
Neuron Circuit Failure and Pattern Learning in Electronic Spiking Neural Networks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Simulation Methods
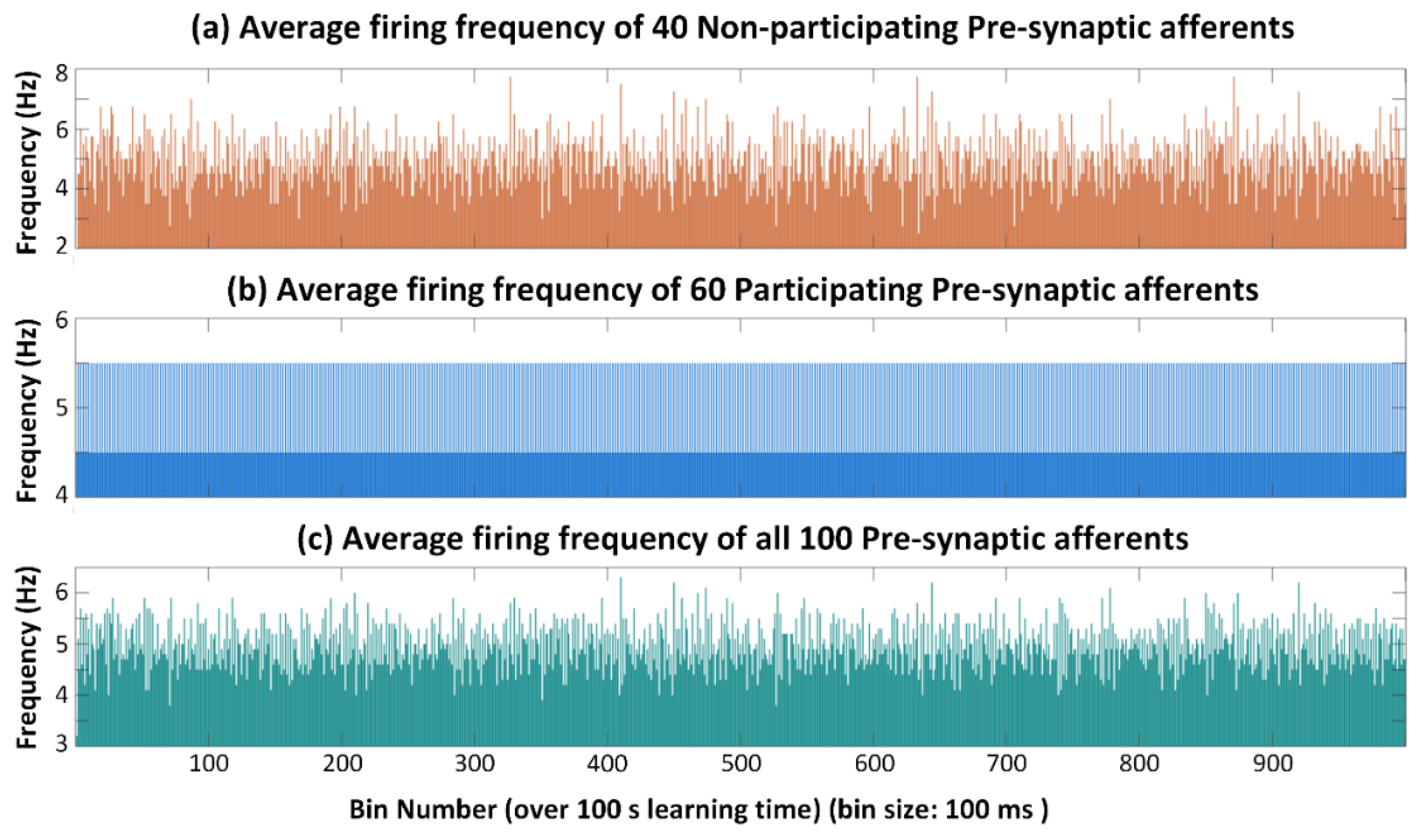
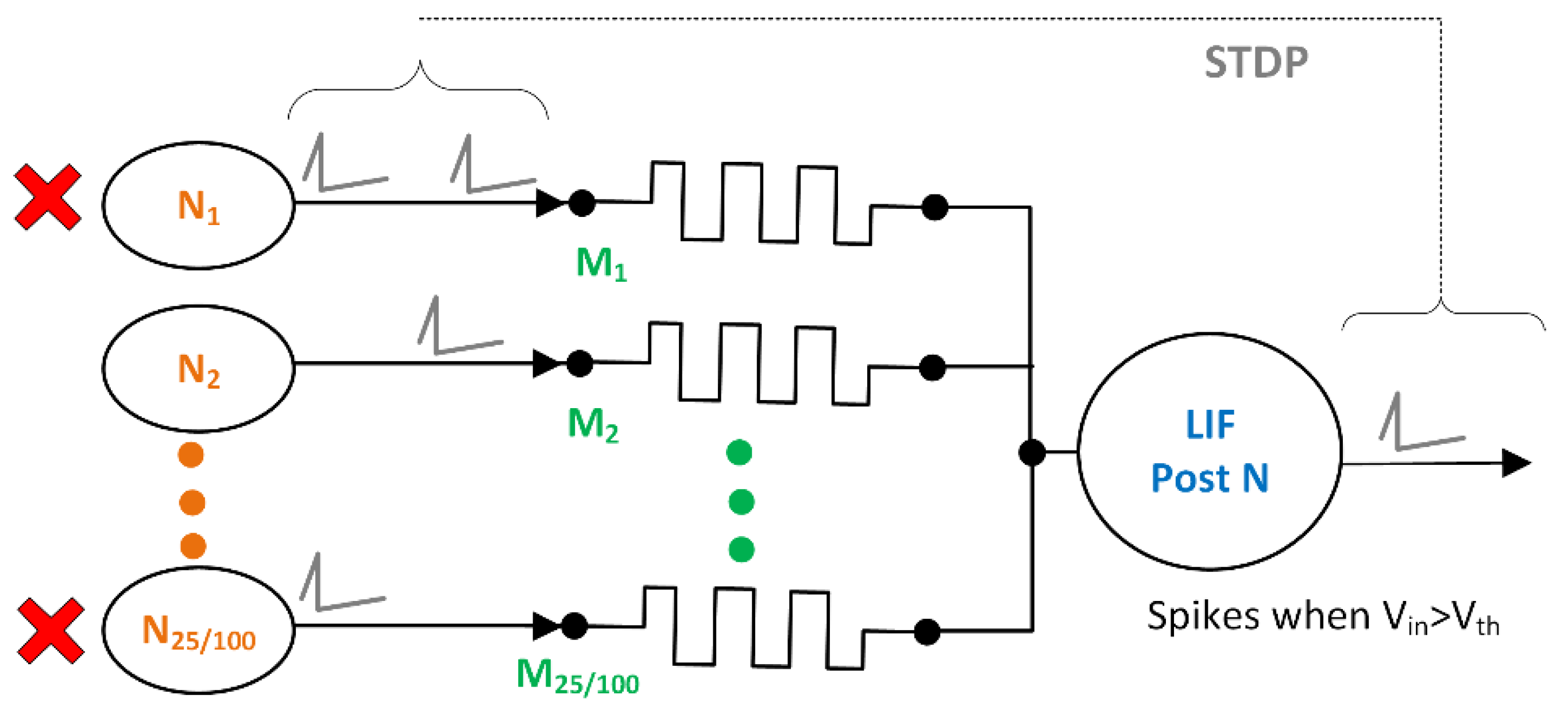
2.1. Neural Network Topology
2.2. Memristor Modeling
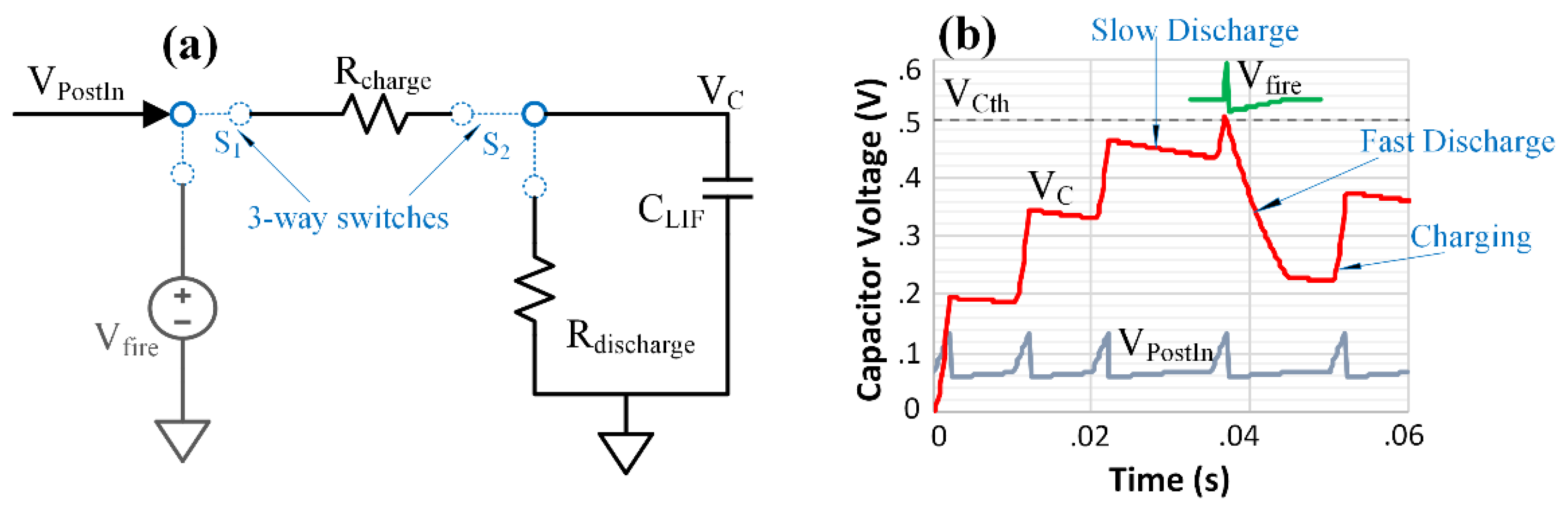
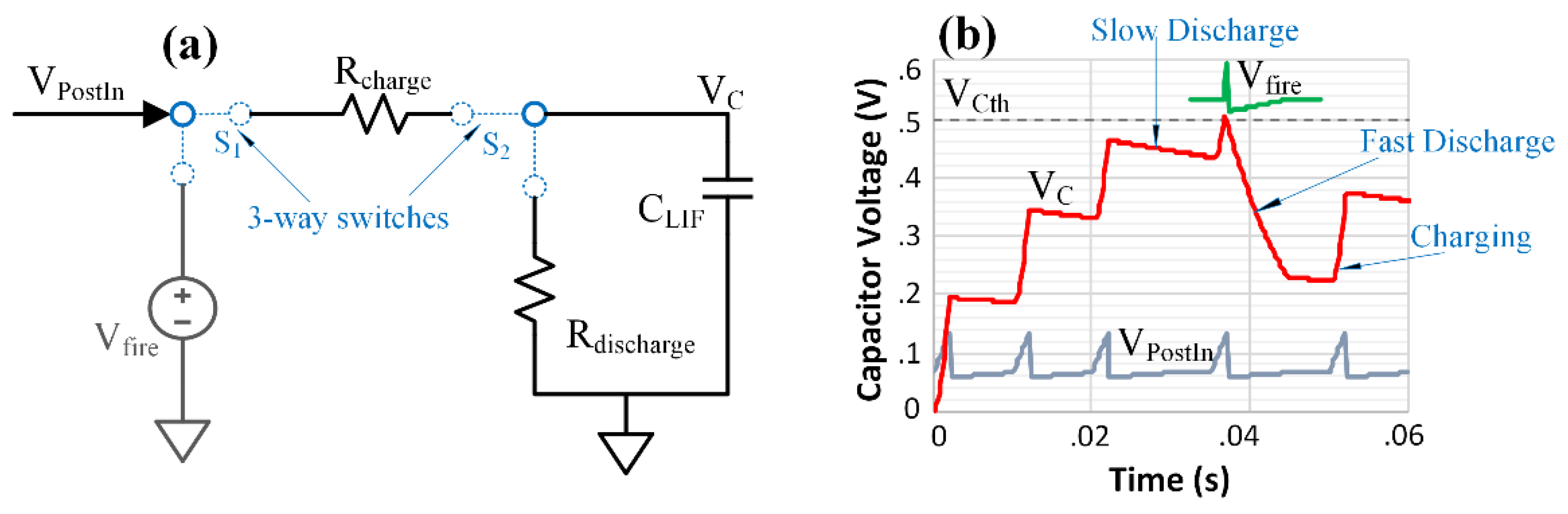
2.3. Post-Synaptic Neuron Design
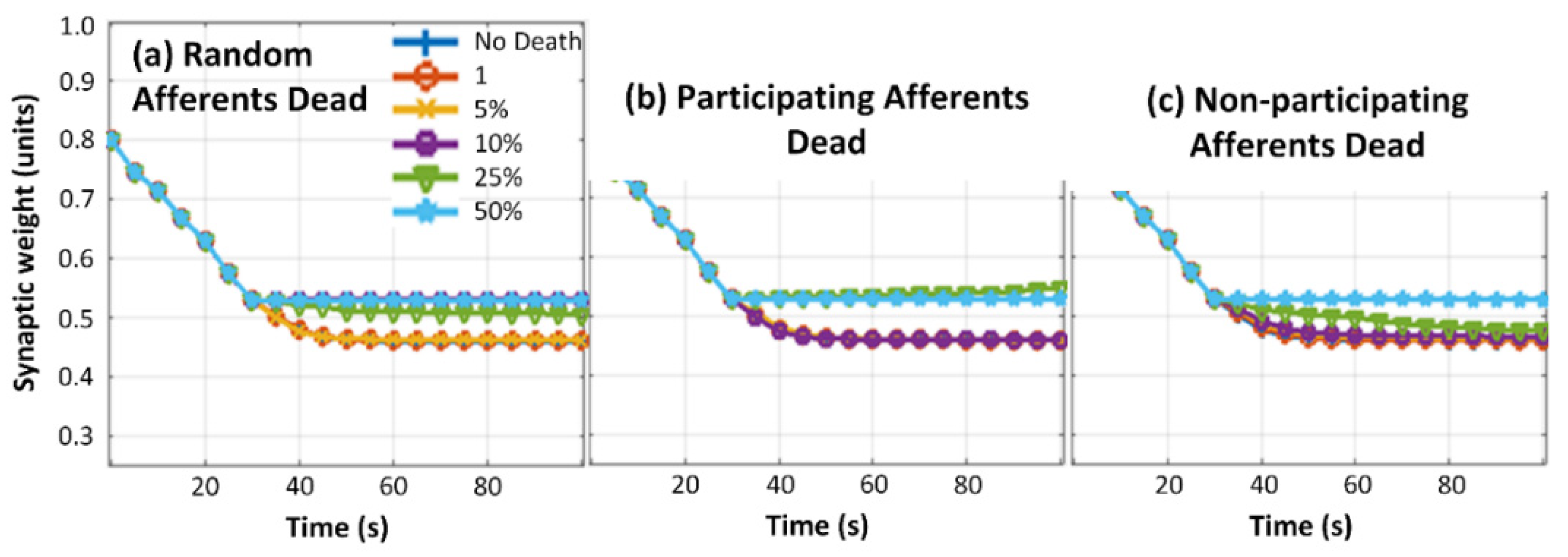
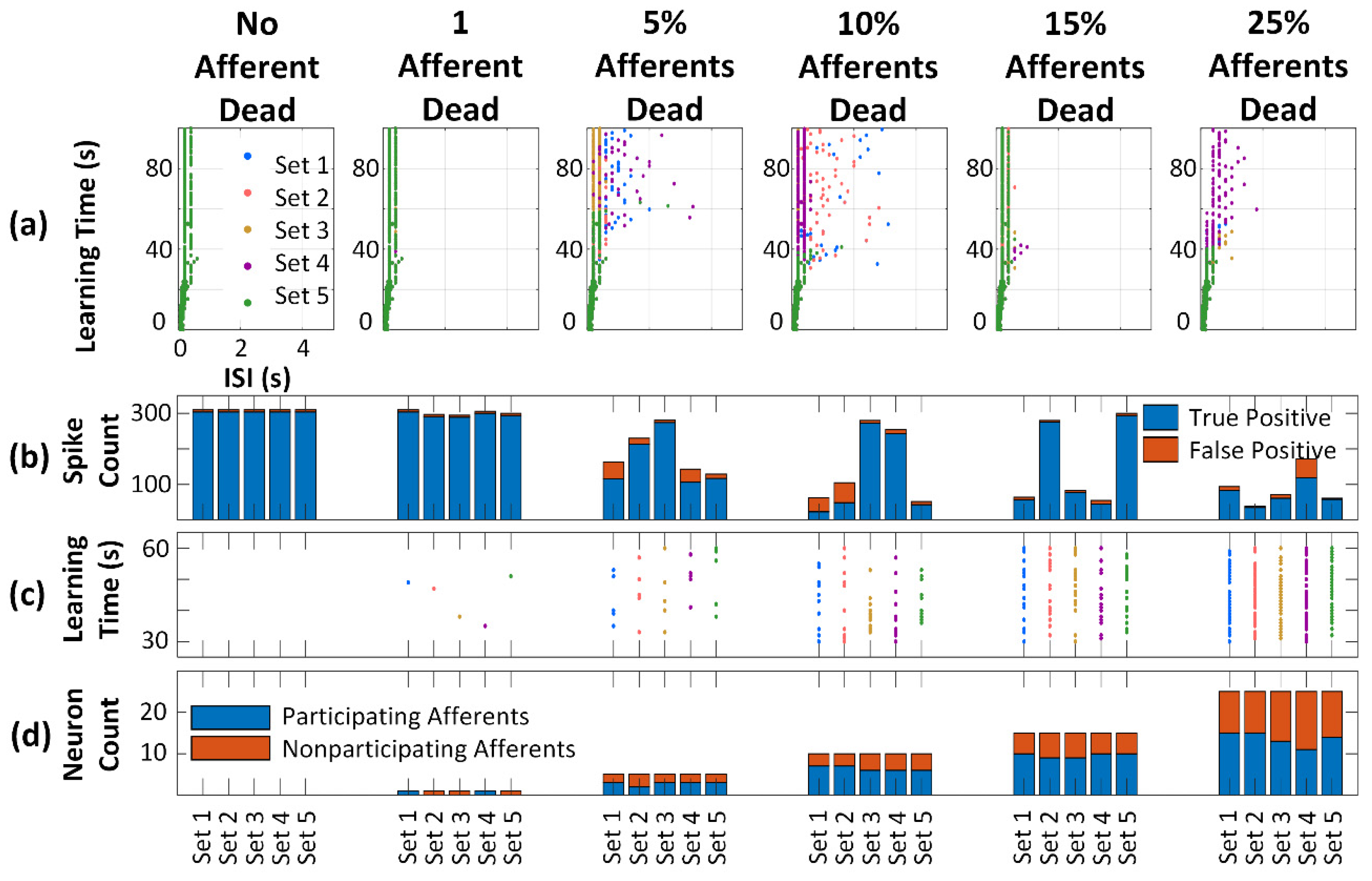
3. Results and Analysis
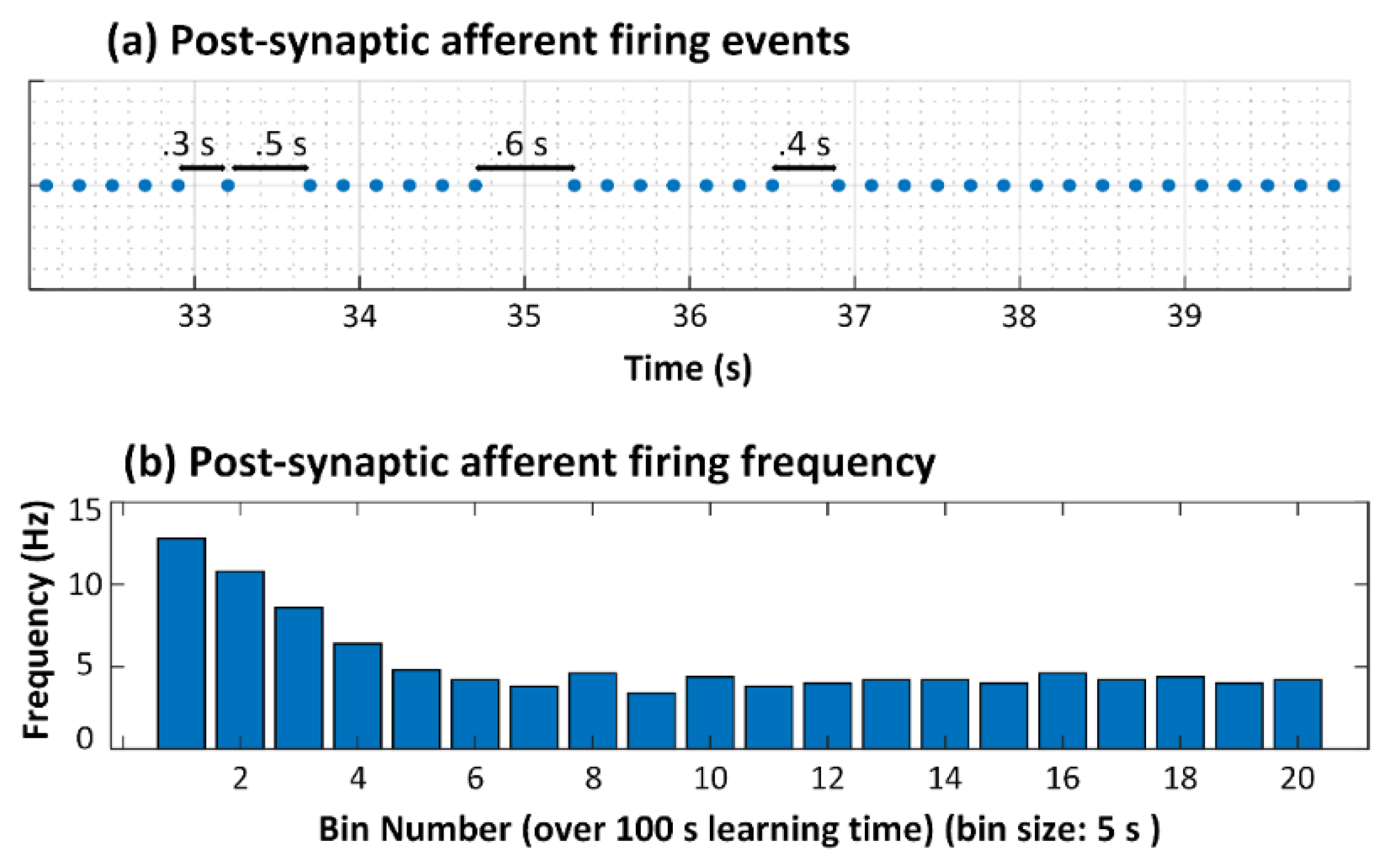
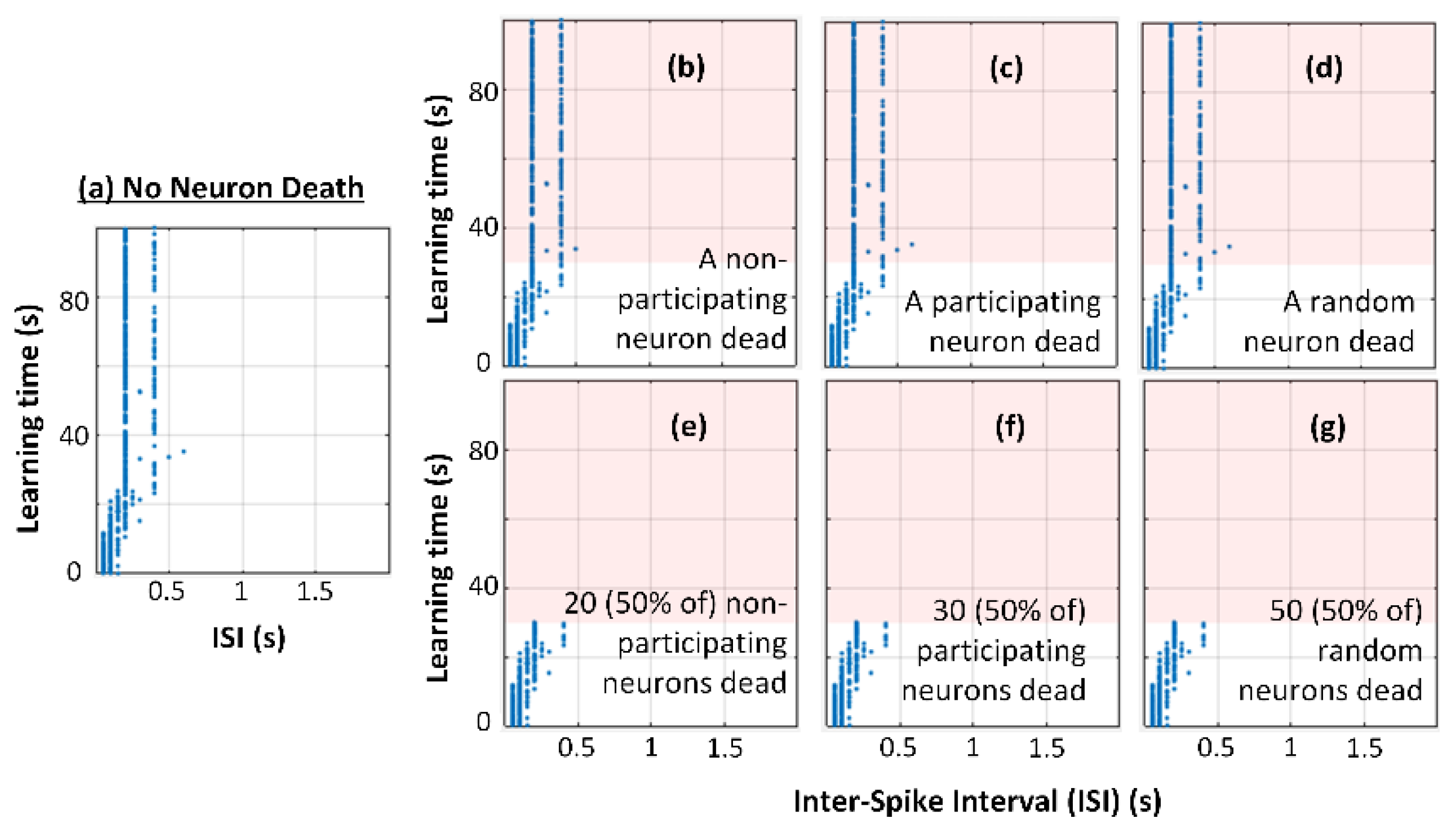
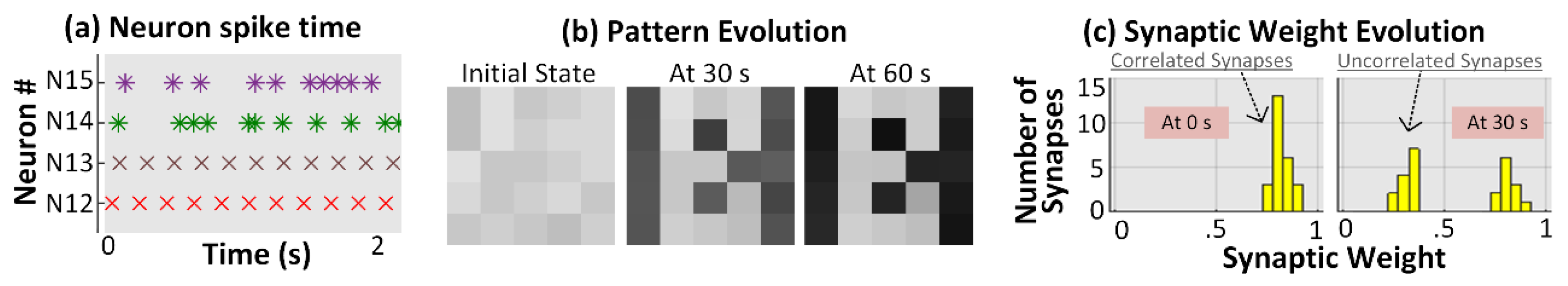
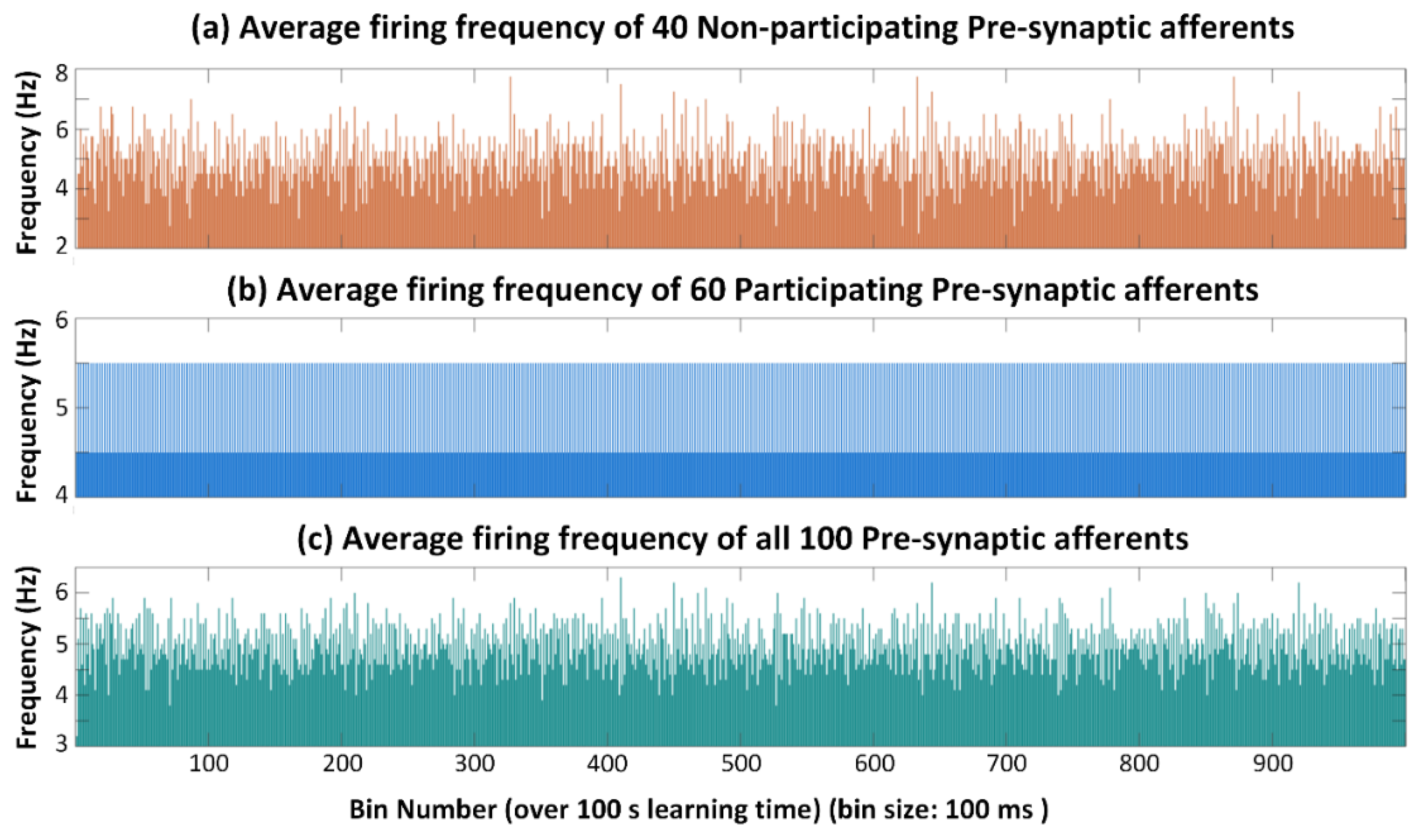
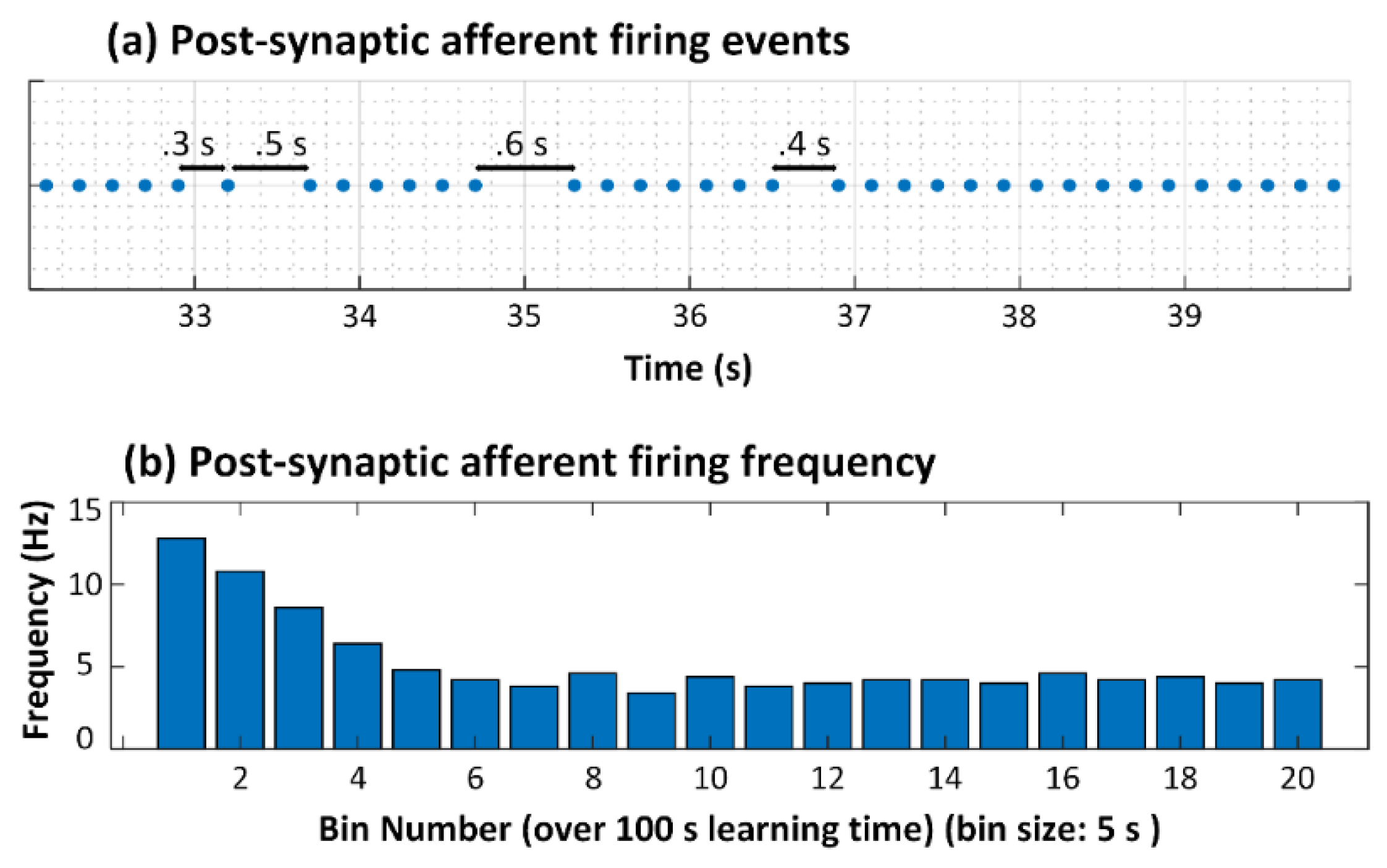
3.1. No Neuron Death (Control Case)
3.2. Neuron Death Setup
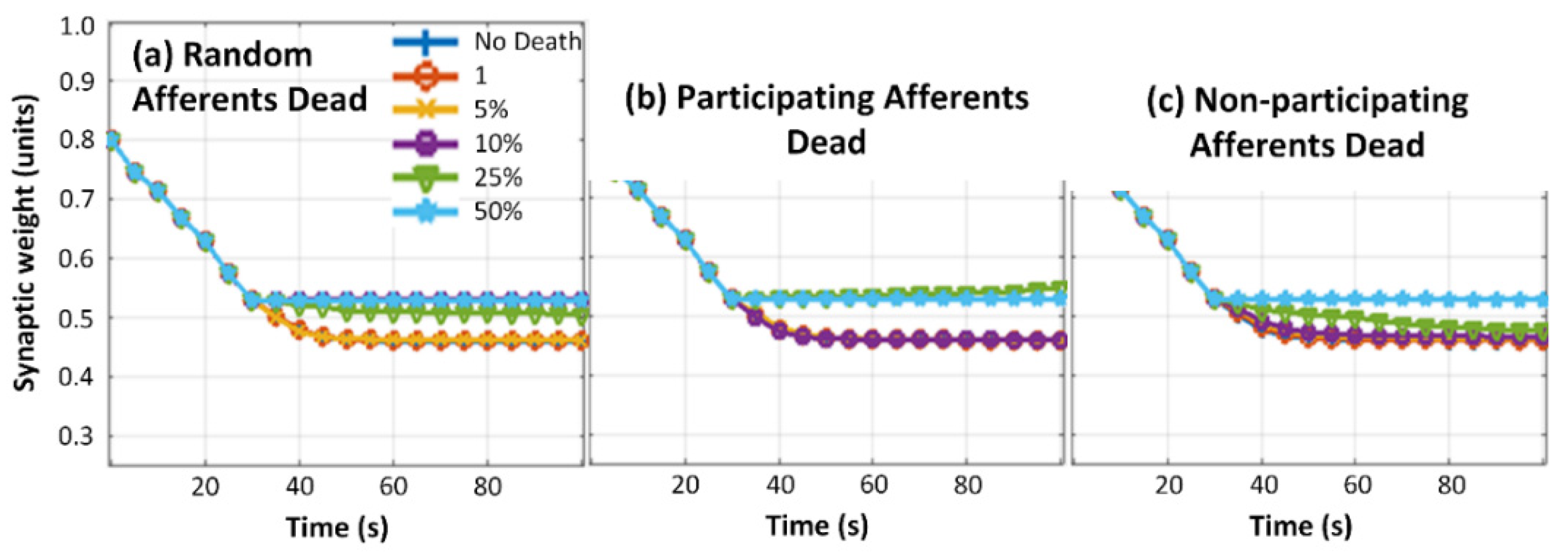
3.3. Neuron Death Simulations
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Morrison, J.H.; Hof, P.R. Life and death of neurons in the aging brain. Science 1997, 278, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Barrett, D.G.T.; Denève, S.; Machens, C.K. Optimal compensation for neuron loss. eLife 2016, 5, e12454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, V.; Benedetti, E.; Antonosante, A.; Catanesi, M.; Pitari, G.; Ippoliti, R.; Cimini, A.; d’Angelo, M. Neuronal cells rearrangement during aging and neurodegenerative disease: Metabolism, oxidative stress and organelles dynamic. Front. Mol. Neurosci. 2019, 12, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, R.D.; Masliah, E.; Salmon, D.P.; Butters, N.; DeTeresa, R.; Hill, R.; Hansen, L.A.; Katzman, R. Physical basis of cognitive alterations in alzheimer’s disease: Synapse loss is the major correlate of cognitive impairment. Ann. Neurol. 1991, 30, 572–580. [Google Scholar] [CrossRef]
- Li, N.; Daie, K.; Svoboda, K.; Druckmann, S. Robust neuronal dynamics in premotor cortex during motor planning. Nature 2016, 532, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Leary, M.C.; Saver, J.L. Annual Incidence of First Silent Stroke in the United States: A Preliminary Estimate. Cerebrovasc. Dis. 2003, 16, 280–285. [Google Scholar] [CrossRef]
- Büchel, J.; Zendrikov, D.; Solinas, S.; Indiveri, G.; Muir, D.R. Supervised training of spiking neural networks for robust deployment on mixed-signal neuromorphic processors. Sci. Rep. 2021, 11, 23376. [Google Scholar] [CrossRef]
- Alemi, A.; Denève, S.; Machens, C.K.; Slotine, J.J. Learning nonlinear dynamics in efficient, balanced spiking networks using local plasticity rules. In Proceedings of the 32nd AAAI Conference on Artificial Intelligence, AAAI 2018, New Orleans, LA, USA, 2–7 February 2018; pp. 588–595. [Google Scholar]
- Keys, A.S.; Adams, J.H.; Cressler, J.D.; Darty, R.C.; Johnson, M.A.; Patrick, M.C. High-performance, radiation-hardened electronics for space and lunar environments. AIP Conf. Proc. 2008, 969, 749–756. [Google Scholar] [CrossRef] [Green Version]
- Dodd, P.E.; Shaneyfelt, M.R.; Schwank, J.R.; Felix, J.A. Current and future challenges in radiation effects on CMOS electronics. IEEE Trans. Nucl. Sci. 2010, 57, 1747–1763. [Google Scholar] [CrossRef]
- Burghard, R.A.; Gwyn, C.W. Radiation failure modes in CMOS integrated circuits. IEEE Trans. Nucl. Sci. 1973, 20, 300–306. [Google Scholar] [CrossRef]
- Paccagnella, A.; Cester, A.; Cellere, G. Ionizing radiation effects on MOSFET thin and ultra-thin gate oxides. In Proceedings of the Technical Digest—International Electron Devices Meeting, IEDM, San Francisco, CA, USA, 13–15 December 2004; pp. 473–476. [Google Scholar]
- Tong, W.M.; Yang, J.J.; Kuekes, P.J.; Stewart, D.R.; Williams, R.S.; DeIonno, E.; King, E.E.; Witczak, S.C.; Looper, M.D.; Osborn, J.V. Radiation hardness of TiO2 memristive junctions. IEEE Trans. Nucl. Sci. 2010, 57, 1640–1643. [Google Scholar] [CrossRef]
- Deionno, E.; Looper, M.D.; Osborn, J.V.; Barnaby, H.J.; Tong, W.M. Radiation effects studies on thin film TiO2 memristor devices. IEEE Aerosp. Conf. Proc. 2013, 15, 1–8. [Google Scholar] [CrossRef]
- Marinella, M.J.; Dalton, S.M.; Mickel, P.R.; Dodd, P.E.D.; Shaneyfelt, M.R.; Bielejec, E.; Vizkelethy, G.; Kotula, P.G. Initial Assessment of the Effects of Radiation on the Electrical Characteristics of Memristive Memories. Nucl. Sci. IEEE Trans. 2012, 59, 2987–2994. [Google Scholar] [CrossRef]
- Barnaby, H.J.; Malley, S.; Land, M.; Charnicki, S.; Kathuria, A.; Wilkens, B.; Deionno, E.; Tong, W.M. Impact of alpha particles on the electrical characteristics of TiO2 memristors. IEEE Trans. Nucl. Sci. 2011, 58, 2838–2844. [Google Scholar] [CrossRef]
- Kheradpisheh, S.R.; Ganjtabesh, M.; Thorpe, S.J.; Masquelier, T. STDP-based spiking deep convolutional neural networks for object recognition. Neural Netw. 2018, 99, 56–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, S.G.; Ivans, R.C.; Cantley, K.D. Learning Behavior of Memristor-Based Neuromorphic Circuits in the Presence of Radiation. In Proceedings of the International Conference on Neuromorphic Systems—ICONS ’19 (under Rev.), Knoxville, TN, USA, 23–25 July 2019. [Google Scholar]
- Dahl, S.G.; Ivans, R.C.; Cantley, K.D. Radiation Effect on Learning Behavior in Memristor-Based Neuromorphic Circuit. In Proceedings of the 2019 IEEE 62nd International Midwest Symposium on Circuits and Systems (MWSCAS), IEEE, Dallas, TX, USA, 4–7 August 2019; Volume 2019, pp. 53–56. [Google Scholar]
- Gandharava Dahl, S.; Ivans, R.C.; Cantley, K.D. Effects of memristive synapse radiation interactions on learning in spiking neural networks. SN Appl. Sci. 2021, 3, 555. [Google Scholar] [CrossRef]
- Strukov, D.B.; Snider, G.S.; Stewart, D.R.; Williams, R.S. The missing memristor found. Nature 2008, 453, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, S.P.; Yang, C.; Kim, H.; Chua, L.O. Memristor bridge synapse-based neural network and its learning. IEEE Trans. Neural Netw. Learn. Syst. 2012, 23, 1426–1435. [Google Scholar] [CrossRef]
- Cantley, K.D.; Subramaniam, A.; Stiegler, H.J.; Chapman, R.A.; Vogel, E.M. Hebbian learning in spiking neural networks with nanocrystalline silicon TFTs and memristive synapses. IEEE Trans. Nanotechnol. 2011, 10, 1066–1073. [Google Scholar] [CrossRef]
- Cantley, K.D.; Subramaniam, A.; Stiegler, H.J.; Chapman, R.A.; Vogel, E.M. Neural learning circuits utilizing nano-crystalline silicon transistors and memristors. IEEE Trans. Neural Netw. Learn. Syst. 2012, 23, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Mcdonald, N.R.; Pino, R.E.; Member, S.; Wysocki, B.T.; Rozwood, P.J. Analysis of dynamic linear and non-linear memristor device models for emerging neuromorphic computing hardware design. In Proceedings of the 2010 International Joint Conference on Neural Networks (IJCNN), Barcelona, Spain, 18–23 July 2010. [Google Scholar]
- Prodromakis, T.; Boon Pin, P.; Papavassiliou, C.; Toumazou, C. A Versatile Memristor Model With Nonlinear Dopant Kinetics. IEEE Trans. Electron Devices 2011, 58, 3099–3105. [Google Scholar] [CrossRef]
- Yakopcic, C.; Taha, T.M.; Subramanyam, G.; Pino, R.E.; Rogers, S. A memristor device model. IEEE Electron Device Lett. 2011, 32, 1436–1438. [Google Scholar] [CrossRef]
- Joglekar, Y.N.; Wolf, S.J. The elusive memristor: Properties of basic electrical circuits. Eur. J. Phys. 2009, 30, 661–675. [Google Scholar] [CrossRef] [Green Version]
- Dahl, S.G.; Ivans, R.; Cantley, K.D. Modeling Memristor Radiation Interaction Events and the Effect on Neuromorphic Learning Circuits. In Proceedings of the International Conference on Neuromorphic Systems—ICONS ’18, Knoxville, TN, USA, 23–26 July 2018; pp. 1–8. [Google Scholar] [CrossRef]
- Dutta, S.; Kumar, V.; Shukla, A.; Mohapatra, N.R.; Ganguly, U. Leaky Integrate and Fire Neuron by Charge-Discharge Dynamics in Floating-Body MOSFET. Sci. Rep. 2017, 7, 8257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozenberg, M.J.; Schneegans, O.; Stoliar, P. An ultra-compact leaky-integrate-and-fire model for building spiking neural networks. Sci. Rep. 2019, 9, 11123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Saxena, V.; Zhu, K.; Balagopal, S. A CMOS Spiking Neuron for Brain-Inspired Neural Networks with Resistive Synapses and in Situ Learning. IEEE Trans. Circuits Syst. II Express Briefs 2015, 62, 1088–1092. [Google Scholar] [CrossRef] [Green Version]
- Cantley, K.D.; Ivans, R.C.; Subramaniam, A.; Vogel, E.M. Spatio-temporal pattern recognition in neural circuits with memory-transistor-driven memristive synapses. In Proceedings of the International Joint Conference on Neural Networks, Anchorage, AK, USA, 14–19 May 2017; Volume 2017, pp. 4633–4640. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, S.; Tuma, T.; Pantazi, A.; Eleftheriou, E. Learning spatio-temporal patterns in the presence of input noise using phase-change memristors. In Proceedings of the IEEE International Symposium on Circuits and Systems, Montreal, QC, Canada, 23–25 May 2016; Volume 2016, pp. 365–368. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandharava, S.; Ivans, R.C.; Etcheverry, B.R.; Cantley, K.D. Neuron Circuit Failure and Pattern Learning in Electronic Spiking Neural Networks. Electronics 2022, 11, 1392. https://doi.org/10.3390/electronics11091392
Gandharava S, Ivans RC, Etcheverry BR, Cantley KD. Neuron Circuit Failure and Pattern Learning in Electronic Spiking Neural Networks. Electronics. 2022; 11(9):1392. https://doi.org/10.3390/electronics11091392
Chicago/Turabian StyleGandharava, Sumedha, Robert C. Ivans, Benjamin R. Etcheverry, and Kurtis D. Cantley. 2022. "Neuron Circuit Failure and Pattern Learning in Electronic Spiking Neural Networks" Electronics 11, no. 9: 1392. https://doi.org/10.3390/electronics11091392
APA StyleGandharava, S., Ivans, R. C., Etcheverry, B. R., & Cantley, K. D. (2022). Neuron Circuit Failure and Pattern Learning in Electronic Spiking Neural Networks. Electronics, 11(9), 1392. https://doi.org/10.3390/electronics11091392