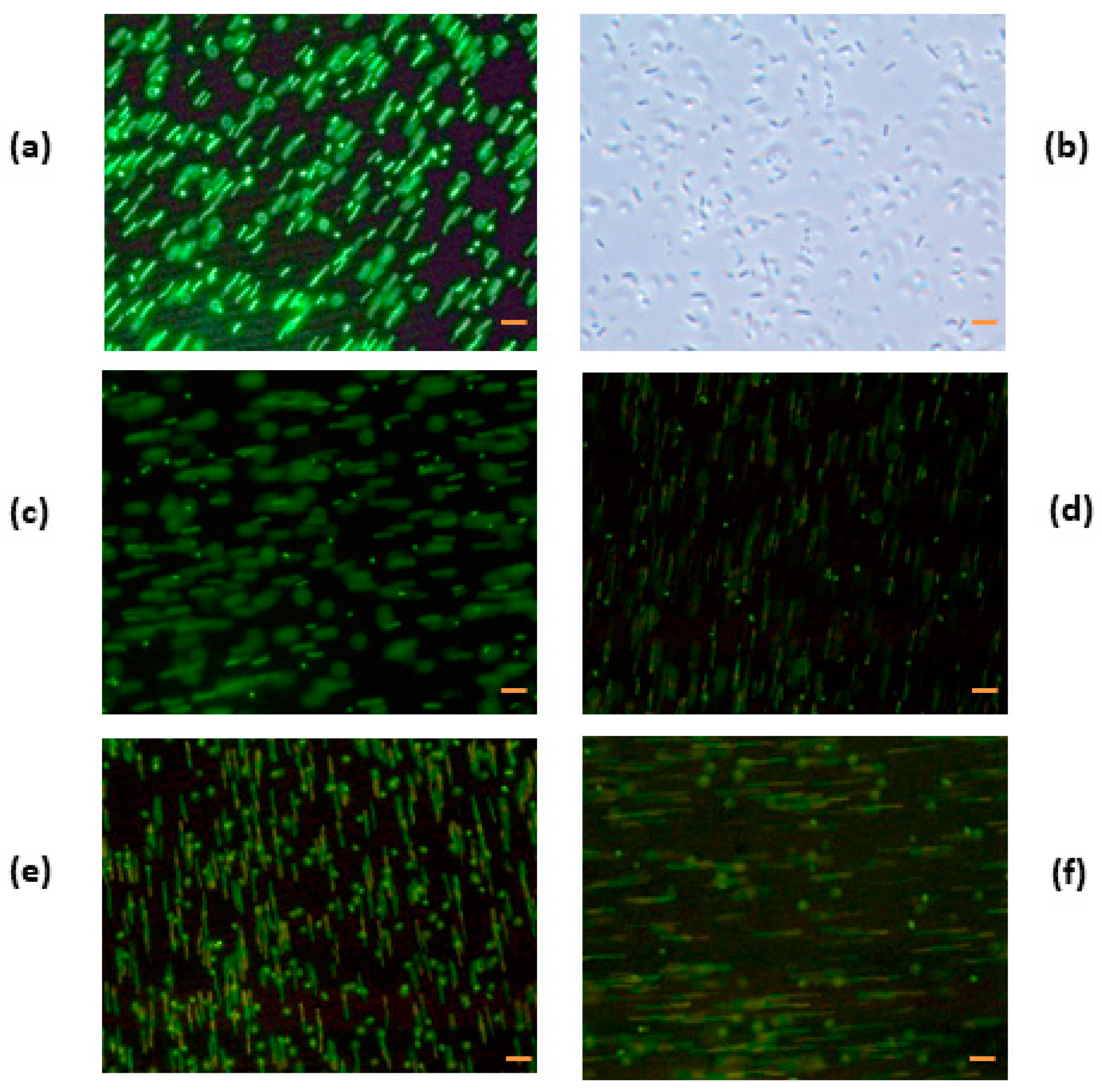
3.4.2. Denatured HEWL at pH 7.0
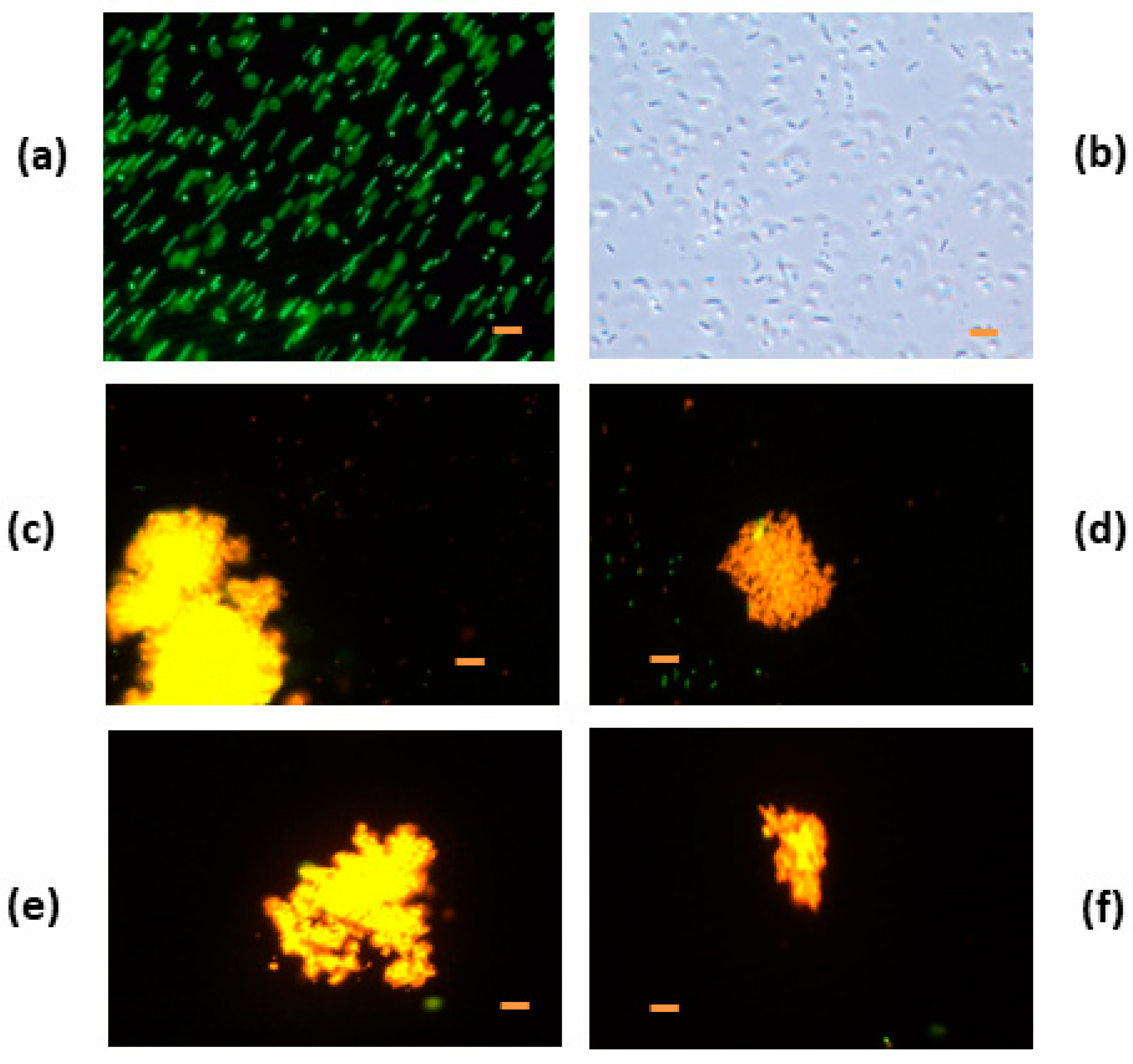
3.4.3. Denatured HEWL/DTT
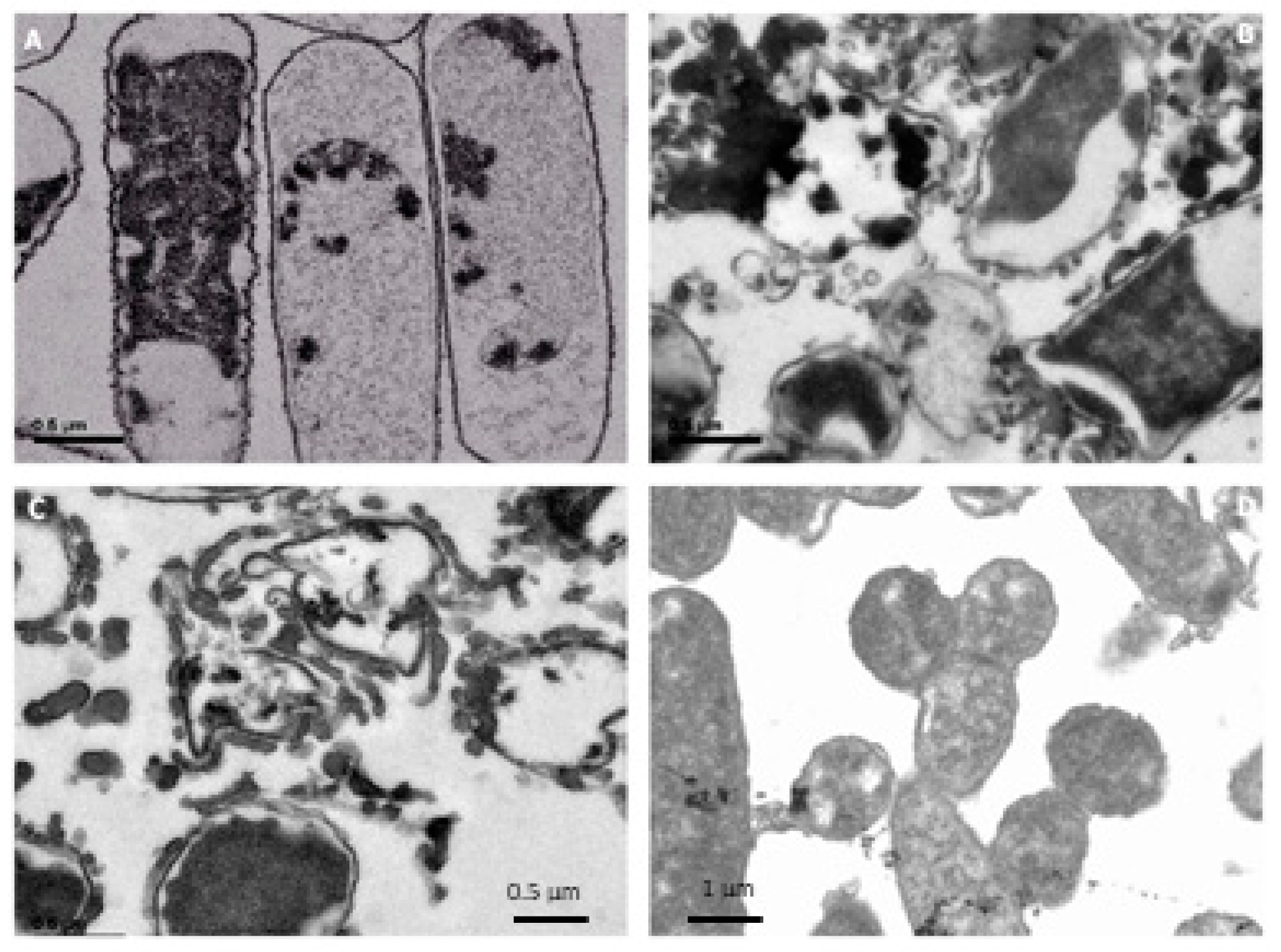
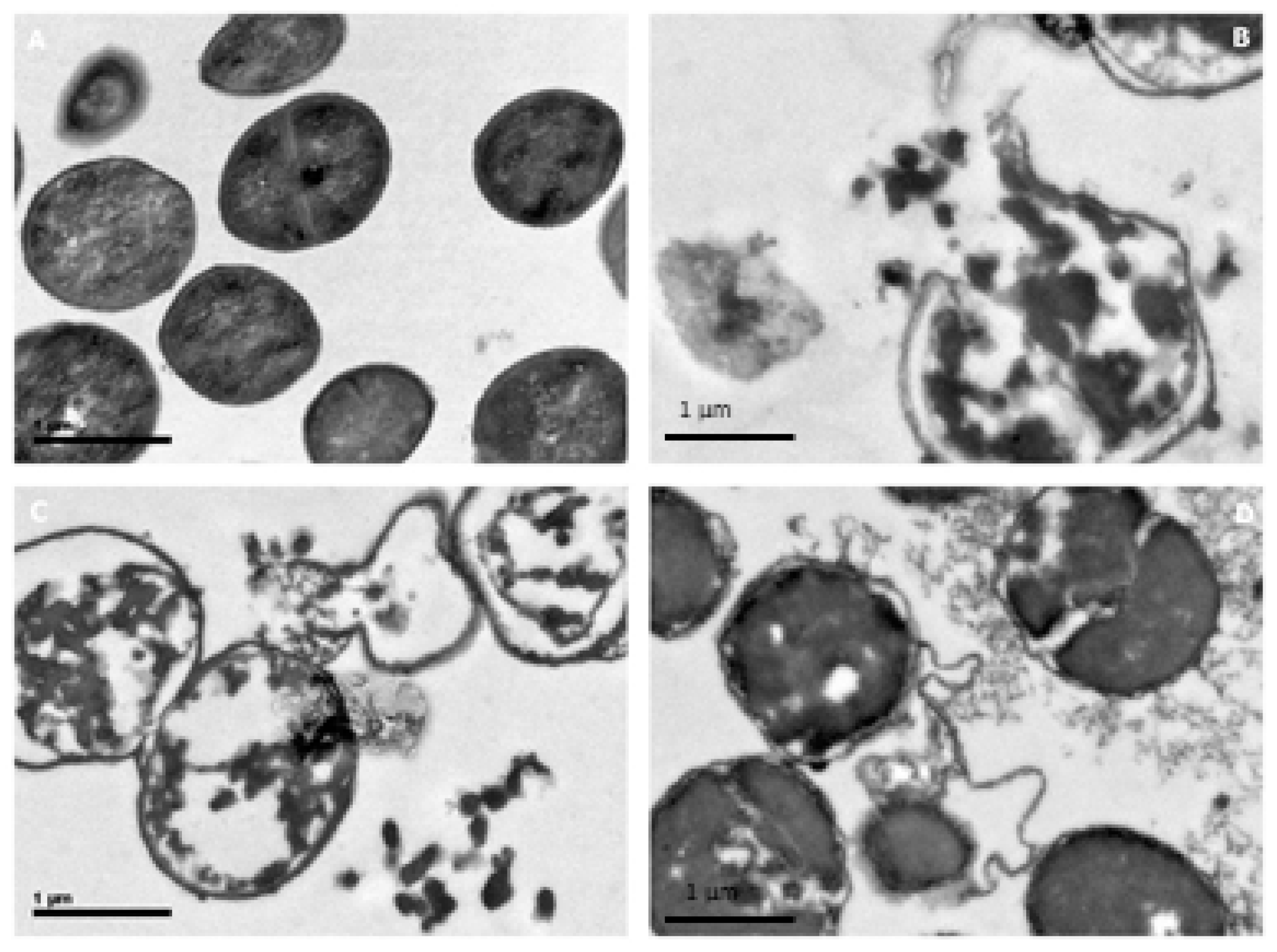
3.5. Electron Microscopy Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lesnierowski, G.; Kijowski, J. Lysozyme: Bioactive Compounds; Huopalahti, R., López-Fandiño, R., Amton, M., Schade, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 33–42. ISBN 978-3-540-37883-9. [Google Scholar]
- Carrillo, W.; Gómez-Ruiz, J.A.; Miralles, B.; Ramos, M.; Barrio, D.; Recio, I. Identification of antioxidant peptides of hen egg-white lysozyme and evaluation of inhibition of lipid peroxidation and cytotoxicity in the Zebrafish model. Eur. Food Res. Technol. 2016, 242, 1777–1785. [Google Scholar] [CrossRef]
- Jiménez-Saiz, R.; Gordon, M.E.; Carrillo, W. Hen Egg White Lysozyme: Antimicrobial Activity & Allergenicity. Lysozymes: Sources, Functions and Role in Disease; Nova Science Publishers: New York, NY, USA, 2013; pp. 215–226. ISBN 978-1-62257-842-9. [Google Scholar]
- Carrillo, W.; Tubón, J.; Vilcacundo, R. Isolation of hen egg white lysozyme by cation exchange chromatography, analysis of its digestibility and evaluation of the inhibition lipid peroxidation in the zebrafish model. Asian J. Pharm. Clin. Res. 2016, 9, 345–349. [Google Scholar]
- Silvetti, T.; Morandi, S.; Hintersteiner, M.; Brasca, M. Use of hen egg white lysozyme in the food industry. In Egg Innovations and Strategies for Improvements; Academic Press: London, UK, 2017; pp. 233–242. ISBN 978-0-12-800879-9. [Google Scholar]
- Carrillo, W.; Barrio, D.; Welbaum, J.; Carpio, C.; Vilcacundo, R.; Morales, D.; Ortiz, J. Antimicrobial and antioxidant peptides obtained from food proteins. In Bioactive Peptides: Types, Roles and Research; Powell, K., Ed.; Nova Science Publisher: New York, NY, USA, 2017; pp. 37–58. ISBN 978-1-53610-994-8. [Google Scholar]
- Jiménez-Saiz, R.; Martos, G.; Carrillo, W.; López-Fandiño, R.; Molina, E. Susceptibility of lysozyme to in-vitro digestion and immunoreactivity of its digests. Food Chem. 2011, 127, 1719–1726. [Google Scholar] [CrossRef]
- Carrillo, W.; Tubón, J. Digestibility and enzymatic activity in vitro of hen egg white lysozyme. Asian J. Pharm. Clin. Res. 2016, 9, 376–378. [Google Scholar]
- Lesnierowski, G.; Cegielska-Radziejewska, R.; Kijowski, J. Thermally and chemical-thermally modified lysozyme and its bacteriostatic activity. World’s Poult. Sci. J. 2004, 60, 303–310. [Google Scholar] [CrossRef]
- Bugatti, V.; Vertuccio, L.; Viscusi, G.; Gorrasi, G. Antimicrobial Membranes of Bio-Based PA 11 and HNTs Filled with Lysozyme Obtained by an Electrospinning Process. Nanomaterials 2018, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Lesnierowski, G.; Stangierski, J. What’s new in chicken egg research and technology for human health promotion? A review. Trends Food Sci. Technol. 2018, 71, 46–51. [Google Scholar] [CrossRef]
- Carrillo, W.; Spindola, H.; Ramos, M.; Recio, I.; Carvalho, J.E. Anti-inflammatory and anti-nociceptive activities of native and modified hen egg white lysozyme. J. Med. Food. 2016, 19, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, W.; García-Ruiz, A.; Recio, I.; Moreno-Arribas, M.V. Antibacterial activity of hen egg white lysozyme modified by heat and enzymatic treatments against oenological lactic acid bacteria and acetic acid bacteria. J. Food Prot. 2014, 77, 1732–1739. [Google Scholar] [CrossRef] [PubMed]
- Masschalck, B.; Van Houdt, R.; Van Haver, E.G.; Michiels, C.W. Inactivation of gram-negative bacteria by lysozyme, denatured lysozyme, and lysozyme-derived peptides under high hydrostatic pressure. Appl. Environ. Microbiol. 2001, 67, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wu, C.; Fu, S.; Wang, L.; Yuan, C.; Chen, S.; Hu, Y. Integration of lysozyme into chitosan nanoparticles for improving antibacterial activity. Carbohydr. Polym. 2017, 155, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Cegielska-Radziejewska, R.; Lesnierowski, G.; Kijowski, J. Antibacterial activity of hen egg white lysozyme modified by thermochemical technique. Eur. Food Res. Technol. 2009, 228, 841–845. [Google Scholar] [CrossRef]
- Mann, K. Proteomics of Egg White. In Proteomics in Food Science; Kuddus, M., Ed.; Academic Press: London, UK, 2017; pp. 261–276. ISBN 9780128132807. [Google Scholar]
- Gorbenko, G.P.; Loffe, V.M.; Kinnunen, P.K.J. Binding of lysozyme to phospholipid bilayers: Evidence for protein aggregation upon membrane association. Biophys. J. 2007, 93, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Yamada, M.; Matsushita, K.; Kobayashi, K.; Kato, A. Enhanced bactericidal action of lysozyme to E. coli by inserting a hydrophobic pentapeptide into its C terminus. J. Biol. Chem. 1994, 269, 5059–5063. [Google Scholar] [PubMed]
- Ibrahim, H.R.; Higashiguchi, S.; Keats, M.; Juneja, F.; Kim, M.; Yamamoto, T.; Sugimoto, Y.; Aoki, T. Partially unfolded lysozyme at neutral pH agglutinates and kills Gram-negative and Gram-positive bacteria through membrane damage mechanism. J. Agric. Food Chem. 1996, 44, 3799–3806. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Higashiguchi, S.; Koketsu, M.; Juneja, L.R.; Kim, M.; Yamamoto, T. A structural phase of heat-denatured lysozyme with novel antimicrobial action. J. Agric. Food Chem. 1996, 44, 1416–1423. [Google Scholar] [CrossRef]
- Touch, V.; Hayakawa, S.; Fukada, K.; Aratani, Y.; Sun, Y. Preparation antimicrobial reduced lysozyme compatible in Food Applications. J. Agric. Food Chem. 2003, 53, 5154–5161. [Google Scholar] [CrossRef] [PubMed]
- Lesnierowski, G.; Kijowski, J.; Cegielska-Radziejewska, R. Ultrafiltration modified chicken egg white lysozyme and its antibacterial action. Int. J. Food Sci. Technol. 2009, 44, 305–311. [Google Scholar] [CrossRef]
- Abdou, A.M.; Higashiguchi, S.; Aboueleinin, A.M.; Kim, M.; Ibrahim, H.R. Antimicrobial peptides derived from hen egg lysozyme with inhibitory effect against Bacillus species. Food Cont. 2007, 18, 173–178. [Google Scholar] [CrossRef]
- Aminlari, L.; Hashemi, M.M.; Aminlari, M. Modified Lysozymes as Novel Broad Spectrum Natural Antimicrobial Agents in Foods. J. Food Sci. 2014, 79, R1077–R1090. [Google Scholar] [CrossRef] [PubMed]
- Nakimbugwe, D.; Masschalck, B.; Atanassova, M.; Zewdie-Bosüner, A.; Michiels, C.W. Comparison of bactericidal activity of six lysozymes at atmospheric pressure and under high hydrostatic pressure. Int. J. Food Microbiol. 2006, 108, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Mine, Y.; Ma, F.P.; Lauriau, S. Antimicrobial peptides released by enzymatic hydrolysis of hen egg white lysozyme. J. Agric. Food Chem. 2004, 58, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, W.; Lucio, A.; Gaibor, J.; Morales, D.; Vásquez, G. Isolation of antibacterial hydrolysates from hen egg white lysozyme and identification of antibacterial peptides. J. Med. Food. 2018, 21, 808–818. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, W.; Ramos, M. Identification of antimicrobial peptides of native and heated hydrolysates from hen egg white lysozyme. J. Med. Food. 2018, 21, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Touch, V.; Hayakawa, S.; Saitoh, K. Relationships between conformational changes and antimicrobial activity of lysozyme upon reduction of its disulfides bonds. Food Chem. 2004, 84, 421–428. [Google Scholar] [CrossRef]
- Shugar, D. The measurement of lysozyme activity and the ultra-violet inactivation of lysozyme. Biochim. Biophys. Acta 1952, 8, 302–309. [Google Scholar] [CrossRef]
- Masschalck, B.; Deckers, D.; Michiels, C.W. Lytic and nonlytic mechanism of inactivation of Gram-positive bacteria by lysozyme under atmospheric and high hydrostatic pressure. J. Food Prot. 2002, 65, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, V.; Rodríguez, N.; Amils, R. Immunocytochemical analysis of the subcellular distribution of ferritin in imperata cylindrica (L.) Raeuschel, an iron hyperaccumulator plant. Acta Histochem. 2012, 114, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Jollès, P.; Jollès, J. What’s new in lysozyme research? Mol. Cell Biochem. 1984, 63, 165–189. [Google Scholar] [CrossRef] [PubMed]
- Cegielska-Radziejewska, R.; Leśnierowski, G.; Kijowski, J. Antibacterial activity of lysozyme modified by the membrane technique. Electr. J. Pol. Agric. Univ. 2003, 6, 2. [Google Scholar]
- Monaco, L.A.; Rosenberger, F. Growth and etching kinetics of tetragonal lysozyme. J. Cryst. Growth 1993, 129, 465–484. [Google Scholar] [CrossRef]
- Tomizawa, H.; Yamada, H.; Tanigawa, K.; Imoto, T. Stabilization of lysozyme against irreversible inactivation by suppression of chemical reactions. J. Biochem. 1995, 117, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Nakada, T.; Sazaki, G.; Miyashita, S.; Durbin, S.D.; Komatsu, H. Direct AFM observations of impurity effects on a lysozyme crystal. J. Cryst. Growth. 1999, 196, 503–510. [Google Scholar] [CrossRef]
- Leśnierowski, G.; Cegielska-Radziejewska, R. Potential possibilities of production, modification and practical application of lysozyme. Acta Sci. Pol. Technol. Aliment. 2012, 11, 223–230. [Google Scholar] [PubMed]
- Siwicki, A.K.; Klein, P.; Morand, M.; Kiczka, W.; Studnicka, M. Immunostimulatory effects of dimerized lysozyme (KLP-602) on the nonspecific defense mechanisms and protection against furunculosis in salmonids. Vet. Immunol. Immunopathol. 1998, 62, 369–378. [Google Scholar] [CrossRef]
- Kiczka. From lysozyme monomer to lysozyme dimmer. In Proceedings of the 18th Buiatrics Congress, Bologna, Italy, 29 August–2 September, 1994; pp. 897–900. [Google Scholar]
- Rymuszka, A.; Studnicka, M.; Siwicki, A.K.; Sierosławska, A.; Bownik, A. The immunomodulatory effects of the dimer of lysozyme (KLP-602) in carp (Cyprinus carpio L) in vivo study. Ecotoxicol. Environ. Saf. 2015, 61, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Matsuzaki, T.; Aoki, T. Genetic evidence that antibacterial activity of lysozyme is independent of its catalytic function. FEBS Lett. 2001, 506, 27–32. [Google Scholar] [CrossRef]
- Ibrahim, H.R. On the novel catalytically-independent antimicrobial function of hen egg-white lysozyme: A conformation-dependent activity. Food-Nahrung 1998, 42, 187–193. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Stączek, S.; Mak, P.; Piersiak, T.; Skrzypiec, K.; Cytryńska, M. The effect of Galleria mellonella apolipophorin III on yeasts and filamentous fungi. J. Insect Physiol. 2012, 58, 164–177. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (2 mg/mL) | E. coli | S. carnosus |
|---|---|---|
| Heat-Treatment at pH 6.0 | ||
| N-HEWL | 1.6 ± 0.15 a | 7.8 ± 0.12 a |
| HEWL/80 °C | 4.3 ± 0.22 b | 4.9 ± 0.24 b |
| HEWL/90 °C | 3.8 ± 0.11 c | 5.2 ± 0.20 b |
| HEWL/95 °C | 3.9 ± 0.18 c | 5.2 ± 0.15 b |
| HEWL/120 °C | 3.7 ± 0.13 c | 5.6 ± 0.35 c |
| Heat-Treatment at pH 7.0 | ||
| N-HEWL | 0.0 | 8.0 ± 0.32 a |
| HEWL/80 °C | 0.3 ± 0.24 a | 3.2 ± 0.23 b |
| HEWL/90 °C | 0.3 ± 0.28 a | 3.3 ± 0.31 b |
| HEWL/95 °C | 0.3 ± 0.30 a | 4.7 ± 0.25 c |
| HEWL/120 °C | 0.5 ± 0.16 a | 5.9 ± 0.40 d |
| DTT-Treatment | ||
| HEWL/DTT 0 h | 0.0 | 8.0 ± 0.16 a |
| HEWL/DTT 1.0 h | 3.3 ± 0.23 a | 5.2 ± 0.50 b |
| HEWL/DTT 1.5 h | 5.5 ± 0.14 b | 3.9 ± 0.30 c |
| HEWL/DTT 2.0 h | 3.6 ± 0.12 c | 4.2 ± 0.31 d |
| HEWL/DTT 4.0 h | 3.6 ± 0.30 c | 4.4 ± 0.23 d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilcacundo, R.; Méndez, P.; Reyes, W.; Romero, H.; Pinto, A.; Carrillo, W. Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments. Sci. Pharm. 2018, 86, 48. https://doi.org/10.3390/scipharm86040048
Vilcacundo R, Méndez P, Reyes W, Romero H, Pinto A, Carrillo W. Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments. Scientia Pharmaceutica. 2018; 86(4):48. https://doi.org/10.3390/scipharm86040048
Chicago/Turabian StyleVilcacundo, Rubén, Pilar Méndez, Walter Reyes, Herman Romero, Adelita Pinto, and Wilman Carrillo. 2018. "Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments" Scientia Pharmaceutica 86, no. 4: 48. https://doi.org/10.3390/scipharm86040048
APA StyleVilcacundo, R., Méndez, P., Reyes, W., Romero, H., Pinto, A., & Carrillo, W. (2018). Antibacterial Activity of Hen Egg White Lysozyme Denatured by Thermal and Chemical Treatments. Scientia Pharmaceutica, 86(4), 48. https://doi.org/10.3390/scipharm86040048