
The Effect of Autotrophic Cultivation of Platymonas subcordiformis in Waters from the Natural Aquatic Reservoir on Hydrogen Yield

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Materials
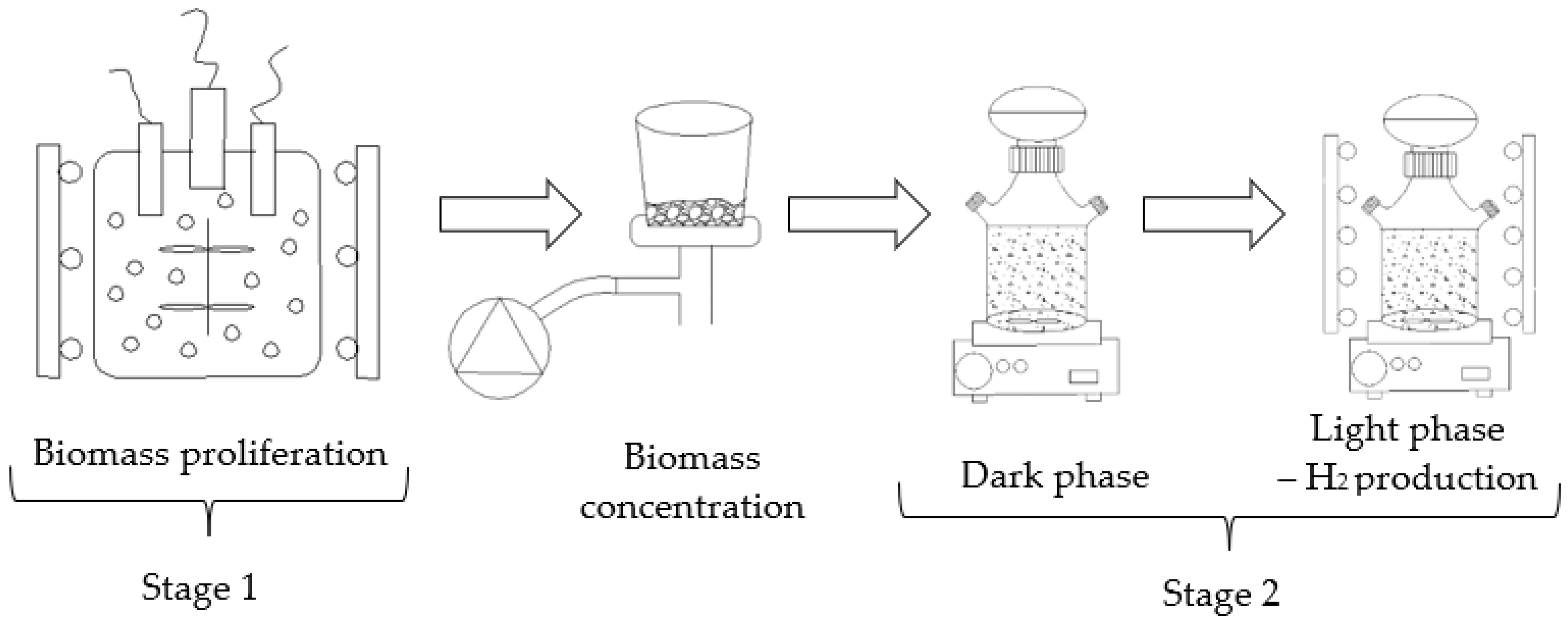
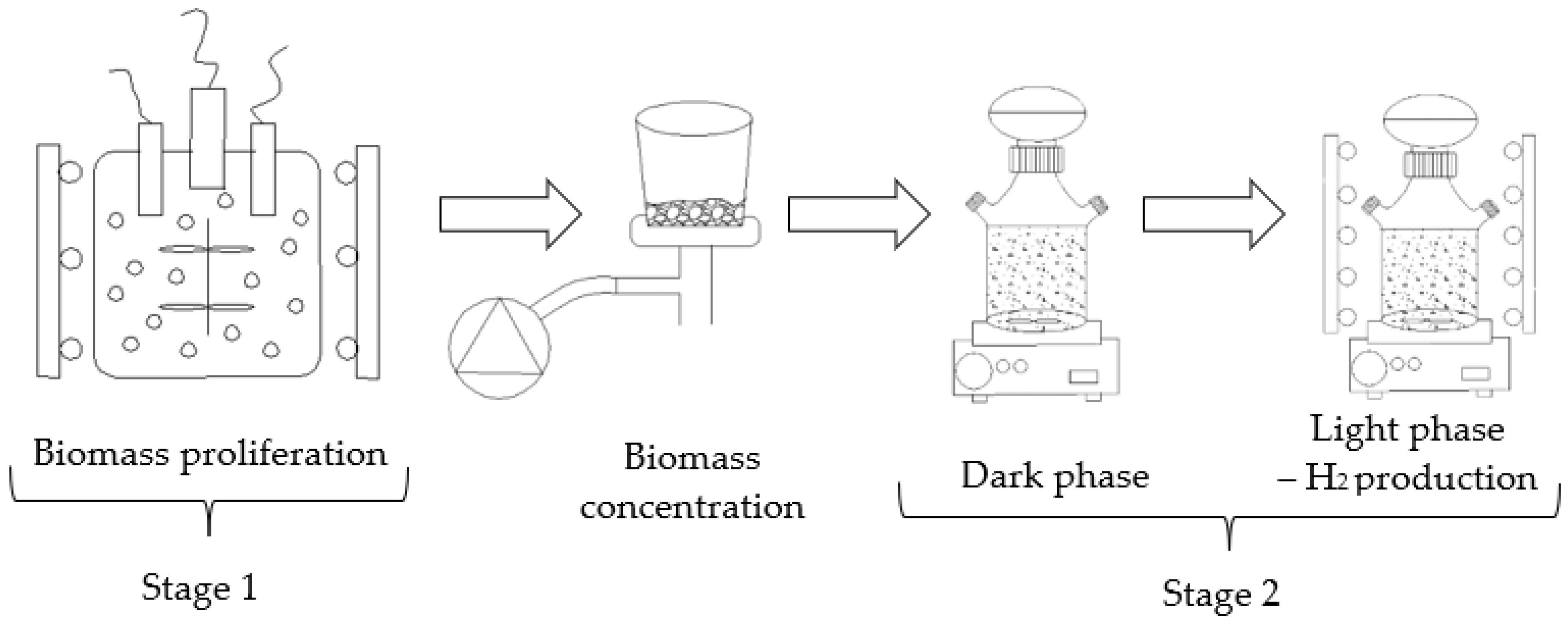
2.3. Experimental Station
2.4. Analytical and Statistical Methods
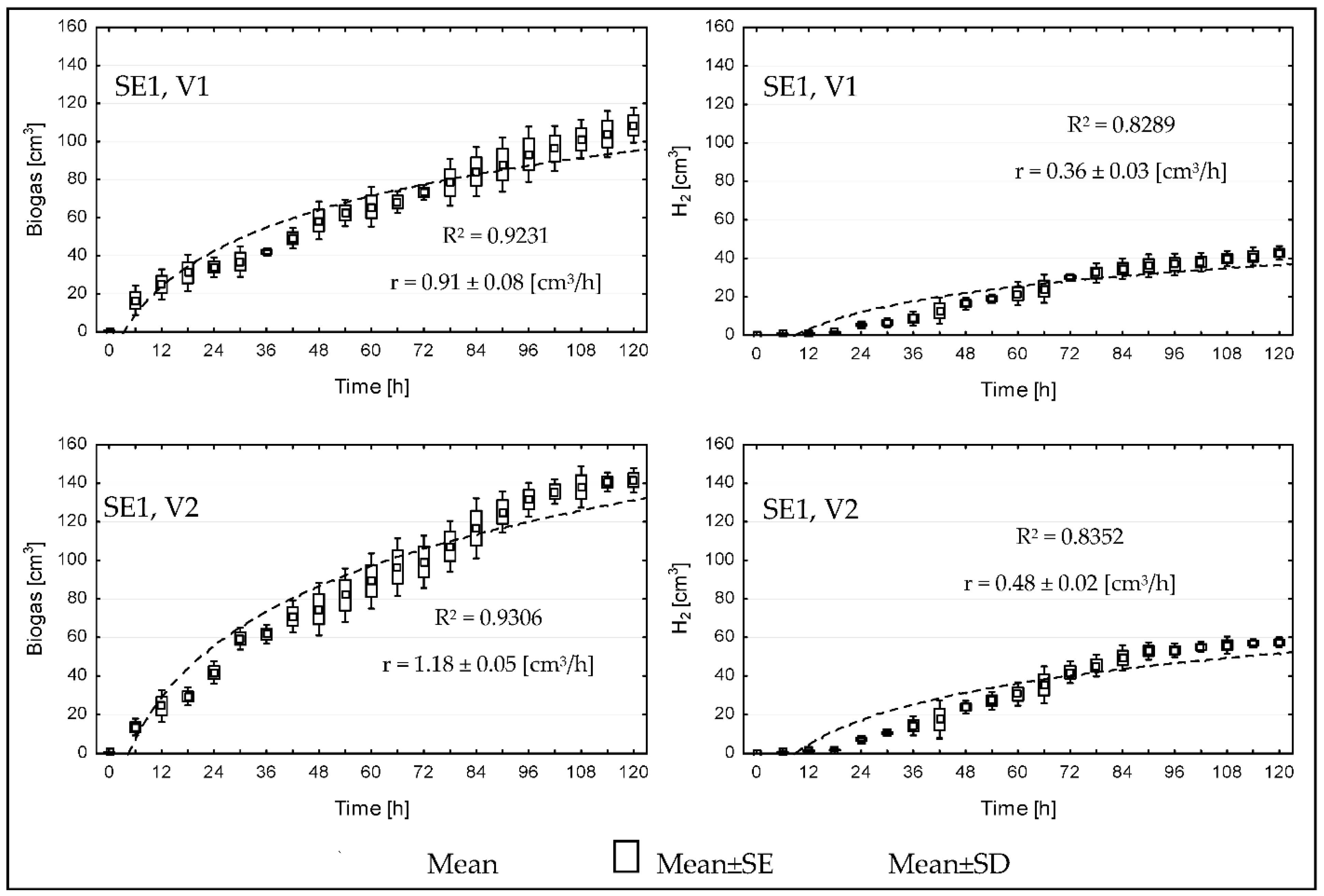
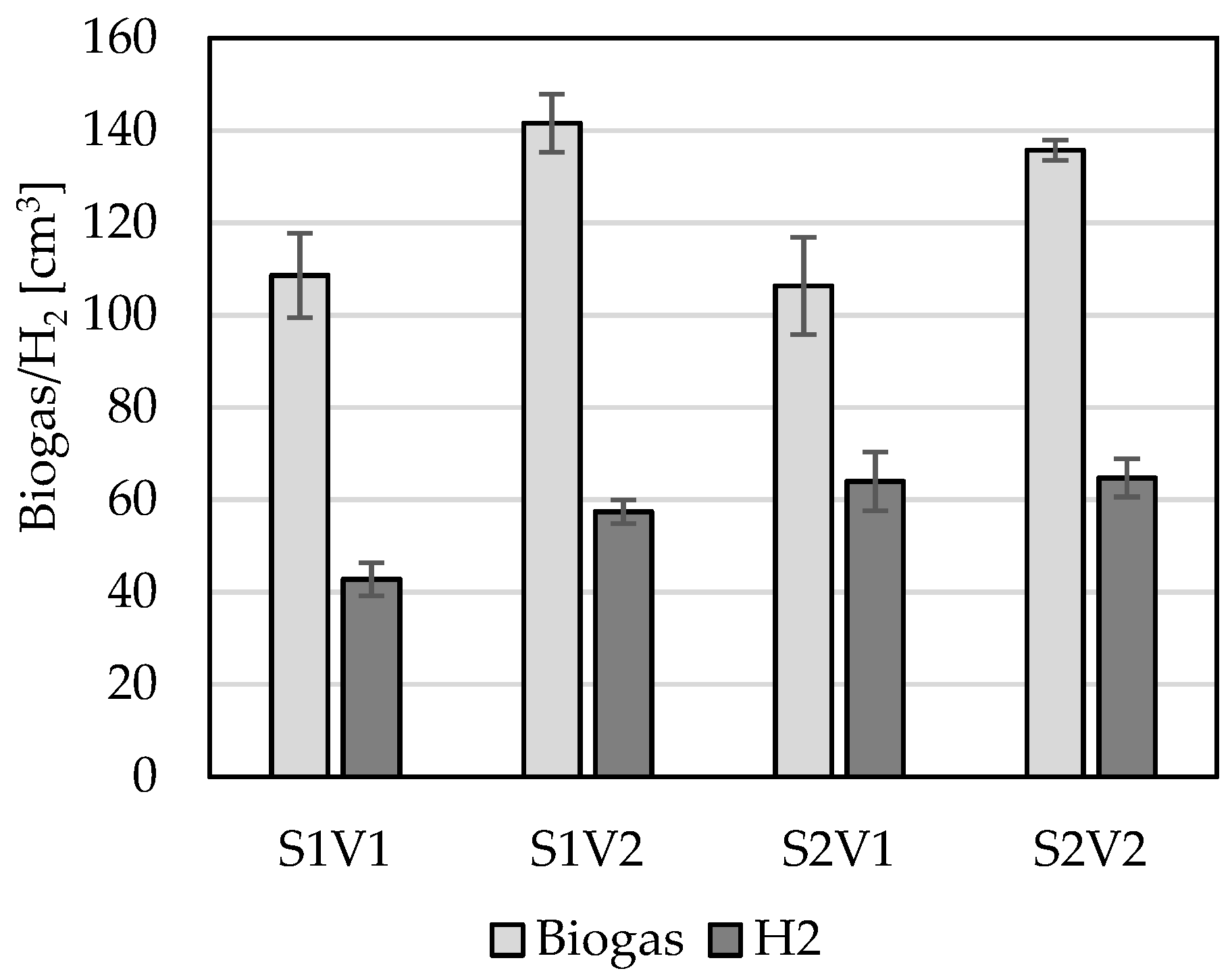
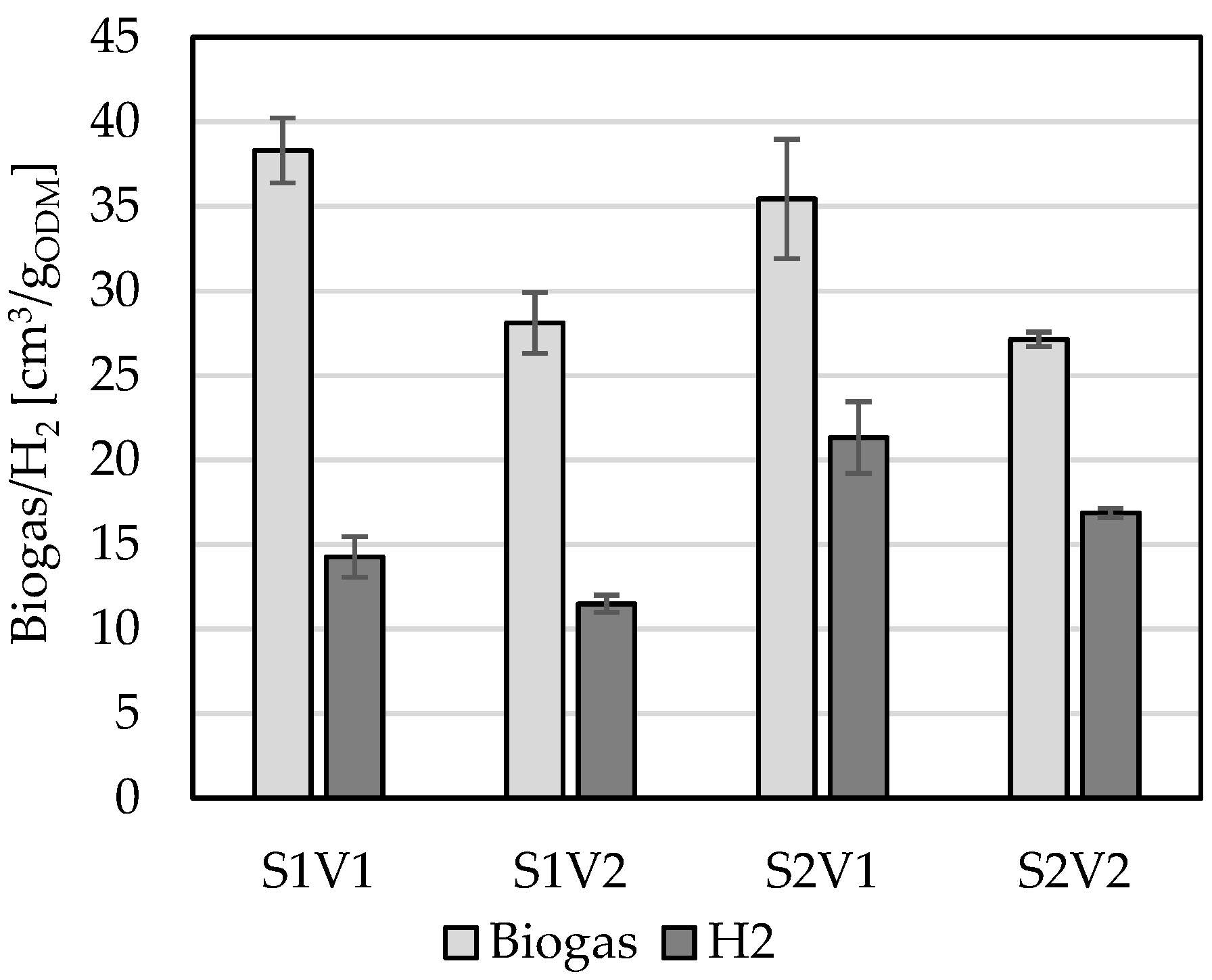
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dębowski, M.; Zieliński, M.; Świca, I.; Kazimierowicz, J. Algae Biomass as a Potential Source of Liquid Fuels. Phycology 2021, 1, 105–118. [Google Scholar] [CrossRef]
- Markowski, M.; Bialobrzewski, I.; Zielinski, M.; Dębowski, M.; Krzemieniewski, M. Optimizing low-temperature biogas production from biomass by anaerobic digestion. Renew. Energy 2014, 69, 219–225. [Google Scholar] [CrossRef]
- Manoharan, Y.; Hosseini, S.E.; Butler, B.; Alzhahrani, H.; Senior, B.T.F.; Ashuri, T.; Krohn, J. Hydrogen Fuel Cell Vehicles; Current Status and Future Prospect. Appl. Sci. 2019, 9, 2296. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Kumar, A.; Wong, W.C.; Chiu, M.-S.; Wang, X. Hydrogen value chain and fuel cells within hybrid renewable energy systems: Advanced operation and control strategies. Appl. Energy 2019, 233, 321–337. [Google Scholar] [CrossRef]
- Noussan, M.; Raimondi, P.P.; Scita, R.; Hafner, M. The Role of Green and Blue Hydrogen in the Energy Transition—A Technological and Geopolitical Perspective. Sustainability 2021, 13, 298. [Google Scholar] [CrossRef]
- Wu, Y.; He, F.; Zhou, J.; Wu, C.; Liu, F.; Tao, Y.; Xu, C. Optimal site selection for distributed wind power coupled hydrogen storage project using a geographical information system based multi-criteria decision-making approach: A case in China. J. Clean. Prod. 2021, 299, 126905. [Google Scholar] [CrossRef]
- Aziz, M.; Darmawan, A.; Juangsa, F.B. Hydrogen production from biomasses and wastes: A technological review. Int. J. Hydrogen Energy 2021, 46, 33756–33781. [Google Scholar] [CrossRef]
- Filippov, S.P.; Yaroslavtsev, A.B. Hydrogen energy: Development prospects and materials. Russ. Chem. Rev. 2021, 90, 627–643. [Google Scholar] [CrossRef]
- Lepszy, S. The cost-effectiveness of an energy storage system using hydrogen and underground gas storage. E3S Web Conf. 2019, 137, 01007. [Google Scholar] [CrossRef] [Green Version]
- Boretti, A. Production of hydrogen for export from wind and solar energy, natural gas, and coal in Australia. Int. J. Hydrogen Energy 2020, 45, 3899–3904. [Google Scholar] [CrossRef]
- Bolatkhan, K.; Kossalbayev, B.D.; Zayadan, B.K.; Tomo, T.; Veziroglu, T.N.; Allakhverdiev, S.I. Hydrogen production from phototrophic microorganisms: Reality and perspectives. Int. J. Hydrogen Energy 2019, 44, 5799–5811. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kisielewska, M.; Kazimierowicz, J.; Dudek, M.; Świca, I.; Rudnicka, A. The Cultivation of Lipid-Rich Microalgae Biomass as Anaerobic Digestate Valorization Technology—A Pilot-Scale Study. Processes 2020, 8, 517. [Google Scholar] [CrossRef]
- Wang, J.; Yin, Y. Fermentative hydrogen production using various biomass-based materials as feedstock. Renew. Sustain. Energy Rev. 2018, 92, 284–306. [Google Scholar] [CrossRef]
- El-Mohsnawy, E.; Abdelaal, A.S.; El-Sheekh, M. Chapter 14—Direct biohydrogen production from algae. In Handbook of Algal Biofuels: Aspects of Cultivation, Conversion, and Biorefinery; El-Sheekh, M., El-Fatah Abomohra, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 303–330. [Google Scholar] [CrossRef]
- Dębowski, M.; Dudek, M.; Zieliński, M.; Nowicka, A.; Kazimierowicz, J. Microalgal Hydrogen Production in Relation to Other Biomass-Based Technologies—A Review. Energies 2021, 14, 6025. [Google Scholar] [CrossRef]
- Grala, A.; Zieliński, M.; Dębowski, M.; Dudek, M. Effects of hydrothermal depolymerization and enzymatic hydrolysis of algae biomass on yield of methane fermentation process. Pol. J. Environ. Stud. 2012, 2, 361–366. [Google Scholar]
- Bhattacharya, M.; Goswami, S. Microalgae—A green multi-product biorefinery for future industrial prospects. Biocatal. Agric. Biotechnol. 2020, 25, 101580. [Google Scholar] [CrossRef]
- Park, H.; Jung, D.; Lee, J.; Kim, P.; Cho, Y.; Jung, I.; Kim, Z.-H.; Lim, S.-M.; Lee, C.-G. Improvement of biomass and fatty acid productivity in ocean cultivation of Tetraselmis sp. using hypersaline medium. Environ. Biol. Fishes 2018, 30, 2725–2735. [Google Scholar] [CrossRef]
- Calhoun, S.; Bell, T.A.S.; Dahlin, L.R.; Kunde, Y.; LaButti, K.; Louie, K.B.; Kuftin, A.; Treen, D.; Dilworth, D.; Mihaltcheva, S.; et al. A multi-omic characterization of temperature stress in a halotolerant Scenedesmus strain for algal biotechnology. Commun. Biol. 2021, 4, 333. [Google Scholar] [CrossRef]
- Pang, H.; Wang, Y.-N.; Chi, Z.-Y.; Xu, Y.-P.; Li, S.-Y.; Che, J.; Wang, J.-H. Enhanced aquaculture effluent polishing by once and repetitive nutrients deprived seawater Chlorella sp. Algal Res. 2021, 60, 102555. [Google Scholar] [CrossRef]
- Dębowski, M.; Zieliński, M.; Kazimierowicz, J.; Kujawska, N.; Talbierz, S. Microalgae Cultivation Technologies as an Opportunity for Bioenergetic System Development—Advantages and Limitations. Sustainability 2020, 12, 9980. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of Light Intensity and Quality on Growth Rate and Composition of Chlorella vulgaris. Plants 2020, 9, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deviram, G.; Mathimani, T.; Anto, S.; Ahamed, T.S.; Ananth, D.A.; Pugazhendhi, A. Applications of microalgal and cyanobacterial biomass on a way to safe, cleaner and a sustainable environment. J. Clean. Prod. 2020, 253, 119770. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, H.; Zhang, X.; Han, F.; Tu, W.; Yang, W. Microalgal hydrogen production. Small Methods 2020, 4, 1900514. [Google Scholar] [CrossRef]
- Limongi, A.R.; Viviano, E.; De Luca, M.; Radice, R.P.; Bianco, G.; Martelli, G. Biohydrogen from Microalgae: Production and Applications. Appl. Sci. 2021, 11, 1616. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Zhu, X.; Liao, Q.; Chang, J.-S.; Ho, S.-H. Biohydrogen production from microalgae for environmental sustainability. Chemosphere 2022, 291 Pt 1, 132717. [Google Scholar] [CrossRef]
- Show, K.Y.; Yan, Y.G.; Ling, M.; Ye, G.X.; Li, T.; Lee, D.J. Hydrogen production from algal biomass—Advances, challenges and prospects. Bioresour. Technol. 2018, 257, 290–300. [Google Scholar] [CrossRef]
- Sun, Y.; He, J.; Yang, G.; Sun, G.; Sage, V. A Review of the Enhancement of Bio-Hydrogen Generation by Chemicals Addition. Catalysts 2019, 9, 353. [Google Scholar] [CrossRef] [Green Version]
- Ghirardi, M.L.; Zhang, L.; Lee, J.W.; Flynn, T.; Seibert, M.; Greenbaum, E.; Melis, A. Microalgae: A green source of renewable H(2). Trends Biotechnol. 2000, 18, 506–511. [Google Scholar] [CrossRef]
- Fouchard, S.; Pruvost, J.; Degrenne, B.; Legrand, J. Investigation of H2 production using the green microalga Chlamydomonas reinhardtii in a fully controlled photobioreactor fitted with on-line gas analysis. Int. J. Hydrogen Energy 2008, 33, 3302–3310. [Google Scholar] [CrossRef] [Green Version]
- Wykoff, D.D.; Davies, J.; Melis, A.; Grossman, A.R. The regulation of photosynthetic electron transport during nutrient deprivation in Chlamydomonas reinhardtii. Plant Physiol. 1998, 117, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Melis, A.; Zhang, L.; Forestier, M.; Ghirardi, M.L.; Seibert, M. Sustained photobiological hydrogen gas production upon reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii. Plant Phys. 2000, 122, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gfeller, R.P.; Gibbs, M. Fermentative metabolism of Chlamydomonas reinhardtii: I. Analysis of fermentative products from starch in dark and light. Plant Physiol. 1984, 75, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghirardi, M.L.; Kosourov, S.; Tsygankov, A.; Rubin, A.; Seibert, M. Cyclic photobiological algal H2-production. In Proceedings of the 2002 U.S. DOE Hydrogen Program Review, NREL/CP-610-32405, Golden, Colorado, 6–10 May 2002; pp. 1–12. [Google Scholar]
- Sadvakasova, A.K.; Kossalbayev, B.D.; Zayadan, B.K.; Bolatkhan, K.; Alwasel, S.; Najafpour, M.M.; Tomo, T.; Allakhverdiev, S.I. Bioprocesses of hydrogen production by cyanobacteria cells and possible ways to increase their productivity. Renew. Sustain. Energy Rev. 2020, 133, 110054. [Google Scholar] [CrossRef]
- Lv, H.X.; Qiao, C.S.; Zhong, C.; Jia, S.R. Metabolic fingerprinting of Dunaliella salina cultured under sulfur deprivation conditions. J. Appl. Phycol. 2018, 30, 355–365. [Google Scholar] [CrossRef]
- Sekine, M.; Yoshida, A.; Akizuki, S.; Kishi, M.; Toda, T. Microalgae cultivation using undiluted anaerobic digestate by introducing aerobic nitrification–desulfurization treatment. Water Sci. Technol. 2020, 82, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Qiu, S.; Tremblay, D.; Viner, K.; Champagne, P.; Jessop, P.G. Centrate wastewater treatment with Chlorella vulgaris: Simultaneous enhancement of nutrient removal, biomass and lipid production. Chem. Eng. J. 2018, 342, 310–320. [Google Scholar] [CrossRef]
- Shokravi, Z.; Shokravi, H.; Chyuan, O.H.; Lau, W.J.; Koloor, S.S.R.; Petrů, M.; Ismail, A.F. Improving ‘Lipid Productivity’ in Microalgae by Bilateral Enhancement of Biomass and Lipid Contents: A Review. Sustainability 2020, 12, 9083. [Google Scholar] [CrossRef]
- Nagarajan, D.; Dong, C.D.; Chen, C.Y.; Lee, D.J.; Chang, J.S. Biohydrogen production from microalgae-Major bottlenecks and future research perspectives. Biotechnol. J. 2021, 16, e2000124. [Google Scholar] [CrossRef]
- Touloupakis, E.; Faraloni, C.; Silva Benavides, A.M.; Torzillo, G. Recent Achievements in Microalgal Photobiological Hydrogen Production. Energies 2021, 14, 7170. [Google Scholar] [CrossRef]
- Ji, C.F.; Legrand, J.; Pruvost, J.; Chen, Z.A.; Zhang, W. Characterization of hydrogen production by Platymonas Subcordiformis in torus photobioreactor. Int. J. Hydrogen Energy 2010, 35, 7200–7205. [Google Scholar] [CrossRef]
- Ban, S.; Lin, W.; Luo, Z.; Luo, J. Improving hydrogen production of Chlamydomonas reinhardtii by reducing chlorophyll content via atmospheric and room temperature plasma. Bioresour. Technol. 2019, 275, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Laurinavichene, T.V.; Tolstygina, I.V.; Galiulina, R.R.; Ghirardi, M.L.; Seibert, M.; Tsygankov, A.A. Dilution methods to deprive Chlamydomonas reinhardtii cultures of sulfur for subsequent hydrogen photoproduction. Int. J. Hydrogen Energy 2002, 27, 1245–1249. [Google Scholar] [CrossRef]
- Faraloni, C.; Ena, A.; Pintucci, C.; Torzillo, G. Enhanced hydrogen production by means of sulfur-deprived Chlamydomonas reinhardtii cultures grown in pretreated olive mill wastewater. Int. J. Hydrogen Energy 2011, 36, 5920–5931. [Google Scholar] [CrossRef]
- Skjanes, K.; Knutsen, G.; Källqvist, T.; Lindblad, P. H2 production from marine and freshwater species of green algae during sulfur deprivation and considerations for bioreactor design. Int. J. Hydrogen Energy 2008, 33, 511–521. [Google Scholar] [CrossRef]
- Jiménez-Llanos, M.R.-C.J.; Rendón-Castrillón, L.; Ocampo-López, C. Sustainable biohydrogen production by Chlorella sp. microalgae: A review. Int. J. Hydrogen Energy 2020, 45, 8310–8328. [Google Scholar] [CrossRef]
- Sirawattanamongkol, T.; Maswanna, T.; Maneeruttanarungroj, C. A newly isolated green alga Chlorella sp. KLSc59: Potential for biohydrogen production. J. Appl. Phycol. 2020, 32, 2927–2936. [Google Scholar] [CrossRef]
- Zhang, L.; He, M.; Liu, J. The enhancement mechanism of hydrogen photoproduction in Chlorella protothecoides under nitro-gen limitation and sulfur deprivation. Int. J. Hydrogen Energy 2014, 39, 8969–8976. [Google Scholar] [CrossRef]
- Song, W.; Rashid, N.; Choi, W.; Lee, K. Biohydrogen production by immobilized Chlorella sp. using cycles of oxygenic photo-synthesis and anaerobiosis. Bioresour. Technol. 2011, 102, 8676–8681. [Google Scholar] [CrossRef]
- Ji, C.F.; Yu, X.J.; Chen, Z.A.; Xue, S.; Legrand, J.; Zhang, W. Effects of nutrient deprivation on biochemical compositions and photo-hydrogen production of Tetraselmis subcordiformis. Int. J. Hydrogen Energy 2011, 36, 5817–5821. [Google Scholar] [CrossRef]
- Guo, Z.; Li, Y.; Guo, H.Y. Characterization of H2 photoproduction by marine green alga Tetraselmis subcordiformis integrated with an alkaline fuel cell. Biotechnol. Lett. 2016, 38, 435–440. [Google Scholar] [CrossRef]
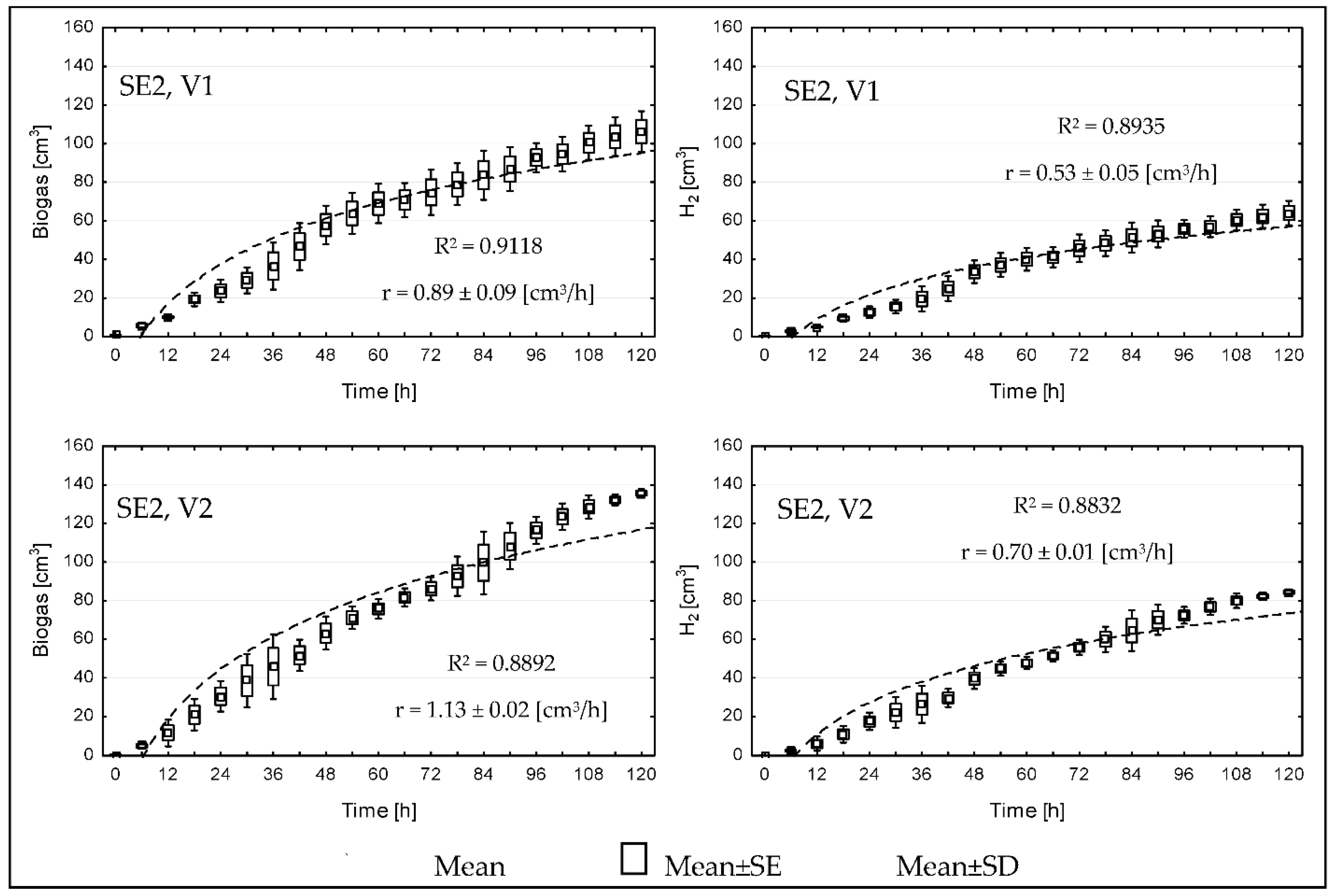

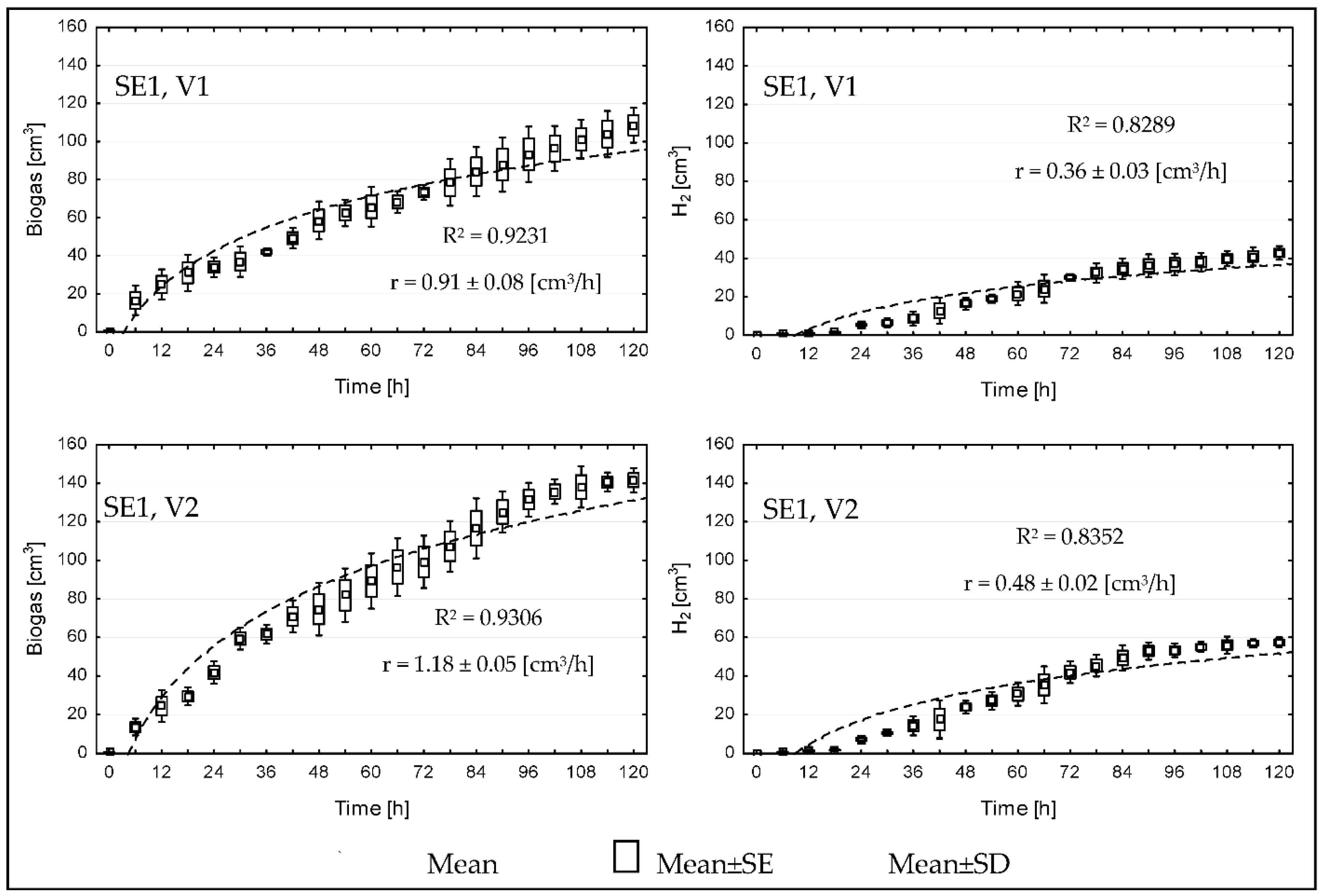
{kind=link}
{kind=link}
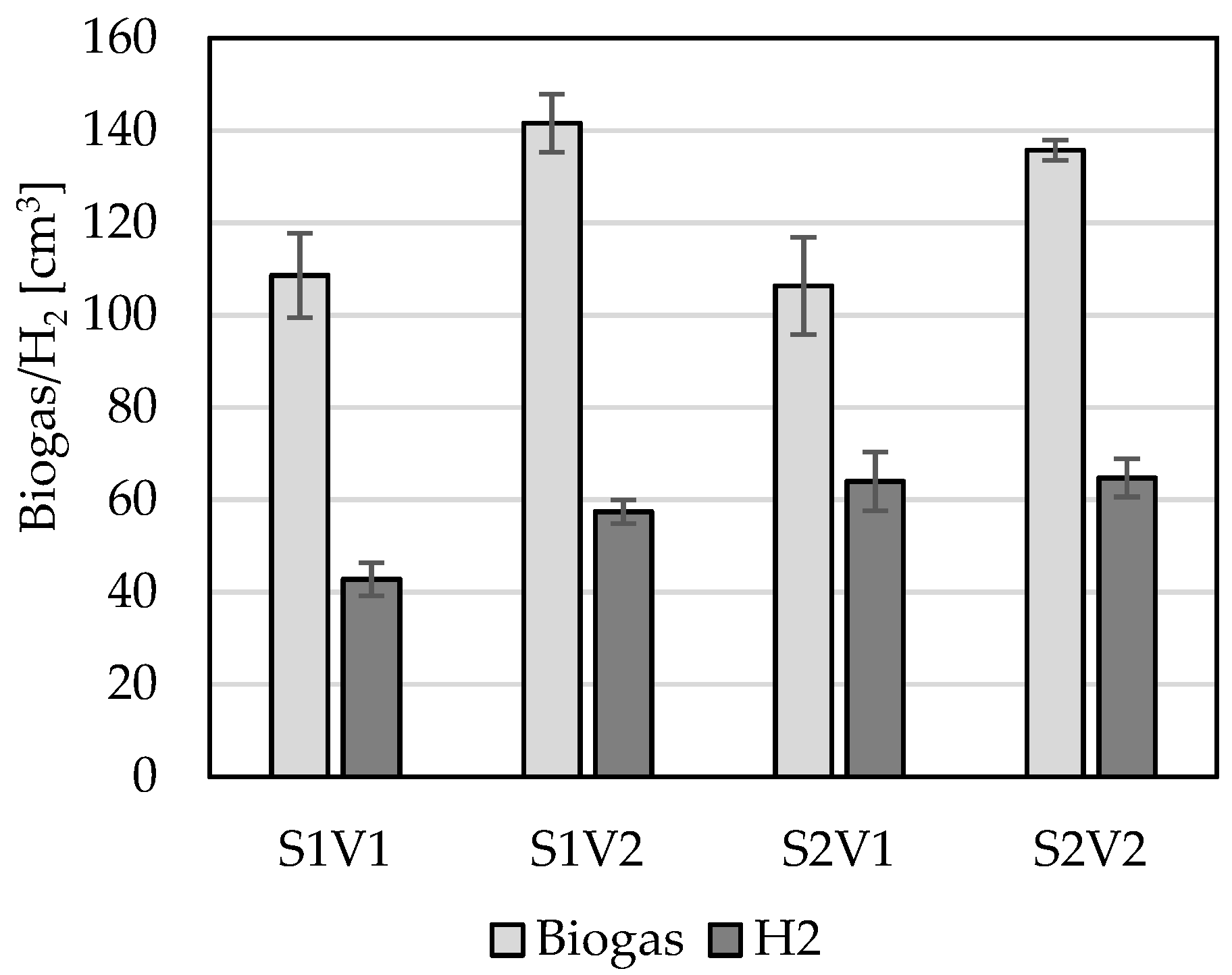{kind=link}
{kind=link}
{kind=link}
| ST1—cultivation of P. subcordiformis | SE1—culture medium based on deionized water and chemical reagents. | ST2—H2 production by P. subcordiformis | SE1 | V1—3.0 gODM/dm3 |
| V2—5.0 gODM/dm3 | ||||
| SE2—culture medium based on water from the Gdańsk Bay supplemented with chemical reagents. | SE2 | V1—3.0 gODM/dm3 | ||
| V2—5.0 gODM/dm3 |
| Series | Indicator | ||||||
|---|---|---|---|---|---|---|---|
| Final Biomass Concentration [mg ODM/dm3] | Biomass Growth Rater [mgODM/dm3·d] | Final Concentration of Chlorophyll a [μg/dm3] | Effectiveness of Ntot. Removal [%] | Effectiveness of Ptot. Removal [%] | Ntot. Consumption for Biomass Growth [mg Ntot/gODM] | Ptot. Consumption for Biomass Growth [mg Ptot/gODM] | |
| SE1 | 3203 ± 35 | 291.2 ± 3.2 | 3686 ± 320 | 98.1 ± 0.6 | 97.6 ± 1.1 | 7.0 ± 0.4 | 1.7 ± 0.1 |
| SE2 | 3493 ± 465 | 317.6 ± 42.3 | 3845 ± 696 | 98.4 ± 0.9 | 99.1 ± 0.4 | 6.7 ± 0.6 | 1.5 ± 0.1 |
| Series | Variant | H2 [%] | CO2 [%] | O2 [%] |
|---|---|---|---|---|
| SE1 | V1 | 41.0 ± 1.4 | 55.3 ± 1.8 | 3.7 ± 0.2 |
| V2 | 40.2 ± 1.4 | 57.3 ± 1.7 | 2.5 ± 0.1 | |
| SE2 | V1 | 59.9 ± 1.6 | 36.5 ± 1.3 | 3.6 ± 0.2 |
| V2 | 63.2 ± 1.4 | 34.6 ± 1.2 | 2.2 ± 0.1 |
| Variant | Total concentration of biogas produced | Variant | Total concentration of biogas produced per gODM | ||||||
| SE1V1 | SE1V2 | SE2V1 | SE2V2 | SE1V1 | SE1V2 | SE2V1 | SE2V2 | ||
| SE1V1 | 0.013492 | 1.000000 | 0.070077 | SE1V1 | 0.050933 | 1.000000 | 0.015422 | ||
| SE1V2 | 0.013492 | 0.006948 | 0.999733 | SE1V2 | 0.050933 | 0.103722 | 0.999989 | ||
| SE2V1 | 1.000000 | 0.006948 | 0.037985 | SE2V1 | 1.000000 | 0.103722 | 0.033422 | ||
| SE2V2 | 0.070077 | 0.999733 | 0.037985 | SE2V2 | 0.015422 | 0.999989 | 0.033422 | ||
| Variant | Total concentration of hydrogen produced | Variant | Total concentration of hydrogen produced per gODM | ||||||
| SE1V1 | SE1V2 | SE2V1 | SE2V2 | SE1V1 | SE1V2 | SE2V1 | SE2V2 | ||
| SE1V1 | 0.611181 | 0.000816 | 0.690782 | SE1V1 | 0.153253 | 0.008029 | 0.000143 | ||
| SE1V2 | 0.611181 | 0.000145 | 0.015779 | SE1V2 | 0.153253 | 0.960106 | 0.000548 | ||
| SE2V1 | 0.000816 | 0.000145 | 0.074735 | SE2V1 | 0.008029 | 0.960106 | 0.011987 | ||
| SE2V2 | 0.690782 | 0.015779 | 0.074735 | SE2V2 | 0.000143 | 0.000548 | 0.011987 | ||
| Variant | % content of hydrogen in biogas | Values in italics denote differences significant at p ≤ 0.05 | |||||||
| SE1V1 | SE1V2 | SE2V1 | SE2V2 | ||||||
| SE1V1 | 0.999974 | 0.000143 | 0.000143 | ||||||
| SE1V2 | 0.999974 | 0.000143 | 0.000143 | ||||||
| SE2V1 | 0.000143 | 0.000143 | 0.523867 | ||||||
| SE2V2 | 0.000143 | 0.000143 | 0.523867 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dudek, M.; Dębowski, M.; Nowicka, A.; Kazimierowicz, J.; Zieliński, M. The Effect of Autotrophic Cultivation of Platymonas subcordiformis in Waters from the Natural Aquatic Reservoir on Hydrogen Yield. Resources 2022, 11, 31. https://doi.org/10.3390/resources11030031
Dudek M, Dębowski M, Nowicka A, Kazimierowicz J, Zieliński M. The Effect of Autotrophic Cultivation of Platymonas subcordiformis in Waters from the Natural Aquatic Reservoir on Hydrogen Yield. Resources. 2022; 11(3):31. https://doi.org/10.3390/resources11030031
Chicago/Turabian StyleDudek, Magda, Marcin Dębowski, Anna Nowicka, Joanna Kazimierowicz, and Marcin Zieliński. 2022. "The Effect of Autotrophic Cultivation of Platymonas subcordiformis in Waters from the Natural Aquatic Reservoir on Hydrogen Yield" Resources 11, no. 3: 31. https://doi.org/10.3390/resources11030031
APA StyleDudek, M., Dębowski, M., Nowicka, A., Kazimierowicz, J., & Zieliński, M. (2022). The Effect of Autotrophic Cultivation of Platymonas subcordiformis in Waters from the Natural Aquatic Reservoir on Hydrogen Yield. Resources, 11(3), 31. https://doi.org/10.3390/resources11030031