

The Screening and Mechanism of Influenza-Virus Sensitive MDCK Cell Lines for Influenza Vaccine Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Influenza Viruses
2.3. Construction of MDCK Cell Subclones by the Limit Dilution Method
2.4. Hemagglutination Titer Test
2.5. Screening of Influenza-Virus-Sensitive MDCK Subclonal Cells
2.6. Cell Proliferation Ability Detection by CCK-8
2.7. Cell Proliferation Activity Detection by Flow Cytometry
2.8. Gene Set Enrichment Analysis (GSEA)
3. Results
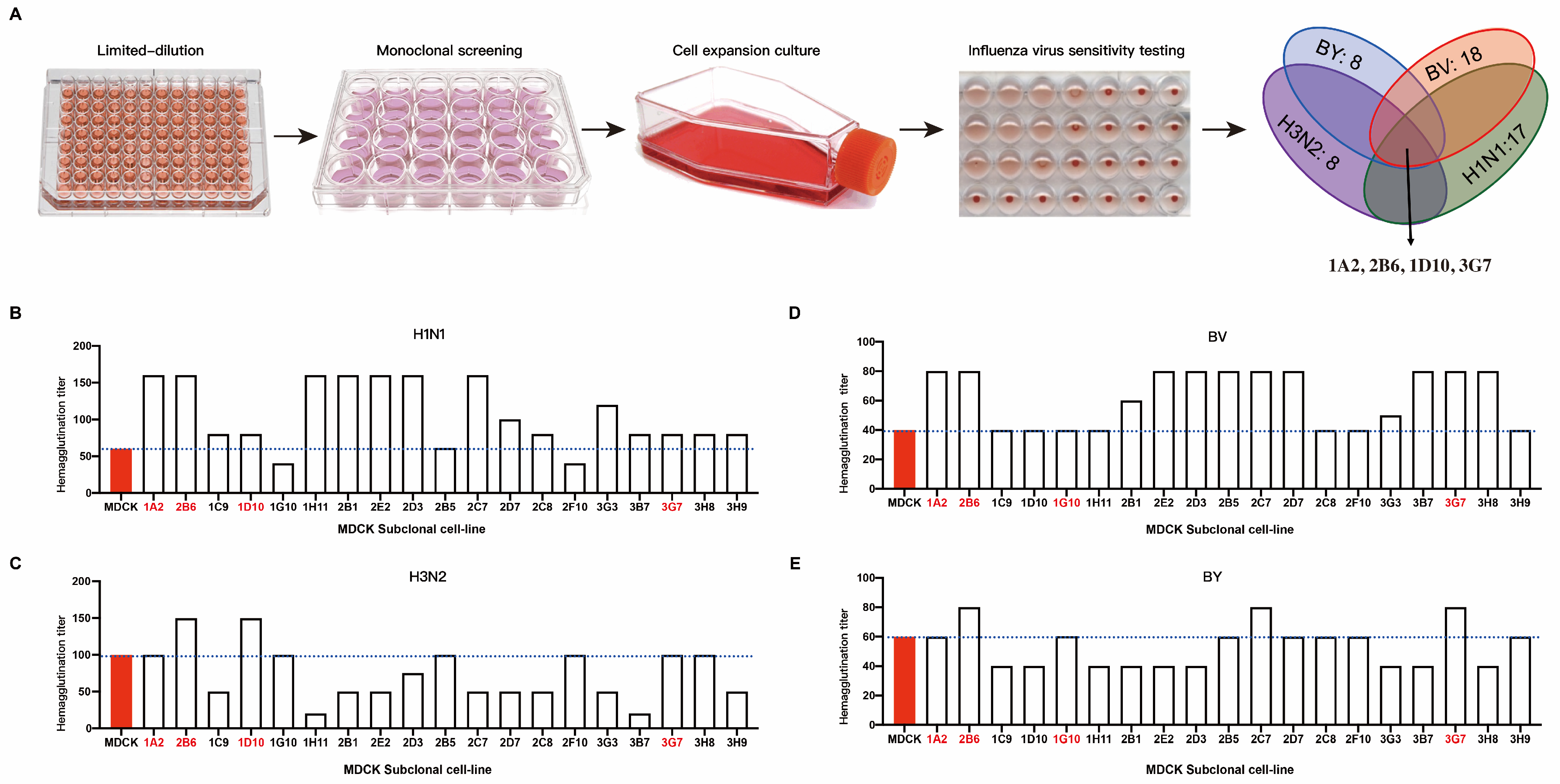
3.1. The First-Round Screening of Influenza-Virus-Sensitive MDCK Subclonal Cells
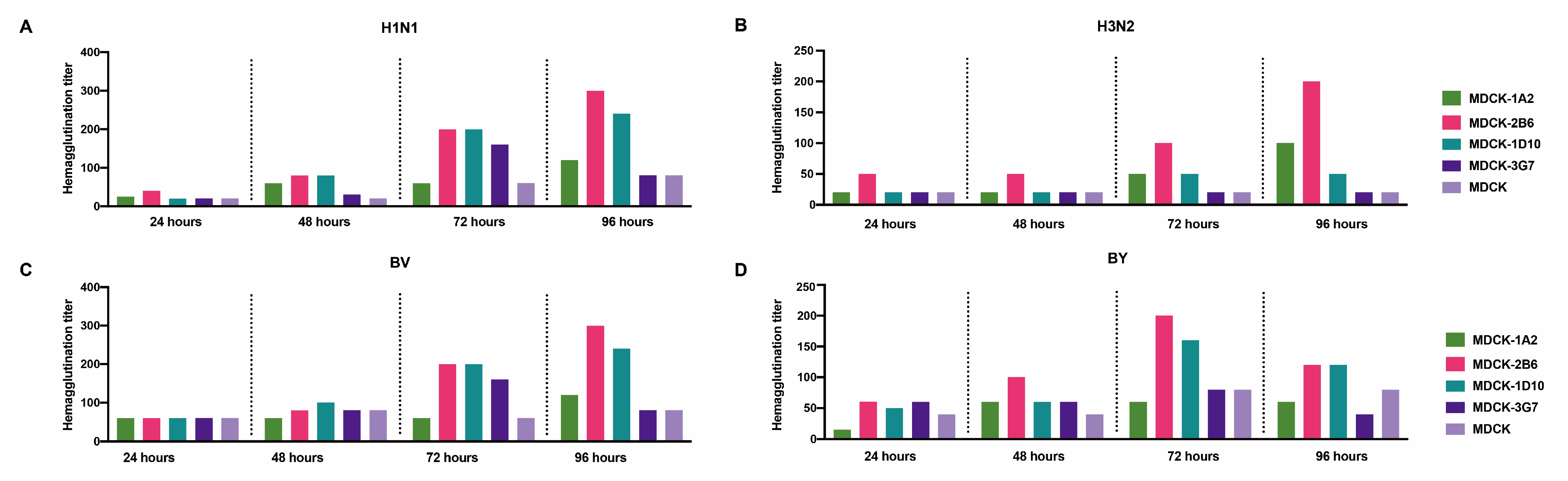
3.2. The Re-Screening of ATCC-MDCK Subclone Cells
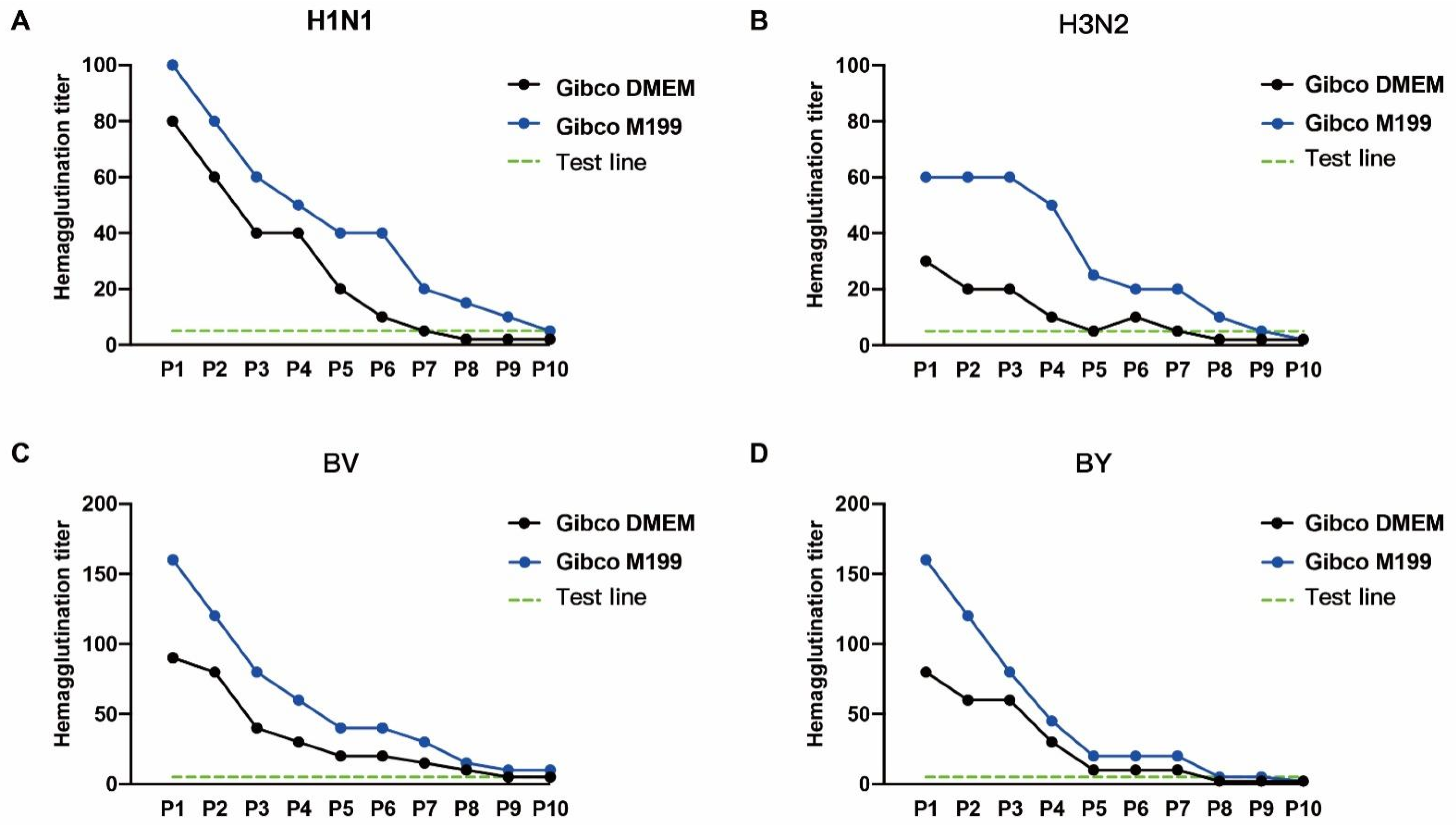
3.3. A Comparison of Influenza Virus Culture Media
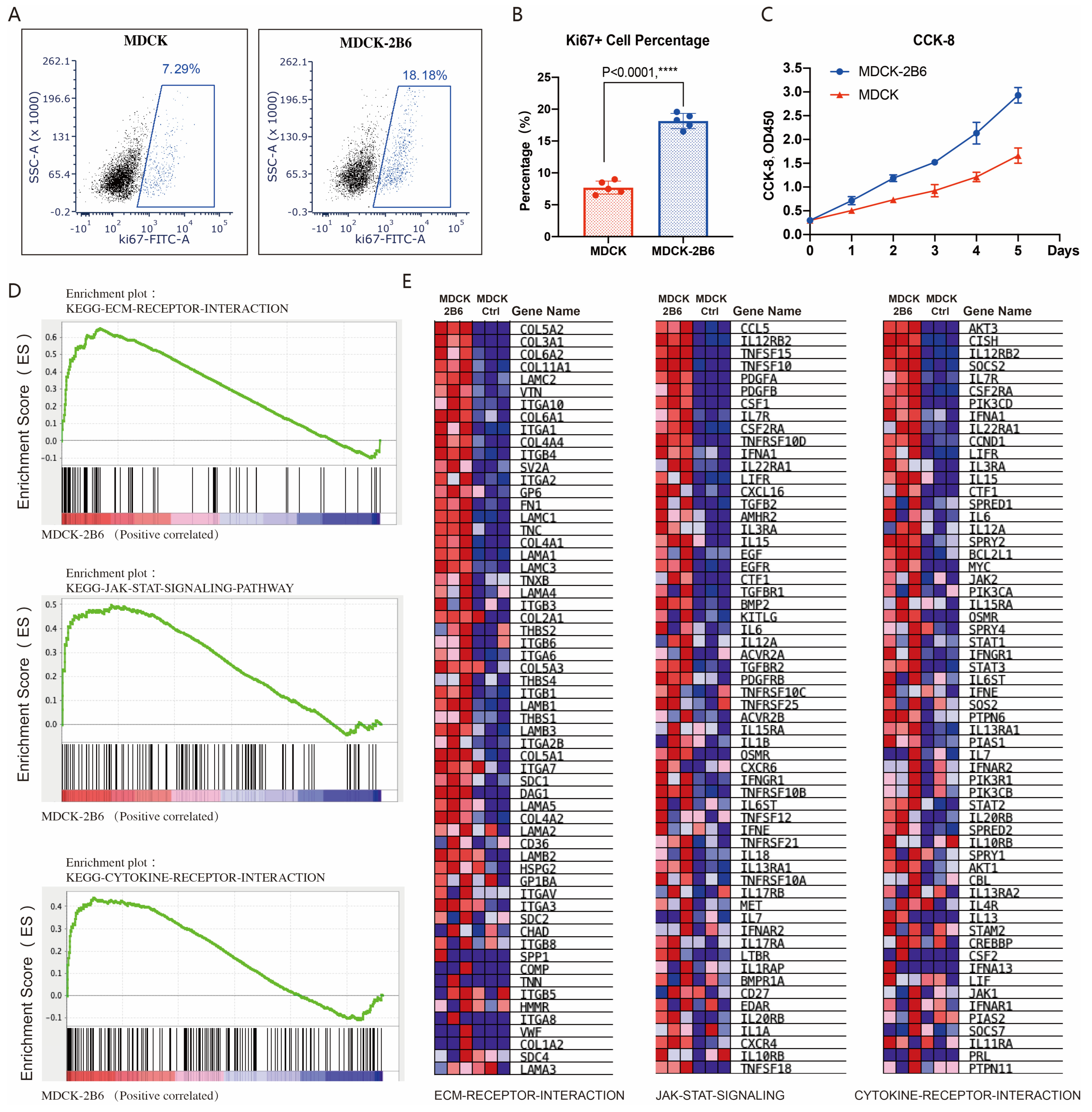
3.4. The Mechanism of MDCK-2B6 Sensitivity to Influenza Viruses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Javanian, M.; Barary, M.; Ghebrehewet, S.; Koppolu, V.; Vasigala, V.R.; Ebrahimpour, S. A brief review of influenza virus infection. J. Med. Virol. 2021, 93, 4638–4646. [Google Scholar] [CrossRef] [PubMed]
- McLean, H.Q.; Belongia, E.A. Influenza Vaccine Effectiveness: New Insights and Challenges. Cold Spring Harb. Perspect. Med. 2021, 11, a038315. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, E.; Wood, J.G.; Chadwick, C.; Newall, A.T.; Torvaldsen, S.; Moen, A.; Torelli, G. Global production capacity of seasonal and pandemic influenza vaccines in 2019. Vaccine 2021, 39, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Sambala, E.Z.; Ngcobo, N.; Machingaidze, S.; Wiyeh, A.B.; Mahasha, P.W.; Jaca, A.; Cooper, S.; Wiysonge, C.S. A global review of seasonal influenza vaccine introduction: Analysis of the WHO/UNICEF Joint Reporting Form. Expert Rev. Vaccines 2019, 18, 859–865. [Google Scholar] [CrossRef]
- Trombetta, C.M.; Marchi, S.; Manini, I.; Lazzeri, G.; Montomoli, E. Challenges in the development of egg-independent vaccines for influenza. Expert Rev. Vaccines 2019, 18, 737–750. [Google Scholar] [CrossRef]
- Gresset-Bourgeois, V.; Leventhal, P.S.; Pepin, S.; Hollingsworth, R.; Kazek-Duret, M.-P.; De Bruijn, I.; Samson, S.I. Quadrivalent inactivated influenza vaccine (VaxigripTetra™). Expert Rev. Vaccines 2018, 17, 1–11. [Google Scholar] [CrossRef]
- Manini, I.; Domnich, A.; Amicizia, D.; Rossi, S.; Pozzi, T.; Gasparini, R.; Panatto, D.; Montomoli, E. Flucelvax (Optaflu) for seasonal in-fluenza. Expert Rev. Vaccines 2015, 14, 789–804. [Google Scholar] [CrossRef]
- Kim, Y.; Hong, K.; Kim, H.; Nam, J. Influenza vaccines: Past, present, and future. Rev. Med. Virol. 2022, 32, e2243. [Google Scholar] [CrossRef]
- Bandell, A.R.; Simoes, E.A. Live attenuated influenza vaccine tetravalent: A clinical review. Expert Rev. Vaccines 2015, 14, 963–973. [Google Scholar] [CrossRef]
- Yang, L.P.H. Recombinant trivalent influenza vaccine (flublok®): A review of its use in the prevention of seasonal influenza in adults. Drugs 2013, 73, 1357–1366. [Google Scholar] [CrossRef]
- Chen, J.-R.; Liu, Y.-M.; Tseng, Y.-C.; Ma, C. Better influenza vaccines: An industry perspective. J. Biomed. Sci. 2020, 27, 33. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.-Z.; Jiao, P.-R.; Qi, W.-B.; Fan, H.-Y.; Liao, M. Development and strategies of cell-culture technology for influenza vaccine. Appl. Microbiol. Biotechnol. 2011, 89, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, M.; Mirzaei, H.; Salemi, M.; Momeni, F.; Mousavi, M.J.; Sadeghalvad, M.; Arjeini, Y.; Solaymani-Mohammadi, F.; Nahand, J.S.; Namdari, H.; et al. Influenza vaccine: Where are we and where do we go? Rev. Med. Virol. 2019, 29, e2014. [Google Scholar] [CrossRef] [PubMed]
- Milián, E.; Kamen, A.A. Current and emerging cell culture manufacturing technologies for influenza vaccines. BioMed Res. Int. 2015, 2015, 504831. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Phan, T.; Hu, W.-S.; Liu, X.; Fan, L.; Tan, W.-S.; Zhao, L. Transcriptomic Characterization Reveals Attributes of High Influenza Virus Productivity in MDCK Cells. Viruses 2021, 13, 2200. [Google Scholar] [CrossRef] [PubMed]
- Oxford, J.S.; Gill, D. Vaccines from the Spanish Influenza as a firm foundation for new developments. Hum. Vaccines Immunother. 2020, 16, 2051–2055. [Google Scholar] [CrossRef] [PubMed]
- Rimmelzwaan, G.F.; Osterhaus, A.D. Influenza vaccines: New developments. Curr. Opin. Pharmacol. 2001, 1, 491–496. [Google Scholar] [CrossRef]
- Demicheli, V.; Jefferson, T.; Di Pietrantonj, C.; Ferroni, E.; Thorning, S.; E Thomas, R.; Rivetti, A. Vaccines for preventing influenza in the elderly. Cochrane Database Syst. Rev. 2018, 2, CD004876. [Google Scholar] [CrossRef]
- National Immunization Advisory Committee (NIAC) Technical Working Group (TWG); Influenza Vaccination TWG. Technical guidelines for seasonal influenza vaccination in China (2023–2024). Zhonghua Liu Xing Bing Xue Za Zhi 2023, 44, 1507–1530. (In Chinese) [Google Scholar] [CrossRef]
- Committee on Infectious Diseases. Recommendations for Prevention and Control of Influenza in Children, 2022–2023. Pediatrics 2022, 150, e2022059274. [Google Scholar] [CrossRef]
- Bruckhoff, B. Zur Herstellung von Influenza-Impfstoffen auf MDCK-Zellkulturen [The production of influenza vaccines from MDCK cell cultures]. Pharm. Unserer Zeit 2011, 40, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Inside the Worldwide Influenza Centre: Monitoring the constant threat of flu and other viruses. BMJ 2023, 381, 929, Erratum in BMJ 2023, 381, 964. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, T.-T.; Lai, C.-C.; Weng, T.-C.; Cyue, M.-H.; Tsai, S.-Y.; Tseng, Y.-F.; Sung, W.-C.; Lee, M.-S.; Hu, A.Y.-C. The stability and immunogenicity of inactivated MDCK cell-derived influenza H7N9 viruses. Vaccine 2019, 37, 7117–7122. [Google Scholar] [CrossRef] [PubMed]
- Tjärnhage, E.; Brown, D.; Bogen, B.; Andersen, T.K.; Grødeland, G. Trimeric, APC-Targeted Subunit Vaccines Protect Mice against Seasonal and Pandemic Influenza. J. Virol. 2023, 97, e0169422. [Google Scholar] [CrossRef] [PubMed]
- Terrier, O.; Si-Tahar, M.; Ducatez, M.; Chevalier, C.; Pizzorno, A.; Le Goffic, R.; Crépin, T.; Simon, G.; Naffakh, N. Influenza viruses and coronaviruses: Knowns, unknowns, and common research challenges. PLoS Pathog. 2021, 17, e1010106. [Google Scholar] [CrossRef] [PubMed]
- Yamayoshi, S.; Kawaoka, Y. Current and future influenza vaccines. Nat. Med. 2019, 25, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Buchy, P.; Badur, S. Who and when to vaccinate against influenza. Int. J. Infect. Dis. 2020, 93, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, A.; Soleimanjahi, H.; Jamali, A.; Mehrbod, P.; Gholami, S.; Kianmehr, Z.; Feizi, N.; Saleh, M.; Bahrami, F.; Mokhtari-Azad, T.; et al. Comparison between MDCK and MDCK-SIAT1 cell lines as preferred host for cell culture-based influenza vaccine production. Biotechnol. Lett. 2016, 38, 941–948. [Google Scholar] [CrossRef]
- Oh, D.Y.; Barr, I.G.; Mosse, J.A.; Laurie, K.L. MDCK-SIAT1 cells show improved isolation rates for recent human influenza viruses compared to conventional MDCK cells. J. Clin. Microbiol. 2008, 46, 2189–2194. [Google Scholar] [CrossRef]
- Wen, Z.; Wu, C.; Chen, W.; Zeng, X.; Shi, J.; Ge, J.; Chen, H.; Bu, Z. Establishment of MDCK Stable Cell Lines Expressing TMPRSS2 and MSPL and Their Applications in Propagating Influenza Vaccine Viruses in Absence of Exogenous Trypsin. Biotechnol. Res. Int. 2015, 2015, 402628. [Google Scholar] [CrossRef]
- Qiu, Z.; Guo, S.; Liu, G.; Pei, M.; Liao, Y.; Wang, J.; Zhang, J.; Yang, D.; Qiao, Z.; Li, Z.; et al. TGM2 inhibits the proliferation, migration and tumorigenesis of MDCK cells. PLoS ONE 2023, 18, e0285136. [Google Scholar] [CrossRef] [PubMed]
- Pérez Rubio, A.; Eiros, J.M. Cell culture-derived flu vaccine: Present and future. Hum. Vaccines Immunother. 2018, 14, 1874–1882. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Yu, S.; Xu, Y.; Zhao, Y.; Li, L.; Sun, J.; Wang, X.; Guo, Y.; Zhang, Y. The Screening and Mechanism of Influenza-Virus Sensitive MDCK Cell Lines for Influenza Vaccine Production. Diseases 2024, 12, 20. https://doi.org/10.3390/diseases12010020
Yang Z, Yu S, Xu Y, Zhao Y, Li L, Sun J, Wang X, Guo Y, Zhang Y. The Screening and Mechanism of Influenza-Virus Sensitive MDCK Cell Lines for Influenza Vaccine Production. Diseases. 2024; 12(1):20. https://doi.org/10.3390/diseases12010020
Chicago/Turabian StyleYang, Zhaona, Shouzhi Yu, Ying Xu, Yuxiu Zhao, Lili Li, Jingjie Sun, Xin Wang, Yancen Guo, and Yuntao Zhang. 2024. "The Screening and Mechanism of Influenza-Virus Sensitive MDCK Cell Lines for Influenza Vaccine Production" Diseases 12, no. 1: 20. https://doi.org/10.3390/diseases12010020
APA StyleYang, Z., Yu, S., Xu, Y., Zhao, Y., Li, L., Sun, J., Wang, X., Guo, Y., & Zhang, Y. (2024). The Screening and Mechanism of Influenza-Virus Sensitive MDCK Cell Lines for Influenza Vaccine Production. Diseases, 12(1), 20. https://doi.org/10.3390/diseases12010020