
Anti-Obesity Effect of Auricularia delicate Involves Intestinal-Microbiota-Mediated Oxidative Stress Regulation in High-Fat-Diet-Fed Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. ADe Component Detection
2.1.1. Nutritional Composition Analysis
2.1.2. Mineral Analysis
2.2. Animal Experiments and Agent Administration Protocol
2.3. Biochemical Analysis
2.4. Histopathological Analysis and Immunohistochemical Examination
2.5. Intestinal Microbiota Analysis
2.6. Lipidome Analysis
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Composition of ADe
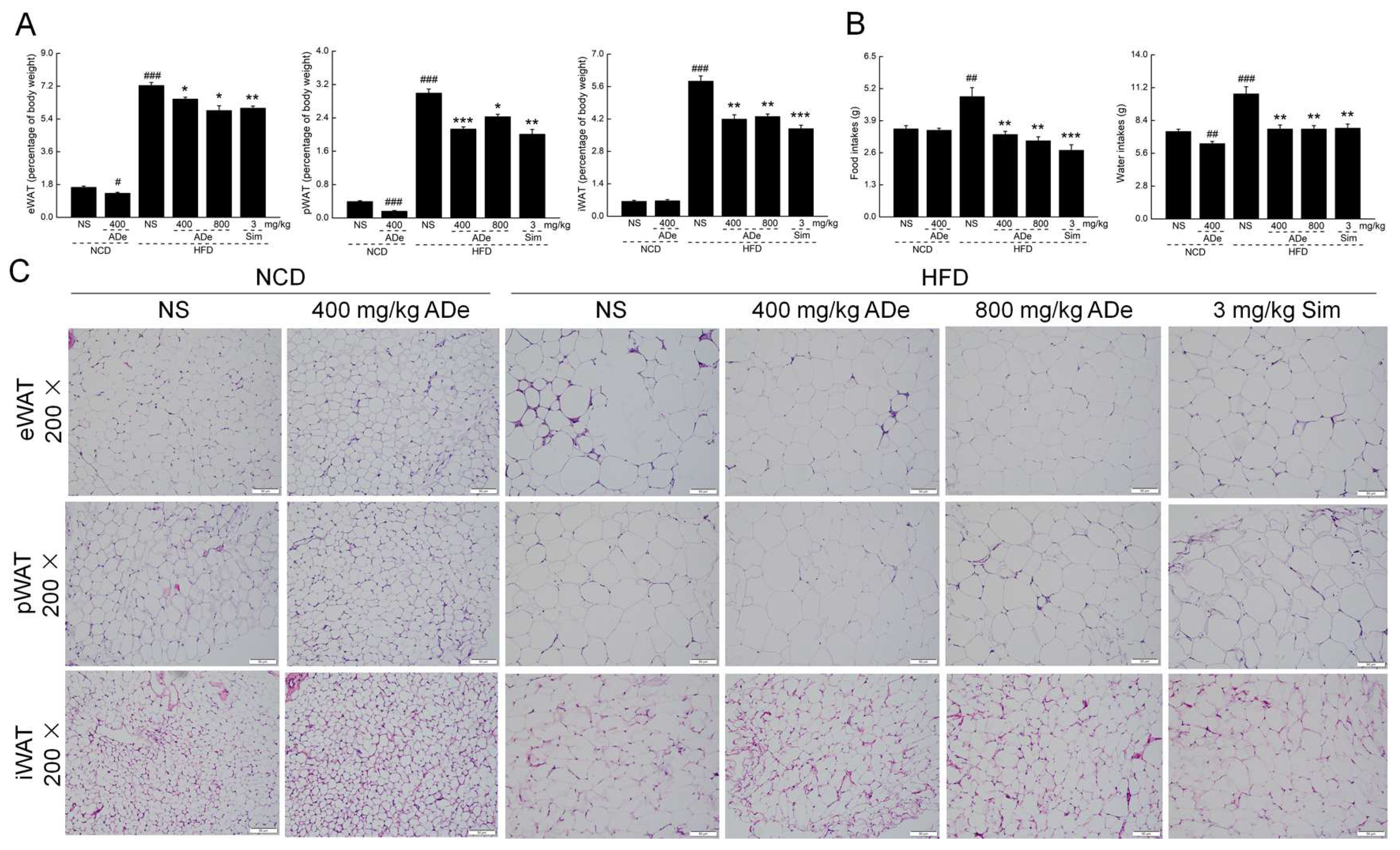
3.2. The Anti-Obesity Effect of ADe
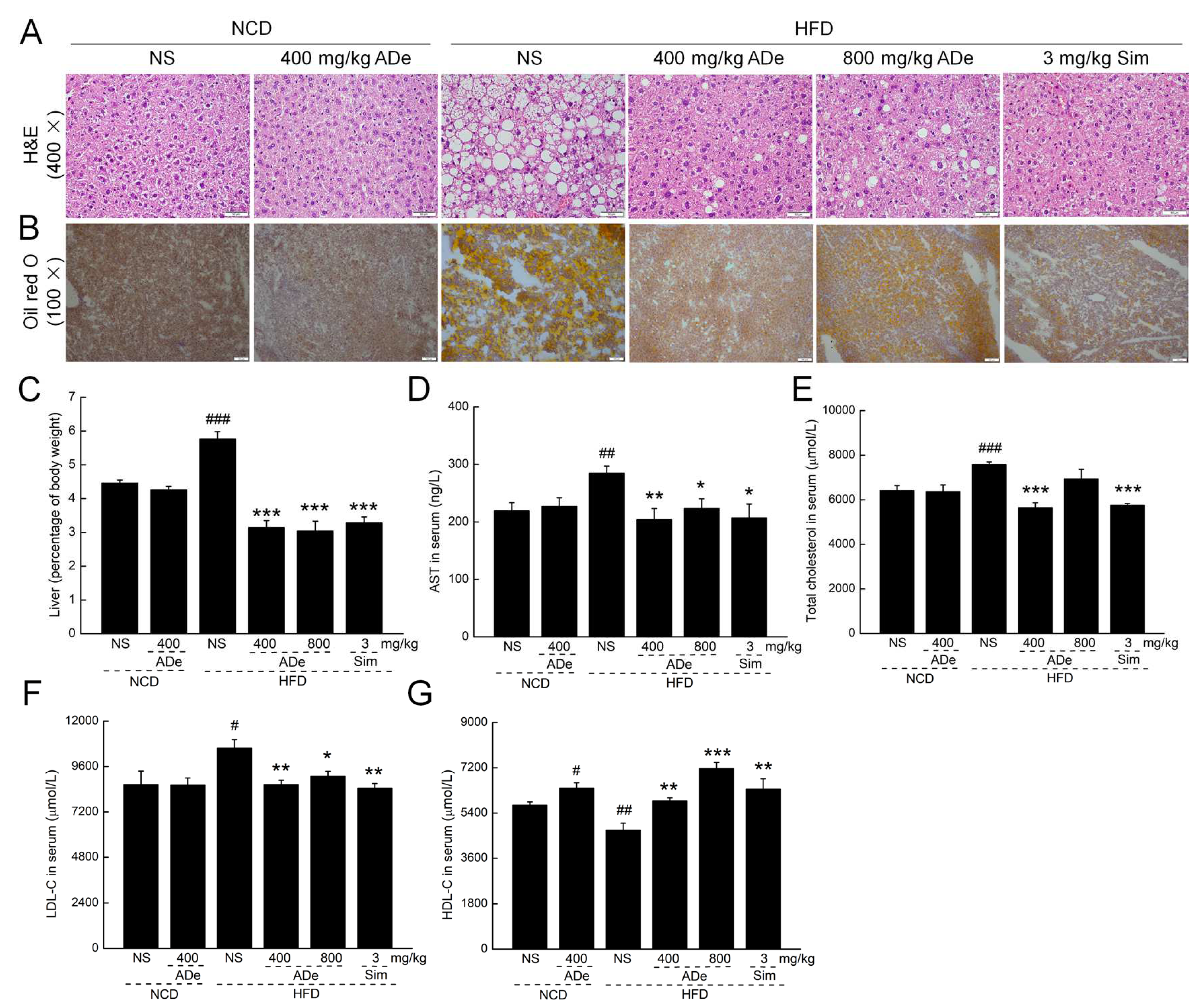
3.3. ADe Treatment Regulated Blood Lipids and Alleviated Hepatic Steatosis
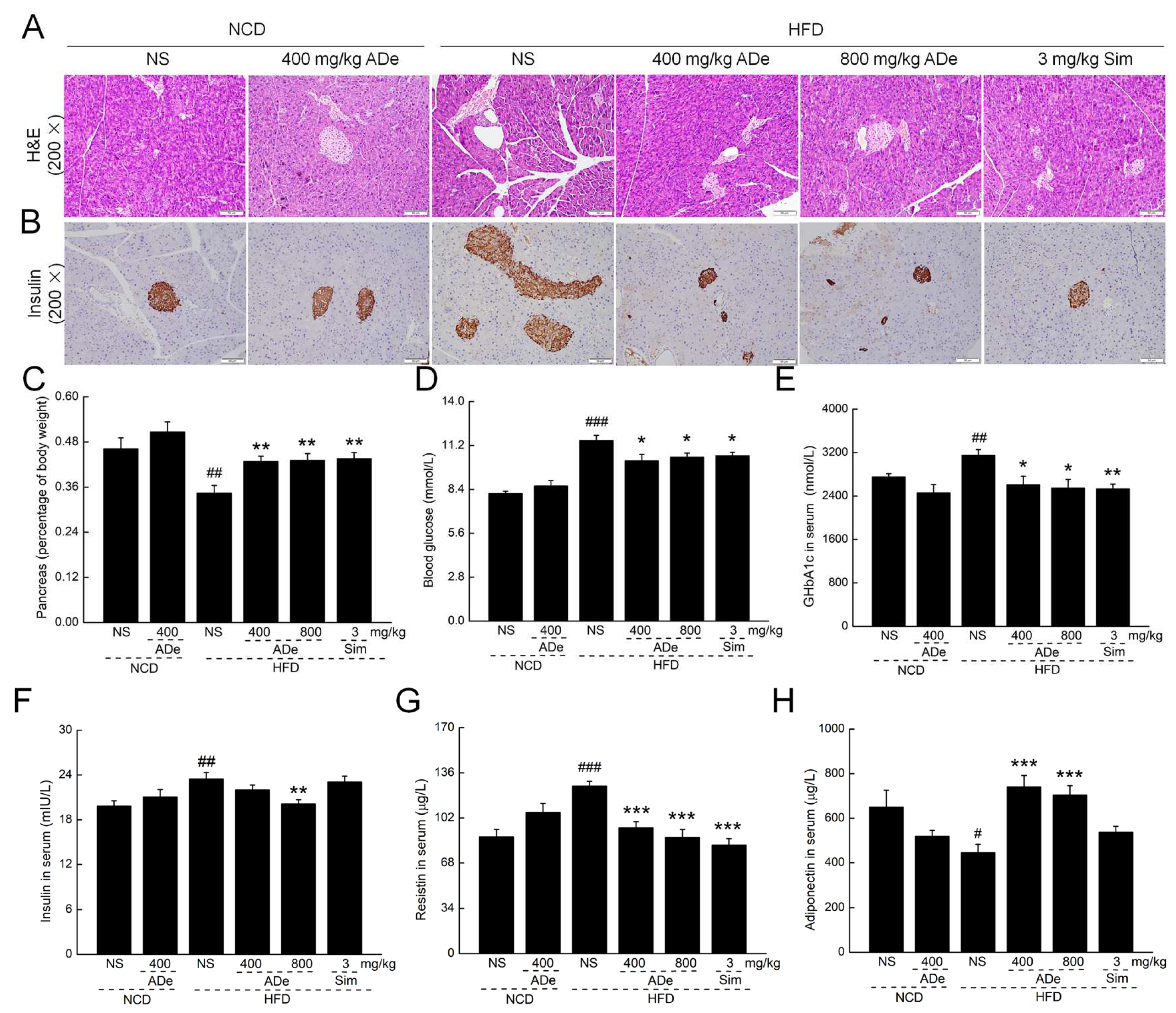
3.4. ADe Protected Pancreatic Function
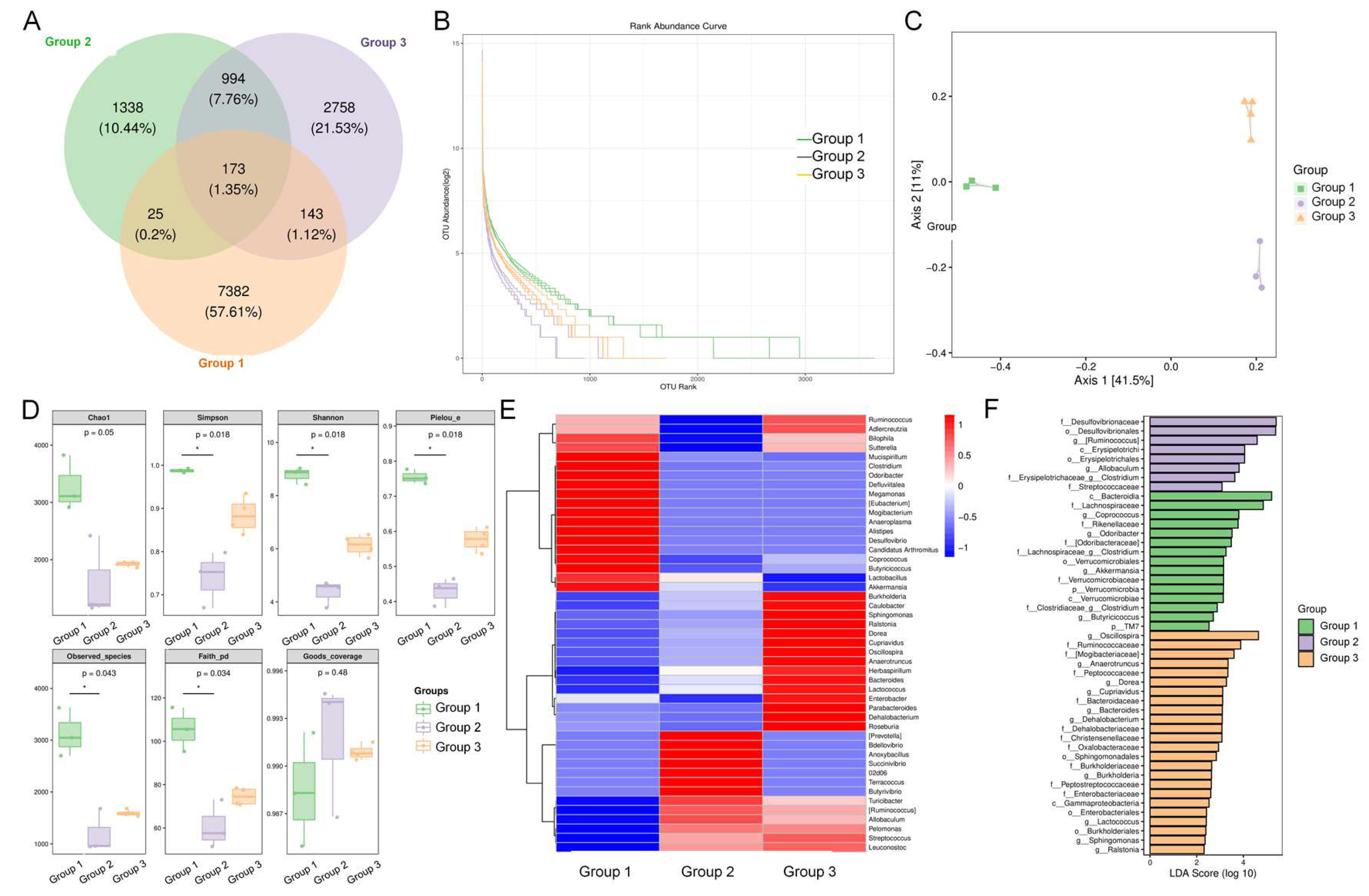
3.5. ADe Regulated the Intestinal Microbiota
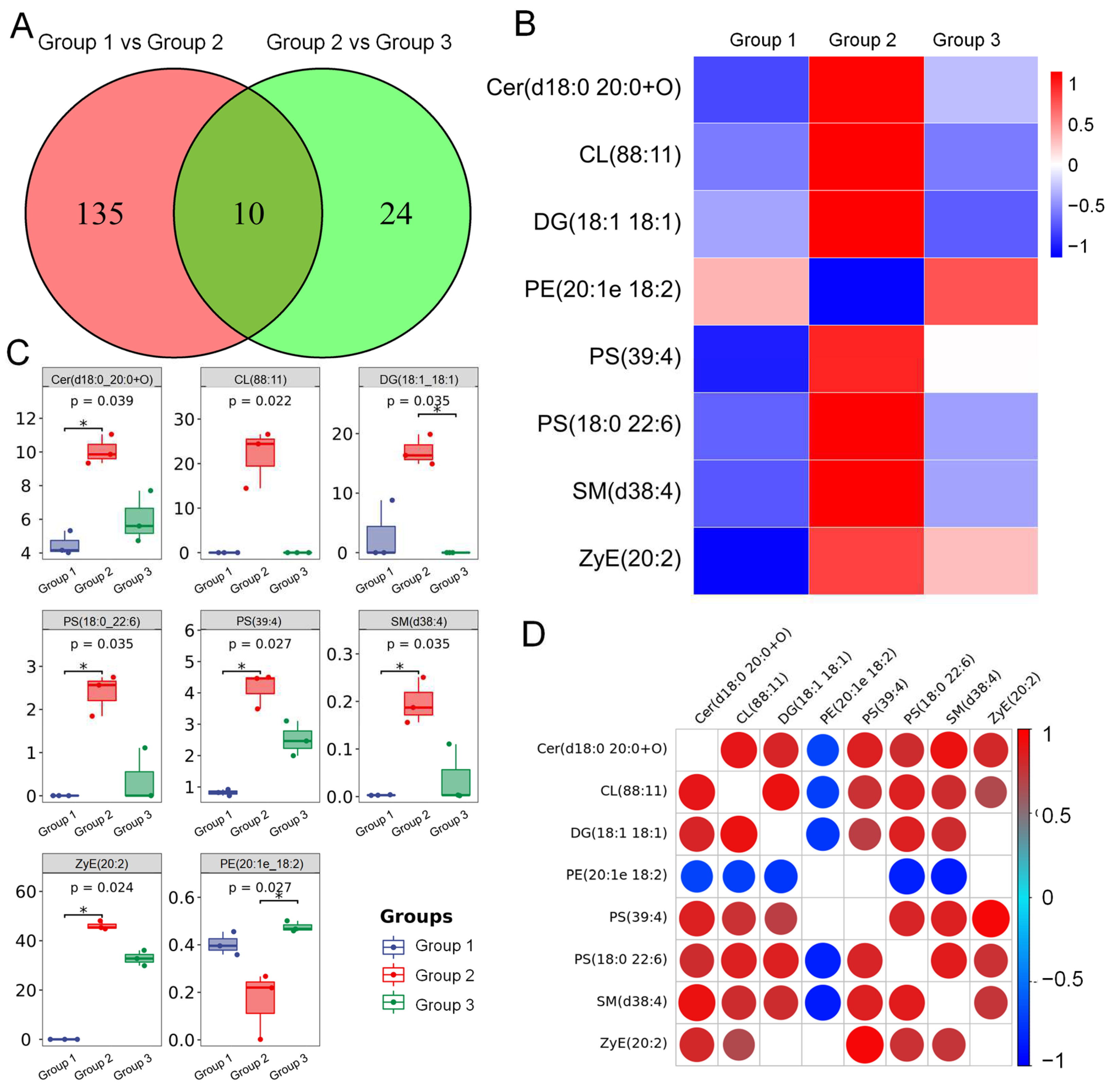
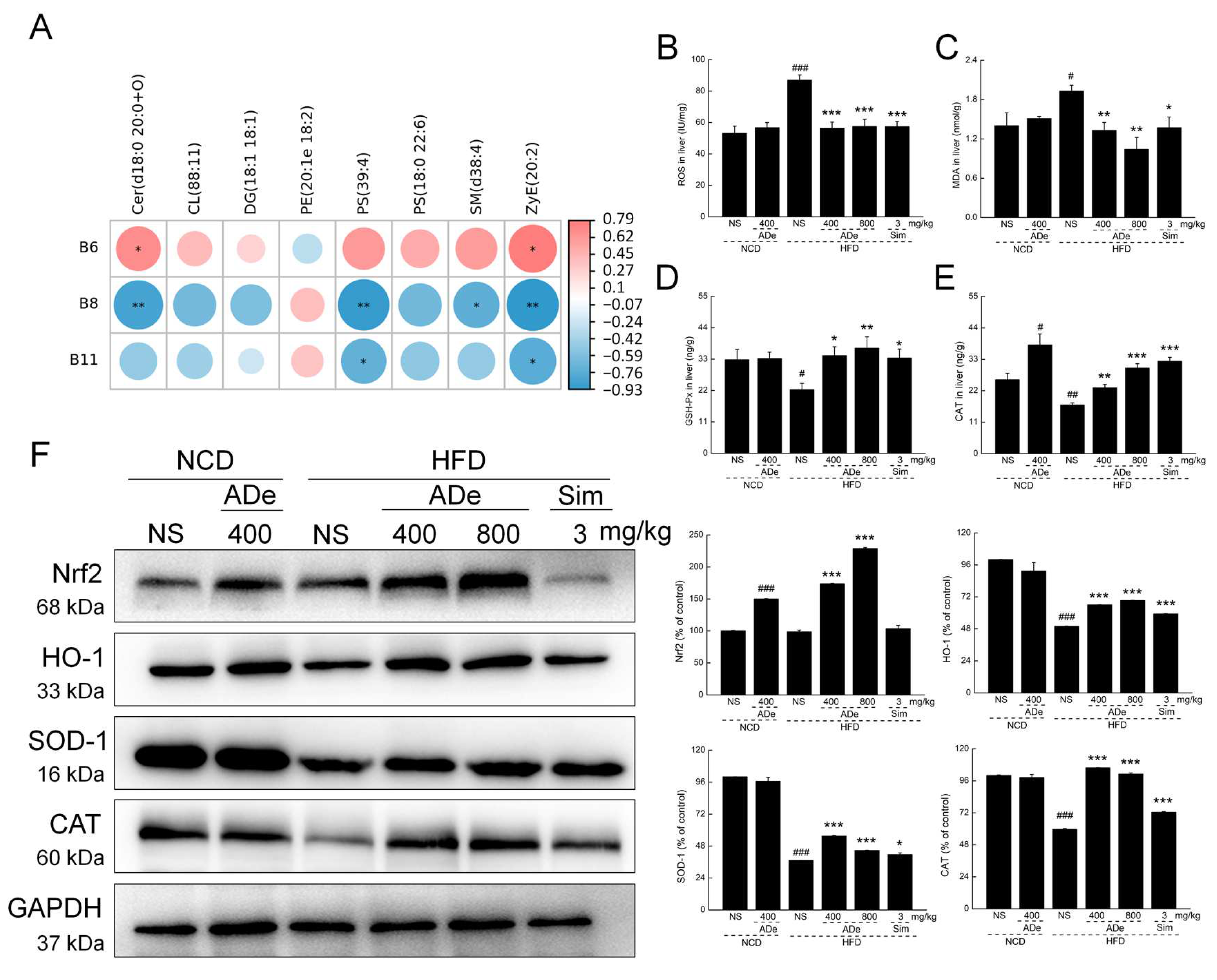
3.6. ADe Regulated the Metabolism of Lipids Related to the Nrf2/HO-1 Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Said, M.M.; Sandra, S.; Jaime, L.; Ignacio, B. Systems and WBANs for Controlling Obesity. J. Healthc. Eng. 2018, 2018, 1564748. [Google Scholar] [CrossRef]
- Pan, X.F.; Wang, L.M.; Pan, A. Epidemiology and determinants of obesity in China. Lancet Diabetes Endocrinol. 2021, 9, 373–392. [Google Scholar] [CrossRef] [PubMed]
- Despres, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.L.; Jiang, W.; Huang, W.L.; Lin, Y.; Chan, F.K.L.; Ng, S.C. Gut microbiota in patients with obesity and metabolic disorders—A systematic review. Genes Nutr. 2022, 17, 18. [Google Scholar] [CrossRef] [PubMed]
- Sjostrom, L.; Narbro, K.; Sjostrom, D.; Karason, K.; Larsson, B.; Wedel, H.; Lystig, T.; Sullivan, M.; Bouchard, C.; Carlsson, B.; et al. Effects of bariatric surgery on mortality in Swedish obese subjects. N. Engl. J. Med. 2007, 357, 741–752. [Google Scholar] [CrossRef]
- Kassir, R.; Debs, T.; Blanc, P.; Gugenheim, J.; Ben Amor, I.; Boutet, C.; Tiffet, O. Complications of bariatric surgery: Presentation and emergency management. Int. J. Surg. 2016, 27, 77–81. [Google Scholar] [CrossRef]
- Buchholz, T.; Melzig, M.F. Polyphenolic Compounds as Pancreatic Lipase Inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef]
- Aagaard, L.; Hallgreen, C.E.; Hansen, E.H. Serious adverse events reported for antiobesity medicines: Postmarketing experiences from the EU adverse event reporting system EudraVigilance. Int. J. Obes. 2016, 40, 1742–1747. [Google Scholar] [CrossRef]
- Seo, Y.-G. Side Effects Associated with Liraglutide Treatment for Obesity as Well as Diabetes. J. Obes. Metab. Syndr. 2021, 30, 12–19. [Google Scholar] [CrossRef]
- Toth, P.P.; Davidson, M.H. Simvastatin plus ezetimibe: Combination therapy for the management of dyslipidaemia. Expert Opin. Pharmacother. 2005, 6, 131–139. [Google Scholar] [CrossRef]
- Zhang, S.R.; Li, H.; Yuan, L.; Zhang, J.; Han, L.; Liu, R.N.; Wu, R.; Shi, Y.; Mushtaq, N.; Ullah, S.; et al. Molecular characterization of gut microbiota in high-lipid diet-induced hyperlipidemic rats treated with simvastatin. Int. J. Mol. Med. 2020, 45, 1601–1615. [Google Scholar] [CrossRef]
- Andoh, A.; Imaeda, H. Comparison of the gut microbial community between obese and lean peoples using 16S gene sequencing in a Japanese population. J. Gastroenterol. Hepatol. 2016, 31, 155. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, H.J.; Li, K.L.; Shi, Y.; Guo, X.F.; Wang, L.; Li, D. Sandalwood seed oil improves insulin sensitivity in high-fat/high-sucrose diet-fed rats associated with altered intestinal microbiota and its metabolites. Food Funct. 2021, 12, 9739–9749. [Google Scholar] [CrossRef]
- Qian, L.M.; Huang, J.M.; Qin, H.L. Probiotics and dietary intervention modulate the colonic mucosa-associated microbiota in high-fat diet populations. Turk. J. Gastroenterol. 2020, 31, 295. [Google Scholar] [CrossRef]
- Nogal, A.; Louca, P.; Zhang, X.Y.; Wells, P.M.; Steves, C.J.; Spector, T.D.; Falchi, M.; Valdes, A.M.; Menni, C. Circulating Levels of the Short-Chain Fatty Acid Acetate Mediate the Effect of the Gut Microbiome on Visceral Fat. Front. Microbiol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Gonzalez-Bosch, C.; Boorman, E.; Zunszain, P.A.; Mann, G.E. Short-chain fatty acids as modulators of redox signaling in health and disease. Redox Biol. 2021, 47, 11. [Google Scholar] [CrossRef]
- Vallianou, N.; Christodoulatos, G.S.; Karampela, I.; Tsilingiris, D.; Magkos, F.; Stratigou, T.; Kounatidis, D.; Dalamaga, M. Understanding the Role of the Gut Microbiome and Microbial Metabolites in Non-Alcoholic Fatty Liver Disease: Current Evidence and Perspectives. Biomolecules 2022, 12, 30. [Google Scholar] [CrossRef]
- Perez-Torres, I.; Castrejon-Tellez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 26. [Google Scholar] [CrossRef]
- Ryoo, I.G.; Kwak, M.K. Regulatory crosstalk between the oxidative stress-related transcription factor Nfe212/Nrf2 and mitochondria. Toxicol. Appl. Pharmacol. 2018, 359, 24–33. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Et Biophys. Acta-Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Yu, Z.; Shao, W.; Chiang, Y.; Foltz, W.; Zhang, Z.; Ling, W.; Fantus, I.G.; Jin, T. Oltipraz upregulates the nuclear respiratory factor 2 alpha subunit (NRF2) antioxidant system and prevents insulin resistance and obesity induced by a high-fat diet in C57BL/6J mice. Diabetologia 2011, 54, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Spim, S.R.V.; de Oliveira, B.; Leite, F.G.; Gerenutti, M.; Grotto, D. Effects of Lentinula edodes consumption on biochemical, hematologic and oxidative stress parameters in rats receiving high-fat diet. Eur. J. Nutr. 2017, 56, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hao, J.; Liu, Z.J.; Ma, X.T.; Feng, Y.X.; Teng, L.R.; Li, Y.; Wang, D. Anti-obesity effects of Grifola frondosa through the modulation of lipid metabolism via ceramide in mice fed a high-fat diet. Food Funct. 2021, 12, 6725–6739. [Google Scholar] [CrossRef]
- Looney, B. Systematics of the genus Auricularia with an emphasis on species from the southeastern United States. North Am. Fungi 2013, 8, 1–25. [Google Scholar] [CrossRef]
- Li, X.; Jacob, M.S.; Zhang, B.; Xu, A. Research Progress on Auricularia delicata. J. Adv. Biol. Biotechnol. 2020, 23, 8–32. [Google Scholar] [CrossRef]
- Wangkheirakpam, S.D.; Joshi, D.D.; Leishangthem, G.D.; Biswas, D.; Deb, L. Hepatoprotective Effect of Auricularia delicata (Agaricomycetes) from India in Rats: Biochemical and Histopathological Studies and Antimicrobial Activity. Int. J. Med. Mushrooms 2018, 20, 213–225. [Google Scholar] [CrossRef]
- Hu, H.W.; Teng, X.; Zhang, S.S.; Liu, T.T.; Li, X.; Wang, D.W. Structural Characteristics, Rheological Properties, and Antioxidant Activity of Novel Polysaccharides from “Deer Tripe Mushroom”. J. Food Qual. 2021, 2021, 12. [Google Scholar] [CrossRef]
- Zhang, L.P.; Meng, B.; Li, L.Z.; Wang, Y.Z.; Zhang, Y.Z.; Fang, X.X.; Wang, D. Boletus aereusprotects against acute alcohol-induced liver damage in the C57BL/6 mouse via regulating the oxidative stress-mediated NF-kappa B pathway. Pharm. Biol. 2020, 58, 905–914. [Google Scholar] [CrossRef]
- Chen, C.; Li, S.X.; Wang, S.M.; Liang, S.W. Investigation into the anti-thrombosis effect and contents of total saponins and flavonoids in the bioactive fraction of Naodesheng prescription. J Ethnopharmacol 2012, 144, 208–212. [Google Scholar] [CrossRef]
- Ajanal, M.; Gundkalle, M.B.; Nayak, S.U. Estimation of total alkaloid in Chitrakadivati by UV-Spectrophotometer. Anc. Sci. Life 2012, 31, 198–201. [Google Scholar] [CrossRef]
- Han, S.N.; Yang, W.H.; Yin, J.J.; Tao, H.L.; Zhang, L.R. Drug Treatment of Hyperlipidemia in Chinese Patients: Focus on the Use of Simvastatin and Ezetimibe Alone and in Combination. Am. J. Cardiovasc. Drugs 2019, 19, 237–247. [Google Scholar] [CrossRef]
- Wang, J.; Teng, L.R.; Liu, Y.G.; Hu, W.J.; Chen, W.Q.; Hu, X.; Wang, Y.W.; Wang, D. Studies on the Antidiabetic and Antinephritic Activities of Paecilomyces hepiali Water Extract in Diet-Streptozotocin-Induced Diabetic Sprague Dawley Rats. J. Diabetes Res. 2016, 2016, 10. [Google Scholar] [CrossRef]
- Takata, T.; Sakasai-Sakai, A.; Takino, J.; Takeuchi, M. Evidence for Toxic Advanced Glycation End-Products Generated in the Normal Rat Liver. Nutrients 2019, 11, 22. [Google Scholar] [CrossRef]
- Ling, Z.X.; Liu, X.; Jia, X.Y.; Cheng, Y.W.; Luo, Y.Q.; Yuan, L.; Wang, Y.Z.; Zhao, C.N.; Guo, S.; Li, L.; et al. Impacts of infection with different toxigenic Clostridium difficile strains on faecal microbiota in children. Sci. Rep. 2014, 4, 10. [Google Scholar] [CrossRef]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Le Bloc’h, J.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef]
- Titchenell, P.M.; Lazar, M.A.; Birnbaum, M.J. Unraveling the Regulation of Hepatic Metabolism by Insulin. Trends Endocrinol. Metab. 2017, 28, 497–505. [Google Scholar] [CrossRef]
- Anderson, J.W.; Baird, P.; Davis, R.H.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health benefits of dietary fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef]
- Choi, M.K.; Song, H.J.; Paek, Y.J.; Lee, H.J. Gender Differences in the Relationship between Vitamin C and Abdominal Obesity. Int. J. Vitam. Nutr. Res. 2013, 83, 377–384. [Google Scholar] [CrossRef]
- Nahra, R.; Wang, T.; Gadde, K.M.; Oscarsson, J.; Stumvoll, M.; Jermutus, L.; Hirshberg, B.; Ambery, P. Effects of Cotadutide on Metabolic and Hepatic Parameters in Adults With Overweight or Obesity and Type 2 Diabetes: A 54-Week Randomized Phase 2b Study. Diabetes Care 2021, 44, 1433–1442. [Google Scholar] [CrossRef]
- Veghari, G.; Sedaghat, M.; Maghsodlo, S.; Banihashem, S.; Moharloei, P.; Angizeh, A.; Tazik, E.; Moghaddami, A.; Joshaghani, H. The association between abdominal obesity and serum cholesterol level. Int. J. Appl. Basic Med. Res. 2015, 5, 83–86. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative Stress in Obesity: A Critical Component in Human Diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.C.; Kang, J.H.; Guan, Y.F. The mechanisms linking adiposopathy to type 2 diabetes. Front. Med. 2013, 7, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Bazotte, R.B.; Silva, L.G.; Schiavon, F.P.M. Insulin resistance in the liver: Deficiency or excess of insulin? Cell Cycle 2014, 13, 2494–2500. [Google Scholar] [CrossRef] [PubMed]
- Acquarone, E.; Monacelli, F.; Borghi, R.; Nencioni, A.; Odetti, P. Resistin: A reappraisal. Mech. Ageing Dev. 2019, 178, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, M.; Asadi, N.; Pouralborz, Y.; Ghodrat, M.; Habibi, S. A Review on Glycosylated Hemoglobin in Polycystic Ovary Syndrome. J. Pediatr. Adolesc. Gynecol. 2016, 29, 562–566. [Google Scholar] [CrossRef]
- Christou, G.A.; Kiortsis, D.N. Adiponectin and lipoprotein metabolism. Obes. Rev. 2013, 14, 939–949. [Google Scholar] [CrossRef]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 27. [Google Scholar] [CrossRef]
- Liu, R.X.; Hong, J.; Xu, X.Q.; Feng, Q.; Zhang, D.Y.; Gu, Y.Y.; Shi, J.; Zhao, S.Q.; Liu, W.; Wang, X.K.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859. [Google Scholar] [CrossRef]
- Miquel, S.; Martin, R.; Rossi, O.; Bermudez-Humaran, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, D.F.; He, Y.; Li, Y.H.; Yang, Z.Y.; Zhao, X.L.; Liu, Y.H.; Wang, Y.; Sun, J.; Feng, X.; et al. Discrepant gut microbiota markers for the classification of obesity-related metabolic abnormalities. Sci. Rep. 2019, 9, 10. [Google Scholar] [CrossRef]
- Fang, S.T.; Qin, T.; Yu, T.; Zhang, G.X. Improvement of the Gut Microbiota In Vivo by a Short-Chain Fatty Acids-Producing Strain Lactococcus garvieae CF11. Processes 2022, 10, 12. [Google Scholar] [CrossRef]
- McNabney, S.M.; Henagan, T.M. Short Chain Fatty Acids in the Colon and Peripheral Tissues: A Focus on Butyrate, Colon Cancer, Obesity and Insulin Resistance. Nutrients 2017, 9, 28. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Mao, G.X.; Zuo, J.H.; Li, S.J.; Yang, Y.; Thring, R.W.; Wu, M.J.; Tong, H.B. Sargassum fusiforme fucoidan alleviates diet-induced insulin resistance by inhibiting colon-derived ceramide biosynthesis. Food Funct. 2021, 12, 8440–8453. [Google Scholar] [CrossRef]
- Zhang, C.N.; Abdo, A.A.A.; Kaddour, B.; Wu, Q.H.; Xin, L.; Li, X.T.; Fan, G.S.; Teng, C. Xylan-oligosaccharides ameliorate high fat diet induced obesity and glucose intolerance and modulate plasma lipid profile and gut microbiota in mice. J. Funct. Foods 2020, 64, 8. [Google Scholar] [CrossRef]
- Cai, H.Y.; Wen, Z.G.; Xu, X.; Wang, J.X.; Li, X.; Meng, K.; Yang, P.L. Serum Metabolomics Analysis for Biomarkers of Lactobacillus plantarum FRT4 in High-Fat Diet-Induced Obese Mice. Foods 2022, 11, 14. [Google Scholar] [CrossRef]
- Wu, T.; Sun, M.Z.; Liu, R.; Sui, W.J.; Zhang, J.J.; Yin, J.J.; Fang, S.G.; Zhu, J.G.; Zhang, M. Bifidobacterium longum subsp. longum Remodeled Roseburia and Phosphatidylserine Levels and Ameliorated Intestinal Disorders and liver Metabolic Abnormalities Induced by High-Fat Diet. J. Agric. Food Chem. 2020, 68, 4632–4640. [Google Scholar] [CrossRef]
- Hanamatsu, H.; Ohnishi, S.; Sakai, S.; Yuyama, K.; Mitsutake, S.; Takeda, H.; Hashino, S.; Igarashi, Y. Altered levels of serum sphingomyelin and ceramide containing distinct acyl chains in young obese adults. Nutr. Diabetes 2014, 4, 7. [Google Scholar] [CrossRef]
- Perez-Canamas, A.; Benvegnu, S.; Rueda, C.B.; Rabano, A.; Satrustegui, J.; Ledesma, M.D. Sphingomyelin-induced inhibition of the plasma membrane calcium ATPase causes neurodegeneration in type A Niemann-Pick disease. Mol. Psychiatry 2017, 22, 711–723. [Google Scholar] [CrossRef]
- Joseph, J.A.; Denisova, N.; Fisher, D.; Bickford, P.; Prior, R.; Cao, G. Age-related neurodegeneration and oxidative stress: Putative nutritional intervention. Neurol. Clin. 1998, 16, 747–755. [Google Scholar] [CrossRef]
- Houstis, N.; Rosen, E.D.; Lander, E.S. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006, 440, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Li, S.R.; Eguchi, N.; Lau, H.; Ichii, H. The Role of the Nrf2 Signaling in Obesity and Insulin Resistance. Int. J. Mol. Sci. 2020, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, P.; Pena, A.; Vettorazzi, A.; Fabra, M.J.; Martinez-Abad, A.; Lopez-Rubio, A.; Pera, J.; Parlade, J.; Castellari, M.; Milagro, F.I.; et al. Grifola frondosa (Maitake) Extract Reduces Fat Accumulation and Improves Health Span in C. elegans through the DAF-16/FOXO and SKN-1/NRF2 Signalling Pathways. Nutrients 2021, 13, 20. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Contents (%) | Name | Contents (%) | |
|---|---|---|---|---|
| General nutritional compositions | Total sugar | 54.6 | Total triterpenoids (×10−1) | 0.2 |
| Reducing sugar (×10−1) | 25.8 | Mannitol (×10−2) | 0.6 | |
| Crude protein (×10−1) | 81.8 | Total phenol (×10−1) | 1.3 | |
| Total ash | 3.8 | Total saponins | 0.1 | |
| Crude fat | 2.6 | Total alkaloids (×10−2) | 1.2 | |
| Crude fiber | 12.4 | Total sterol (×10−2) | 38.5 | |
| Total flavonoids | UDa | |||
| Minerals | Calcium (Ca) (×10−2) | 22.8 | Magnesium (Mg) (×10−2) | 16.8 |
| Iron (Fe) (×10−2) | 1.6 | Potassium (K) (×10−1) | 11.5 | |
| Zinc (Zn) (×10−5) | 93.6 | Sodium (Na) (×10−6) | 6.8 | |
| Selenium (Se) (×10−6) | 10.3 | Manganese (Mn) (×10−5) | 57.5 | |
| Lead (Pb) (×10−6) | 56.5 | Cadmium (Cd) | UDb | |
| Arsenic (As) (×10−4) | 10.6 | Copper (Cu) (×10−5) | 29.8 | |
| Mercury (Hg) | UDb | Chromium (Cr) (×10−4) | 23.9 | |
| Vitamins | Vitamin A | UDc | Pyridoxine | UDf |
| Vitamin E (×10−5) | 36.1 | Pyridoxal (×10−5) | 84.7 | |
| Vitamin C (×10−3) | 81.1 | Pyridoxamine | UDf | |
| Vitamin B1 | UDd | Vitamin D2 (×10−4) | 5.6 | |
| Vitamin B2 | UDe | Vitamin D3 (×10−5) | 55.5 | |
| Vitamin B3 (×10−4) | 19.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhai, S.; Wang, R.; Kong, F.; Yang, A.; Wang, C.; Yu, H.; Li, Y.; Wang, D. Anti-Obesity Effect of Auricularia delicate Involves Intestinal-Microbiota-Mediated Oxidative Stress Regulation in High-Fat-Diet-Fed Mice. Nutrients 2023, 15, 872. https://doi.org/10.3390/nu15040872
Li L, Zhai S, Wang R, Kong F, Yang A, Wang C, Yu H, Li Y, Wang D. Anti-Obesity Effect of Auricularia delicate Involves Intestinal-Microbiota-Mediated Oxidative Stress Regulation in High-Fat-Diet-Fed Mice. Nutrients. 2023; 15(4):872. https://doi.org/10.3390/nu15040872
Chicago/Turabian StyleLi, Lanzhou, Siyu Zhai, Ruochen Wang, Fange Kong, Anhui Yang, Chunyue Wang, Han Yu, Yu Li, and Di Wang. 2023. "Anti-Obesity Effect of Auricularia delicate Involves Intestinal-Microbiota-Mediated Oxidative Stress Regulation in High-Fat-Diet-Fed Mice" Nutrients 15, no. 4: 872. https://doi.org/10.3390/nu15040872
APA StyleLi, L., Zhai, S., Wang, R., Kong, F., Yang, A., Wang, C., Yu, H., Li, Y., & Wang, D. (2023). Anti-Obesity Effect of Auricularia delicate Involves Intestinal-Microbiota-Mediated Oxidative Stress Regulation in High-Fat-Diet-Fed Mice. Nutrients, 15(4), 872. https://doi.org/10.3390/nu15040872