Red-Light Irradiation of Horse Spermatozoa Increases Mitochondrial Activity and Motility through Changes in the Motile Sperm Subpopulation Structure

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Suppliers
2.2. Animals and Ejaculates
2.3. Experimental Design
2.4. Analysis of Sperm Motility
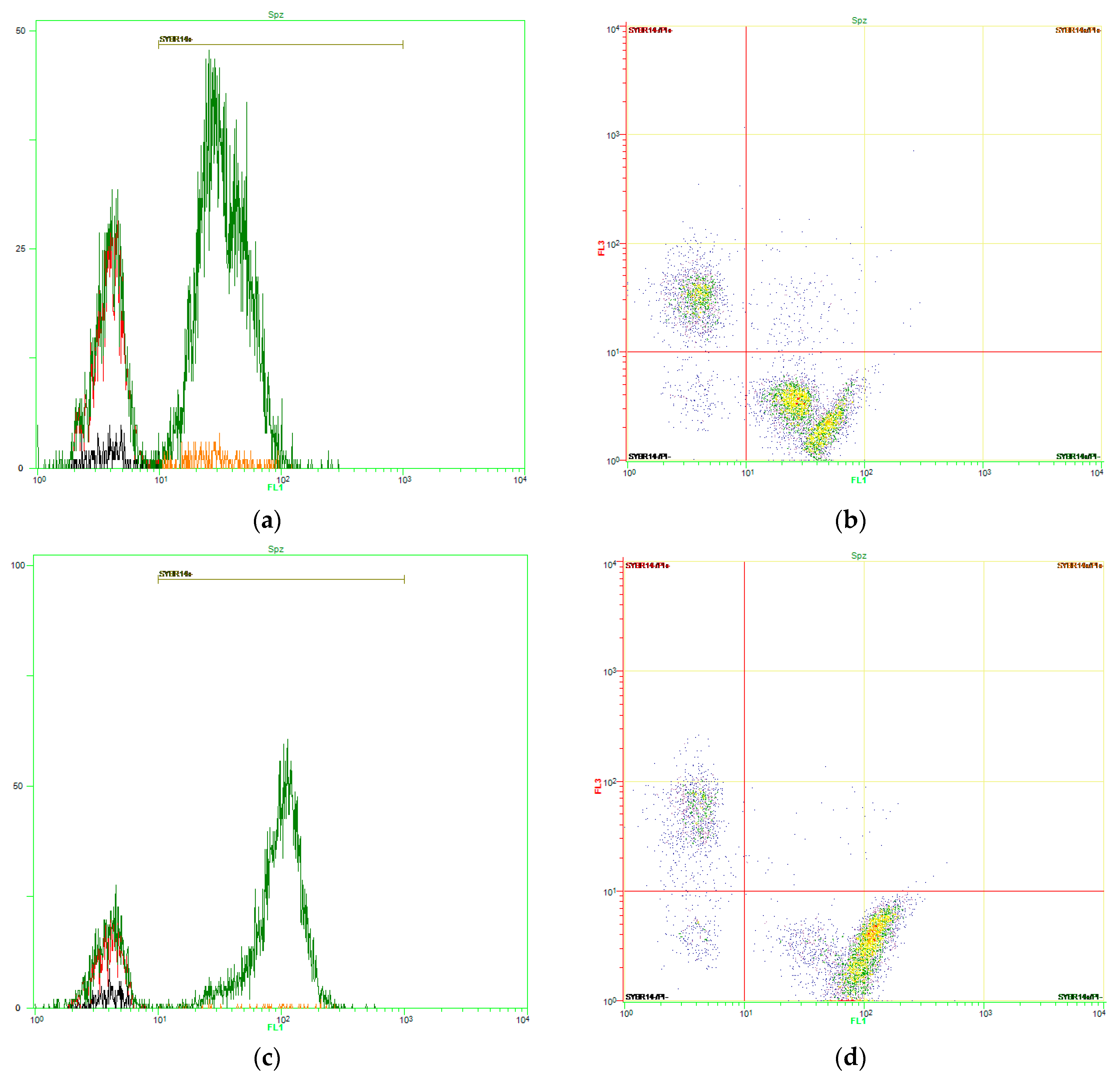
2.5. Flow Cytometry Analyses
2.5.1. General Information about Flow Cytometry Analyses
2.5.2. Plasma Membrane Integrity
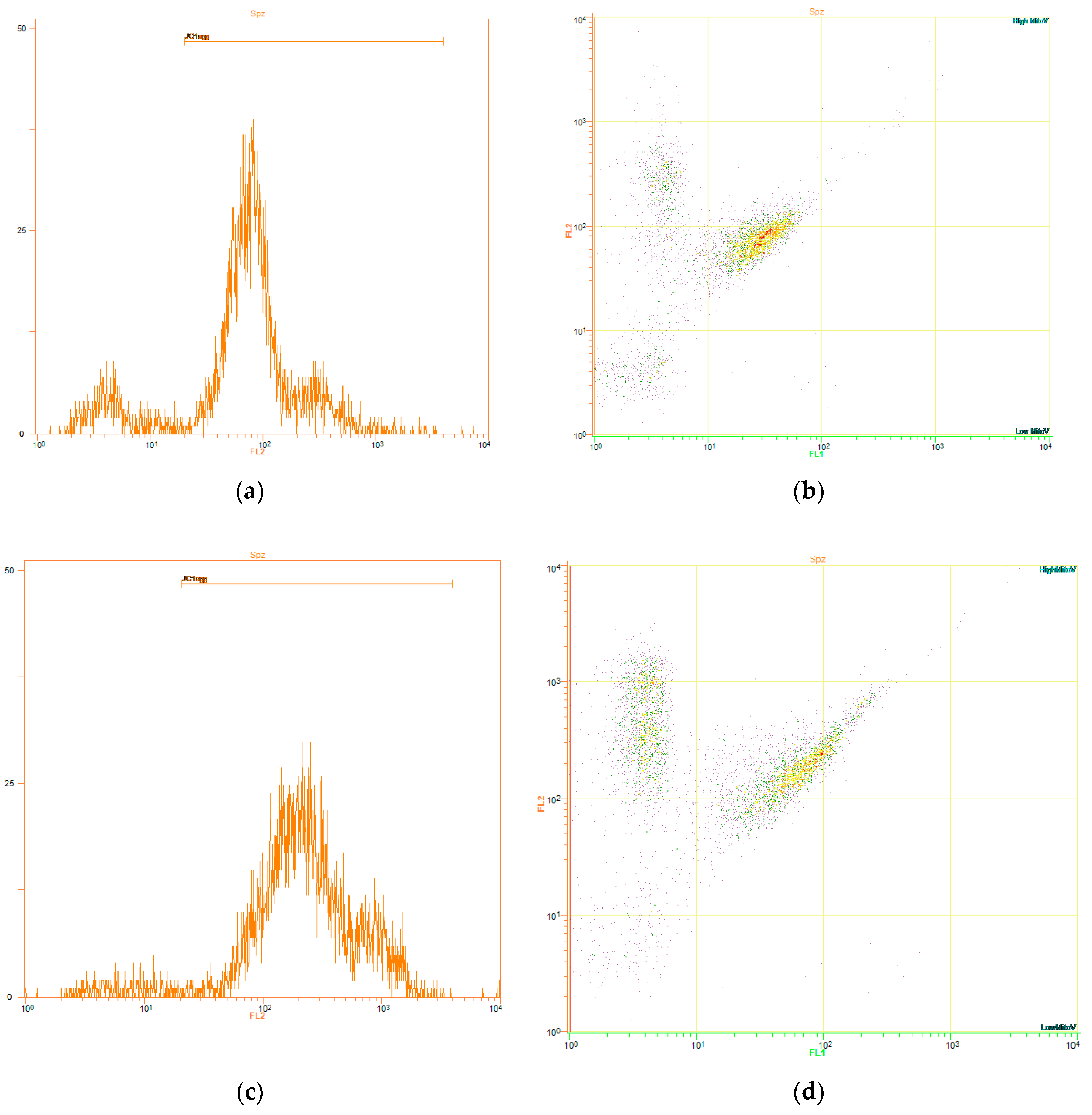
2.5.3. Evaluation of Mitochondrial Membrane Potential (∆Ψm, JC1)
2.5.4. DNA Integrity (SCSA Test)
2.6. Determination of Intracellular ATP Levels
2.7. Determination of O2 Consumption Rate
2.8. Statistical Analyses
3. Results
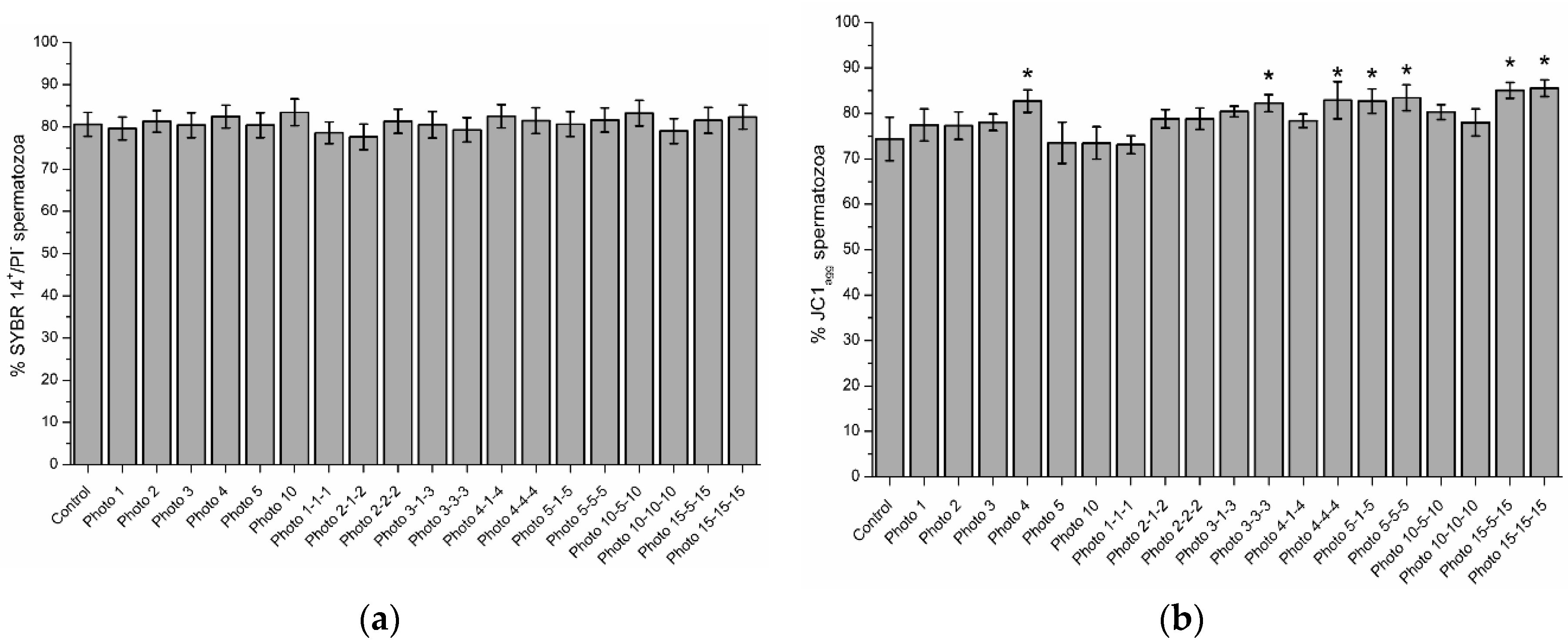
3.1. Effects of Red-Light Irradiation on Sperm Viability
3.2. Effects of Red-Light Irradiation on Mitochondrial Membrane Potential (ΔΨm)
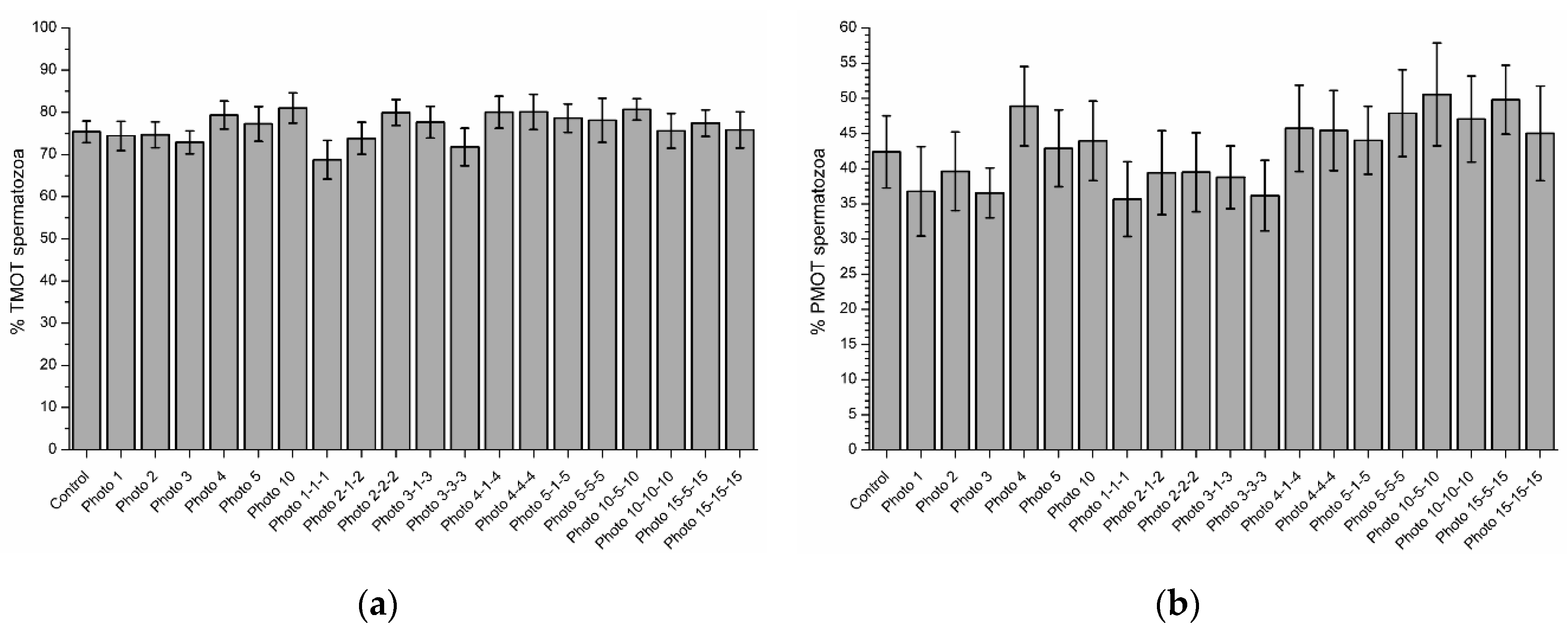
3.3. Effects of Red-Light Irradiation on Sperm Motility
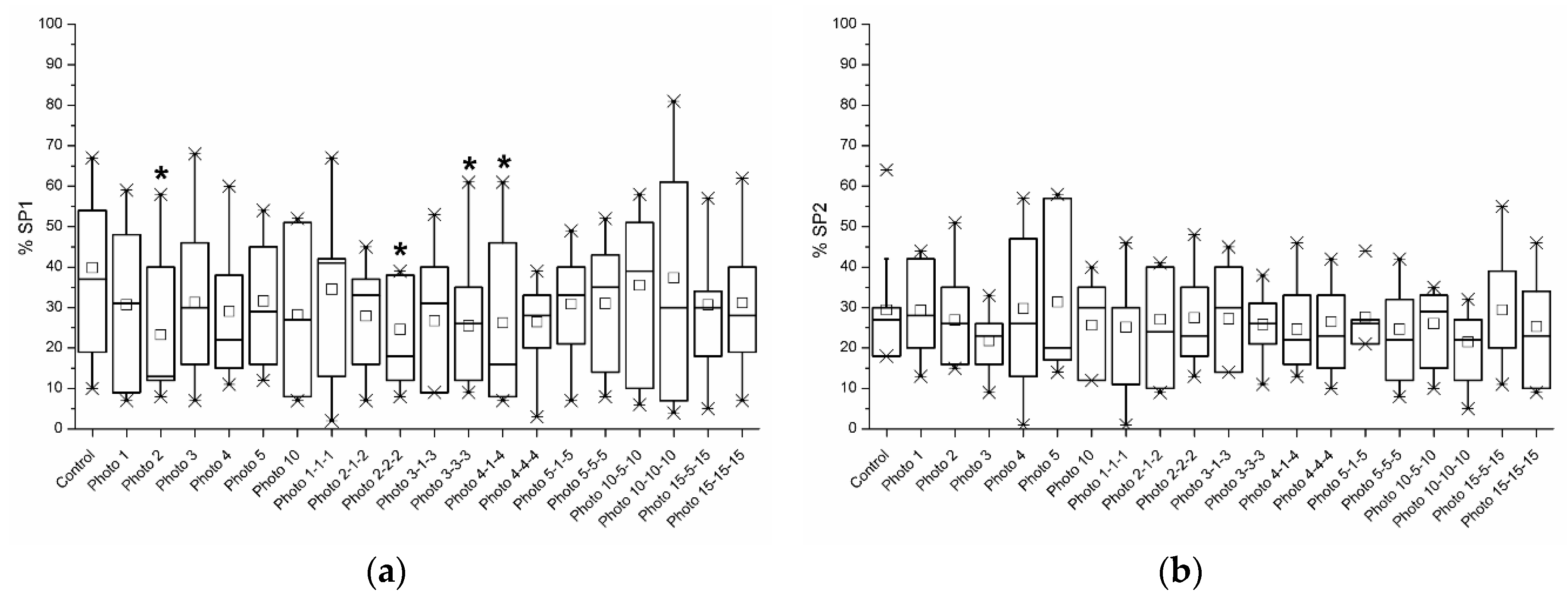
3.4. Sperm Subpopulations
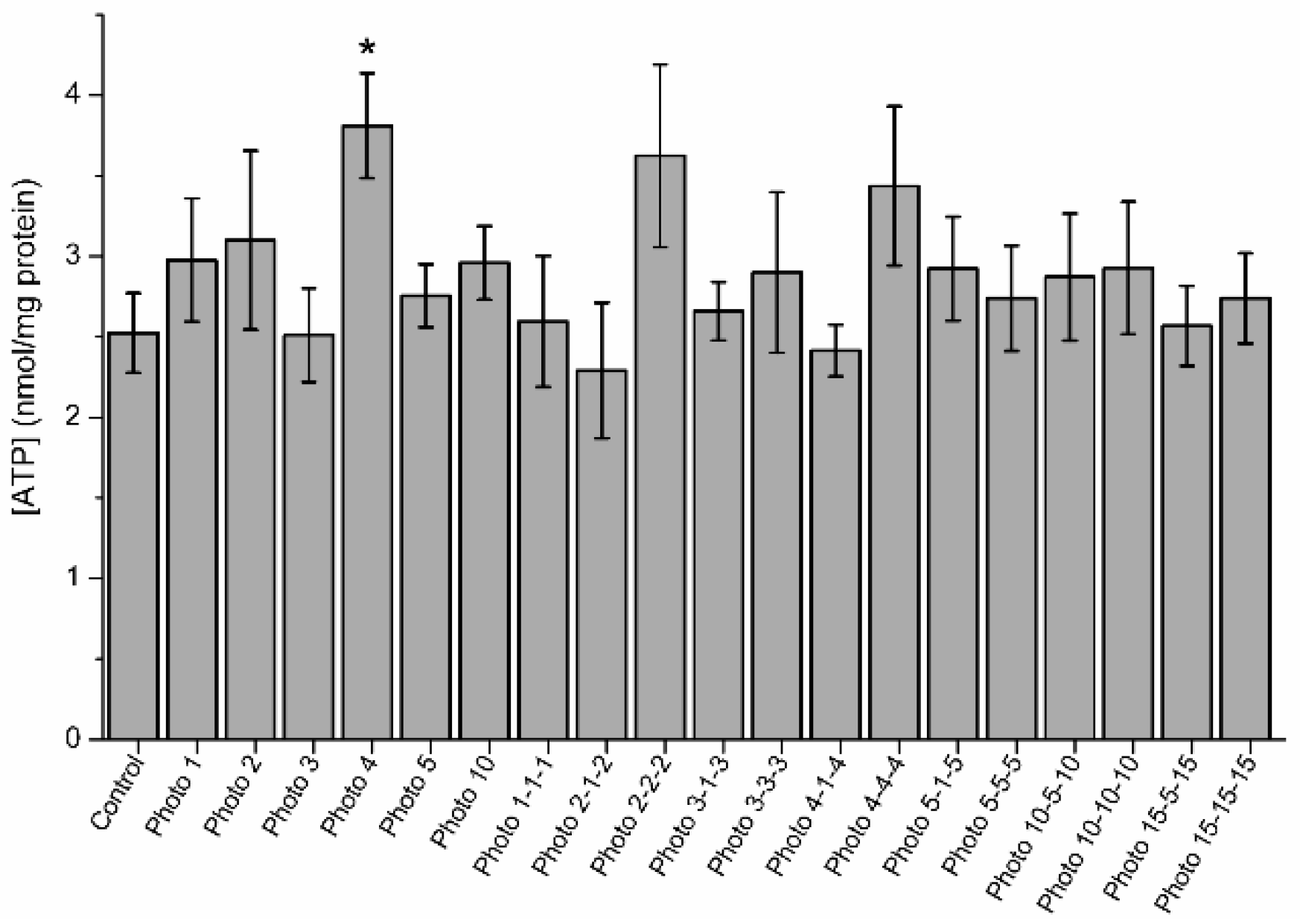
3.5. Effects of Red-Light Stimulation on Intracellular ATP Levels
3.6. Effects of Red-Light Stimulation on Oxygen Consumption
3.7. Effects of Red-Light Stimulation on DNA Fragmentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brinsko, S.P.; Varner, D.D. Artificial insemination and preservation of semen. Vet. Clin. N. Am. Equine Pract. 1992, 8, 205–218. [Google Scholar] [CrossRef]
- Aurich, J.E. Artificial Insemination in Horses-More than a Century of Practice and Research. J. Equine Vet. Sci. 2012, 32, 458–463. [Google Scholar] [CrossRef]
- Canisso, I.F.; Souza, F.A.; Marlén, J.; Escobar, O.; Ribeiro De Carvalho, G.; Davies Morel, M.C.; Capistrano Da Silva, E.; Domingos Guimarães, J.; Lima, A.L. Freezing of Donkey Semen (Equus Asinus). Rev. Investig. Vet. Perú 2008, 19, 113–125. [Google Scholar]
- Losinno, L.; Aguilar, J. Reproducción y Biotecnologías en la Producción Equina: Curso de Producción Equina I. 2002, pp. 1–4. Available online: http://www.produccion-animal.com.ar/produccion_equinos/curso_equinos_I/14-reproduccion_y_biotecnologias.pdf (accessed on 16 August 2020).
- Varner, D.D. Strategies for Processing Semen from Subfertile Stallions for Cooled Transport. Vet. Clin. N. Am. Equine Pract. 2016, 32, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Loomis, P.R. Advanced Methods for Handling and Preparation of Stallion Semen. Vet. Clin. N. Am. Equine Pract. 2006, 22, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Codony, F.; Estrada, E.; Lleonart, M.; Balasch, S.; Peña, A.; Bonet, S.; Rodríguez-Gil, J.E. Specific LED-based red light photo-stimulation procedures improve overall sperm function and reproductive performance of boar ejaculates. Sci. Rep. 2016, 6, 22569. [Google Scholar] [CrossRef] [Green Version]
- Blanco Prieto, O.; Catalán, J.; Lleonart, M.; Bonet, S.; Yeste, M.; Rodríguez-Gil, J.E. Red-light stimulation of boar semen prior to artificial insemination improves field fertility in farms: A worldwide survey. Reprod. Domest. Anim. 2019, 54, 1145–1148. [Google Scholar] [CrossRef]
- Pezo, F.; Zambrano, F.; Uribe, P.; Ramírez-Reveco, A.; Romero, F.; Sanchéz, R. LED-based red light photostimulation improves short-term response of cooled boar semen exposed to thermal stress at 37 °C. Andrologia 2019, 51, e13237. [Google Scholar] [CrossRef]
- Cohen, N.; Lubart, R.; Rubinstein, S.; Breitbart, H. Light Irradiation of Mouse Spermatozoa: Stimulation of In Vitro Fertilization and Calcium Signals. Photochem. Photobiol. 1998, 68, 407–413. [Google Scholar] [CrossRef]
- Zan-Bar, T.; Bartoov, B.; Segal, R.; Yehuda, R.; Lavi, R.; Lubart, R.; Avtalion, R.R. Influence of Visible Light and Ultraviolet Irradiation on Motility and Fertility of Mammalian and Fish Sperm. Photomed. Laser Surg. 2005, 23, 549–555. [Google Scholar] [CrossRef]
- Preece, D.; Chow, K.W.; Gomez-Godinez, V.; Gustafson, K.; Esener, S.; Ravida, N.; Durrant, B.; Berns, M.W. Red light improves spermatozoa motility and does not induce oxidative DNA damage. Sci. Rep. 2017, 7, 46480. [Google Scholar] [CrossRef] [PubMed]
- Gabel, C.P.; Carroll, J.; Harrison, K. Sperm motility is enhanced by Low Level Laser and Light Emitting Diode photobiomodulation with a dose-dependent response and differential effects in fresh and frozen samples. Laser Ther. 2018, 27, 131–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeste, M.; Castillo-Martín, M.; Bonet, S.; Rodríguez-Gil, J.E. Impact of light irradiation on preservation and function of mammalian spermatozoa. Anim. Reprod. Sci. 2018, 194, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Iaffaldano, N.; Paventi, G.; Pizzuto, R.; Passarella, S.; Cerolini, S.; Zaniboni, L.; Marzoni, M.; Castillo, A.; Rosato, M.P. The post-thaw irradiation of avian spermatozoa with He–Ne laser differently affects chicken, pheasant and turkey sperm quality. Anim. Reprod. Sci. 2013, 142, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Corral-Baqués, M.I.; Rigau, T.; Rivera, M.; Rodríguez, J.E.; Rigau, J. Effect of 655-nm diode laser on dog sperm motility. Lasers Med. Sci. 2005, 20, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Corral-Baqués, M.I.; Rivera, M.M.; Rigau, T.; Rodríguez-Gil, J.E.; Rigau, J. The effect of low-level laser irradiation on dog spermatozoa motility is dependent on laser output power. Lasers Med. Sci. 2009, 24, 703–713. [Google Scholar] [CrossRef]
- Iaffaldano, N.; Rosato, M.P.; Paventi, G.; Pizzuto, R.; Gambacorta, M.; Manchisi, A.; Passarella, S. The irradiation of rabbit sperm cells with He–Ne laser prevents their in vitro liquid storage dependent damage. Anim. Reprod. Sci. 2010, 119, 123–129. [Google Scholar] [CrossRef]
- Rodríguez-Gil, J.E. Photostimulation and thermotaxis of sperm: Overview and practical implications in porcine reproduction. Theriogenology 2019, 137, 8–14. [Google Scholar] [CrossRef]
- Karu, T. Photobiology of low-power laser effects. Health Phys. 1989, 56, 691–704. [Google Scholar] [CrossRef]
- Gao, X.; Xing, D. Molecular mechanisms of cell proliferation induced by low power laser irradiation. J. Biomed. Sci. 2009, 16, 4. [Google Scholar] [CrossRef] [Green Version]
- Kenney, M.R. Minimal contamination techniques for breeding mares: Techniques and priliminary findings. Proc. Am. Assoc. Equine Pract. 1975, 327–336. [Google Scholar]
- Lee, J.A.; Spidlen, J.; Boyce, K.; Cai, J.; Crosbie, N.; Dalphin, M.; Furlong, J.; Gasparetto, M.; Goldberg, M.; Goralczyk, E.M.; et al. MIFlowCyt: The minimum information about a flow cytometry experiment. Cytom. Part A 2008, 73A, 926–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrunkina, A.M.; Waberski, D.; Bollwein, H.; Sieme, H. Identifying non-sperm particles during flow cytometric physiological assessment: A simple approach. Theriogenology 2010, 73, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M.; Flores, E.; Estrada, E.; Bonet, S.; Rigau, T.; Rodríguez-Gil, J.E. Reduced glutathione and procaine hydrochloride protect the nucleoprotein structure of boar spermatozoa during freeze–thawing by stabilising disulfide bonds. Reprod. Fertil. Dev. 2013, 25, 1036. [Google Scholar] [CrossRef] [Green Version]
- Garner, D.L.; Johnson, L.A. Viability Assessment of Mammalian Sperm Using SYBR-14 and Propidium Iodide1. Biol. Reprod. 1995, 53, 276–284. [Google Scholar] [CrossRef]
- Ortega-Ferrusola, C.; Sotillo-Galan, Y.; Varela-Fernandez, E.; Gallardo-Bolanos, J.M.; Muriel, A.; Gonzalez-Fernandez, L.; Tapia, J.A.; Pena, F.J. Detection of “Apoptosis-Like” Changes During the Cryopreservation Process in Equine Sperm. J. Androl. 2007, 29, 213–221. [Google Scholar] [CrossRef]
- Evenson, D.P.; Darzynkiewicz, Z.; Melamed, M.R. Relation of mammalian sperm chromatin heterogeneity to fertility. Science 1980, 210, 1131–1133. [Google Scholar] [CrossRef]
- Evenson, D.P. Sperm Chromatin Structure Assay (SCSA®). Methods Mol. Biol. 2013, 927, 147–164. [Google Scholar] [CrossRef]
- Morrell, J.M.; Johannisson, A.; Dalin, A.M.; Hammar, L.; Sandebert, T.; Rodriguez-Martinez, H. Sperm morphology and chromatin integrity in Swedish warmblood stallions and their relationship to pregnancy rates. Acta Vet. Scand. 2008, 50, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chida, J.; Yamane, K.; Takei, T.; Kido, H. An efficient extraction method for quantitation of adenosine triphosphate in mammalian tissues and cells. Anal. Chim. Acta 2012, 727, 8–12. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Luna, C.; Yeste, M.; Rivera Del Alamo, M.M.; Domingo, J.; Casao, A.; Rodriguez-Gil, J.E.; Pérez-Pé, R.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. Effect of seminal plasma proteins on the motile sperm subpopulations in ram ejaculates. Reprod. Fertil. Dev. 2017, 29, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Salam, Z.; Dessouki, S.H.M.; Abdel-Salam, S.A.M.; Ibrahim, M.A.M.; Harith, M.A. Green laser irradiation effects on buffalo semen. Theriogenology 2011, 75, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Salman Yazdi, R.; Bakhshi, S.; Jannat Alipoor, F.; Akhoond, M.R.; Borhani, S.; Farrahi, F.; Lotfi Panah, M.; Sadighi Gilani, M.A. Effect of 830-nm diode laser irradiation on human sperm motility. Lasers Med. Sci. 2014, 29, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R.; Huang, Y.Y.; Sharma, S.K.; Carroll, J. Biphasic dose response in low level light therapy-an update. Dose Response 2011, 9, 602–618. [Google Scholar] [CrossRef]
- Ban Frangez, H.; Frangez, I.; Verdenik, I.; Jansa, V.; Virant Klun, I. Photobiomodulation with light-emitting diodes improves sperm motility in men with asthenozoospermia. Lasers Med. Sci. 2015, 30, 235–240. [Google Scholar] [CrossRef]
- Huang, Y.-Y.; Chen, A.C.-H.; Carroll, J.D.; Hamblin, M.R. Biphasic dose response in low level light therapy. Dose Response 2009, 7, 358–383. [Google Scholar] [CrossRef]
- Lubart, R.; Lavi, R.; Friedmann, H.; Rochkind, S. Photochemistry and Photobiology of Light Absorption by Living Cells. Photomed. Laser Surg. 2006, 24, 179–185. [Google Scholar] [CrossRef]
- Chow, R.T.; Heller, G.Z.; Barnsley, L. The effect of 300 mW, 830 nm laser on chronic neck pain: A double-blind, randomized, placebo-controlled study. Pain 2006, 124, 201–210. [Google Scholar] [CrossRef]
- Firestone, R.S.; Esfandiari, N.; Moskovtsev, S.I.; Burstein, E.; Videna, G.T.; Librach, C.; Bentov, Y.; Casper, R.F. The Effects of Low-Level Laser Light Exposure on Sperm Motion Characteristics and DNA Damage. J. Androl. 2012, 33, 469–473. [Google Scholar] [CrossRef]
- Lenzi, A.; Claroni, F.; Gandini, L.; Lombardo, F.; Barbieri, C.; Lino, A.; Dondero, F. Laser Radiation and Motility Patterns of Human Sperm. Arch. Androl. 1989, 23, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, A.F.P.; Maria, F.S.; Mendes, C.M.; Hamilton, T.R.S.; Dalmazzo, A.; Dreyer, T.R.; da Silva, H.M.; Nichi, M.; Milazzotto, M.P.; Visintin, J.A.; et al. Effects of photobiomodulation therapy (PBMT) on bovine sperm function. Lasers Med. Sci. 2016, 31, 1245–1250. [Google Scholar] [CrossRef]
- Sato, H.; Landthaler, M.; Haina, D.; Schill, W.B. The effects of laser light on sperm motility and velocity in vitro. Andrologia 1984, 16, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Iaffaldano, N.; Paventi, G.; Pizzuto, R.; Di Iorio, M.; Bailey, J.L.; Manchisi, A.; Passarella, S. Helium-neon laser irradiation of cryopreserved ram sperm enhances cytochrome c oxidase activity and ATP levels improving semen quality. Theriogenology 2016, 86, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Moreno, A.; Miró, J.; Teresa Rigau, A.; Rodríguez-Gil, J.E. Identification of sperm subpopulations with specific motility characteristics in stallion ejaculates. Theriogenology 2003, 59, 1973–1990. [Google Scholar] [CrossRef]
- Miró, J.; Lobo, V.; Quintero-Moreno, A.; Medrano, A.; Peña, A.; Rigau, T. Sperm motility patterns and metabolism in Catalonian donkey semen. Theriogenology 2005, 63, 1706–1716. [Google Scholar] [CrossRef]
- Muiño, R.; Tamargo, C.; Hidalgo, C.O.; Peña, A.I. Identification of sperm subpopulations with defined motility characteristics in ejaculates from Holstein bulls: Effects of cryopreservation and between-bull variation. Anim. Reprod. Sci. 2008, 109, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Corredor, L.H.; Rodríguez, O.C.; Torres, A.S.; Páez, J.D.M.; Moreno, A.Q. Effects of cryopreservation on sperm subpopulations in goats. Rev. Investig. Vet. Peru 2018, 29, 882–893. [Google Scholar] [CrossRef] [Green Version]
- Dorado, J.; Molina, I.; Muñoz-Serrano, A.; Hidalgo, M. Identification of sperm subpopulations with defined motility characteristics in ejaculates from Florida goats. Theriogenology 2010, 74, 795–804. [Google Scholar] [CrossRef]
- Begum, R.; Powner, M.B.; Hudson, N.; Hogg, C.; Jeffery, G. Treatment with 670 nm Light Up Regulates Cytochrome C Oxidase Expression and Reduces Inflammation in an Age-Related Macular Degeneration Model. PLoS ONE 2013, 8, e57828. [Google Scholar] [CrossRef] [Green Version]
- Breitbart, H.; Levinshal, T.; Cohen, N.; Friedmann, H.; Lubart, R. Changes in calcium transport in mammalian sperm mitochondria and plasma membrane irradiated at 633 nm (HeNe laser). J. Photochem. Photobiol. B Biol. 1996, 34, 117–121. [Google Scholar] [CrossRef]
- Moscatelli, N.; Spagnolo, B.; Pisanello, M.; Lemma, E.D.; De Vittorio, M.; Zara, V.; Pisanello, F.; Ferramosca, A. Single-cell-based evaluation of sperm progressive motility via fluorescent assessment of mitochondria membrane potential. Sci. Rep. 2017, 7, 17931. [Google Scholar] [CrossRef]
- Meyers, S.; Bulkeley, E.; Foutouhi, A. Sperm mitochondrial regulation in motility and fertility in horses. Reprod. Domest. Anim. 2019, 54, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Energy Conversion: Mitochondria and Chloroplasts. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002; pp. 753–812. [Google Scholar]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patterns | VCL (µm/s) | VSL (µm/s) | VAP (µm/s) | LIN (%) | STR (%) | WOB (%) | ALH (µm) | BCF (Hz) |
|---|---|---|---|---|---|---|---|---|
| Control | 107.2 ± 5.9 a | 56.9 ± 2.5 a | 81.9 ± 4.7 a | 55.6 ± 4.7 a | 71.0 ± 4.6 a | 77.6 ± 2.1 a | 2.7 ± 0.1 a | 9.7 ± 0.6 a |
| Ph 1 | 116.0 ± 8.3 a | 58.9 ± 3.9 a | 88.5 ± 4.7 a | 52.2 ± 6.2 a | 66.3 ± 5.7 a | 77.2 ± 2.8 a | 2.9 ± 0.2 a | 9.3 ± 0.4 a |
| Ph 2 | 117.2 ± 4.8 a | 62.3 ± 5.3 a | 92.0 ± 4.9 a | 53.6 ± 4.8 a | 68.0 ± 4.2 a | 78.1 ± 2.6 a | 2.9 ± 0.1 a | 9.1 ± 0.3 a |
| Ph 3 | 113.2 ± 7.0 a | 60.7 ± 4.0 a | 90.4 ± 6.3 a | 54.5 ± 4.2 a | 68.2 ± 4.5 a | 79.6 ± 2.1 a | 2.9 ± 0.2 a | 9.1 ± 0.2 a |
| Ph 4 | 117.7 ± 5.2 a | 62.5 ± 4.5 a | 94.4 ± 4.6 b | 53.2 ± 4.6 a | 66.2 ± 4.8 a | 80.2 ± 2.2 a | 3.0 ± 0.3 a | 9.2 ± 0.8 a |
| Ph 5 | 111.3 ± 6.6 a | 58.9 ± 2.6 a | 87.7 ± 4.3 a | 54.8 ± 5.0 a | 68.6 ± 5.0 a | 79.2 ± 2.0 a | 2.6 ± 0.1 a | 9.4 ± 0.5 a |
| Ph 10 | 115.4 ± 6.9 a | 62.6 ± 3.3 a | 91.8 ± 6.3 a | 55.4 ± 4.2 a | 69.4 ± 4.3 a | 79.5 ± 2.0 a | 2.9 ± 0.1 a | 9.3 ± 0.2 a |
| Ph 1-1-1 | 112.4 ± 8.5 a | 60.7 ± 2.5 a | 84.6 ± 5.7 a | 55.0 ± 5.6 a | 70.6 ± 5.3 a | 77.0 ± 2.3 a | 3.2 ± 0.3 a | 9.1 ± 0.7 a |
| Ph 2-1-2 | 116.3 ± 7.4 a | 62.9 ± 3.3 a | 90.6 ± 4.5 a | 56.1 ± 5.5 a | 70.6 ± 5.0 a | 78.6 ± 2.4 a | 3.2 ± 0.3 a | 8.8 ± 0.6 a |
| Ph 2-2-2 | 121.6 ± 7.3 a | 62.4 ± 3.4 a | 96.0 ± 4.7 b | 53.5 ± 5.0 a | 65.9 ± 4.6 a | 79.6 ± 2.5 a | 2.9 ± 0.3 a | 8.6 ± 0.8 a |
| Ph 3-1-3 | 115.6 ± 6.8 a | 61.1 ± 3.5 a | 91.4 ± 6.2 a | 53.7 ± 3.5 a | 68.2 ± 4.1 a | 78.6 ± 1.8 a | 3.3 ± 0.3 a | 8.6 ± 0.6 a |
| Ph 3-3-3 | 119.5 ± 6.9 a | 65.7 ± 4.8 b | 96.4 ± 5.6 b | 55.6 ± 4.8 a | 68.0 ± 4.7 a | 79.7 ± 2.5 a | 3.1 ± 0.3 a | 8.4 ± 0.6 b |
| Ph 4-1-4 | 112.3 ± 7.7 a | 63.9 ± 2.8 a | 93.3 ± 5.7 a | 59.6 ± 4.5 a | 72.6 ± 5.0 a | 81.9 ± 1.4 a | 2.8 ± 0.1 a | 9.6 ± 0.3 a |
| Ph 4-4-4 | 112.0 ± 7.6 a | 64.4 ± 3.2 a | 94.8 ± 3.9 b | 59.0 ± 4.3 a | 71.6 ± 4.7 a | 82.2 ± 1.7 a | 2.6 ± 0.1 a | 9.0 ± 0.5 a |
| Ph 5-1-5 | 113.0 ± 6.8 a | 64.0 ± 2.0 a | 91.0 ± 4.1 a | 58.1 ± 4.0 a | 71.2 ± 3.8 a | 81.1 ± 1.9 a | 3.1 ± 0.3 a | 8.4 ± 0.6 b |
| Ph 5-5-5 | 112.4 ± 7.4 a | 66.7 ± 2.0 b | 92.7 ± 4.3 a | 61.3 ± 4.4 a | 74.4 ± 4.4 a | 82.4 ± 1.5 a | 2.9 ± 0.1 a | 8.9 ± 0.3 a |
| Ph 10-5-10 | 105.8 ± 8.3 a | 61.3 ± 2.5 a | 86.0 ± 5.4 a | 60.8 ± 5.4 a | 73.7 ± 5.8 a | 81.9 ± 1.7 a | 2.5 ± 0.1 a | 9.1 ± 0.3 a |
| Ph 10-10-10 | 109.4 ± 12.3 a | 58.1 ± 5.1 a | 84.0 ± 8.8 a | 57.3 ± 6.5 a | 73.0 ± 6.9 a | 77.4 ± 3.4 a | 2.8 ± 0.3 a | 9.7 ± 0.6 a |
| Ph 15-5-15 | 109.2 ± 8.9 a | 60.5 ± 3.4 a | 83.6 ± 4.9 a | 58.6 ± 6.2 a | 74.3 ± 6.1 a | 77.4 ± 2.8 a | 3.0 ± 0.2 a | 9.8 ± 0.4 a |
| Ph 15-15-15 | 112.2 ± 9.4 a | 59.6 ± 6.1 a | 85.7 ± 5.1 a | 56.7 ± 7.4 a | 70.9 ± 7.5 a | 77.7 ± 3.7 a | 2.8 ± 0.2 a | 9.7 ± 0.3 a |
| SP1 | SP2 | SP3 | SP4 | |||||
|---|---|---|---|---|---|---|---|---|
| N | 11,893 | 10,098 | 11,210 | 8142 | ||||
| Parameter | Mean ± SEM | Range | Mean ± SEM | Range | Mean ± SEM | Range | Mean ± SEM | Range |
| VCL (µm/s) | 72.7 ± 0.1 | 0.0–120.0 | 109.1 ± 0.1 | 71.6–181.9 | 120.1 ± 0.2 | 87.2–213.9 | 158.3 ± 0.3 | 83.1–372.2 |
| VSL (µm/s) | 45.9 ± 0.1 | 0.0–84.6 | 64.7 ± 0.2 | 0.9-105.6 | 94.5 ± 0.2 | 43.4–199.0 | 47.2 ± 0.3 | 0.6–221.2 |
| VAP (µm/s) | 59.9 ± 0.1 | 0.0–112.9 | 85.1 ± 0.1 | 34.4–129.3 | 112.7 ± 0.2 | 83.9–224.8 | 109.5 ± 0.2 | 24.1-282.5 |
| LIN (%) | 63.2 ± 0.2 | 0.0–100.0 | 60.7 ± 0.2 | 0.8–97.7 | 79.4 ± 0.1 | 33.8–99.3 | 30.0 ± 0.2 | 0.4–84.4 |
| STR (%) | 72.1 ± 0.2 | 0.0–100.0 | 76.6 ± 0.2 | 1.4–99.7 | 84.3 ± 0.1 | 35.4–100.0 | 43.7 ± 0.2 | 0.5–98.3 |
| WOB (%) | 80.6 ± 0.1 | 0.0–100.0 | 78.6 ± 0.1 | 28.1–100.0 | 93.8 ± 0.1 | 71.2–100.0 | 69.7 ± 0.2 | 17.6–100.0 |
| ALH (µm) | 2.2 ± 0.1 | 0.0–5.9 | 3.5 ± 0.1 | 0.9–6.6 | 2.5 ± 0.1 | 0.4–5.30 | 5.7 ± 0.1 | 2.0–16.9 |
| BCF (Hz) | 7.7 ± 0.1 | 0.0–21.0 | 11.0 ± 0.1 | 3.6–22.0 | 7.9 ± 0.1 | 0.0–20.0 | 8.4 ± 0.1 | 0.0–22.0 |
| Patterns | % DFI | Mean DFI | % HDS |
|---|---|---|---|
| Control | 15.4 ± 3.0 | 322.9 ± 30.1 | 21.1 ± 5.1 |
| Photo 1 | 15.4 ± 0.8 | 335.5 ± 33.3 | 22.8 ± 8.9 |
| Photo 2 | 15.4 ± 1.2 | 332.1 ± 19.5 | 22.8 ± 7.6 |
| Photo 3 | 15.9 ± 2.9 | 327.5 ± 25.4 | 22.2 ± 5.1 |
| Photo 4 | 14.2 ± 1.4 | 319.0 ± 21.3 | 18.1 ± 5.6 |
| Photo 5 | 15.8 ± 1.4 | 326.9 ± 29.9 | 19.7 ± 8.9 |
| Photo 10 | 14.6 ± 0.9 | 319.7 ± 25.4 | 19.3 ± 8.5 |
| Photo 1-1-1 | 17.0 ± 1.4 | 326.0 ± 34.8 | 25.3 ± 8.1 |
| Photo 2-1-2 | 15.6 ± 0.7 | 319.7 ± 29.0 | 21.1 ± 5.8 |
| Photo 2-2-2 | 17.1 ± 1.7 | 332.6 ± 30.1 | 25.9 ± 6.1 |
| Photo 3-1-3 | 16.4 ± 2.9 | 351.7 ± 46.2 | 22.7 ± 8.9 |
| Photo 3-3-3 | 15.8 ± 2.9 | 330.4 ± 30.2 | 21.8 ± 3.1 |
| Photo 4-1-4 | 14.3 ± 1.0 | 332.5 ± 26.4 | 18.7 ± 8.3 |
| Photo 4-4-4 | 16.8 ± 2.9 | 357.7 ± 44.1 | 32.3 ± 7.2 |
| Photo 5-1-5 | 17.5 ± 3.9 | 323.7 ± 28.1 | 28.2 ± 12.2 |
| Photo 5-5-5 | 17.5 ± 2.7 | 350.0 ± 37.5 | 31.6 ± 6.0 |
| Photo 10-5-10 | 18.4 ± 3.8 | 341.4 ± 27.0 | 31.9 ± 7.0 |
| Photo 10-10-10 | 18.4 ± 3.3 | 324.6 ± 27.4 | 31.4 ± 10.4 |
| Photo 15-5-15 | 19.3 ± 4.5 | 313.7 ± 24.3 | 29.7 ± 9.2 |
| Photo 15-15-15 | 18.3 ± 4.3 | 314.5 ± 19.3 | 25.1 ± 9.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catalán, J.; Papas, M.; Gacem, S.; Mateo-Otero, Y.; Rodríguez-Gil, J.E.; Miró, J.; Yeste, M. Red-Light Irradiation of Horse Spermatozoa Increases Mitochondrial Activity and Motility through Changes in the Motile Sperm Subpopulation Structure. Biology 2020, 9, 254. https://doi.org/10.3390/biology9090254
Catalán J, Papas M, Gacem S, Mateo-Otero Y, Rodríguez-Gil JE, Miró J, Yeste M. Red-Light Irradiation of Horse Spermatozoa Increases Mitochondrial Activity and Motility through Changes in the Motile Sperm Subpopulation Structure. Biology. 2020; 9(9):254. https://doi.org/10.3390/biology9090254
Chicago/Turabian StyleCatalán, Jaime, Marion Papas, Sabrina Gacem, Yentel Mateo-Otero, Joan E. Rodríguez-Gil, Jordi Miró, and Marc Yeste. 2020. "Red-Light Irradiation of Horse Spermatozoa Increases Mitochondrial Activity and Motility through Changes in the Motile Sperm Subpopulation Structure" Biology 9, no. 9: 254. https://doi.org/10.3390/biology9090254
APA StyleCatalán, J., Papas, M., Gacem, S., Mateo-Otero, Y., Rodríguez-Gil, J. E., Miró, J., & Yeste, M. (2020). Red-Light Irradiation of Horse Spermatozoa Increases Mitochondrial Activity and Motility through Changes in the Motile Sperm Subpopulation Structure. Biology, 9(9), 254. https://doi.org/10.3390/biology9090254