The Teleost Thymus in Health and Disease: New Insights from Transcriptomic and Histopathological Analyses of Turbot, Scophthalmus maximus

 , ,
, ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Histopathology
2.2. Immunohistochemistry
2.3. Transcriptome Analysis
3. Results
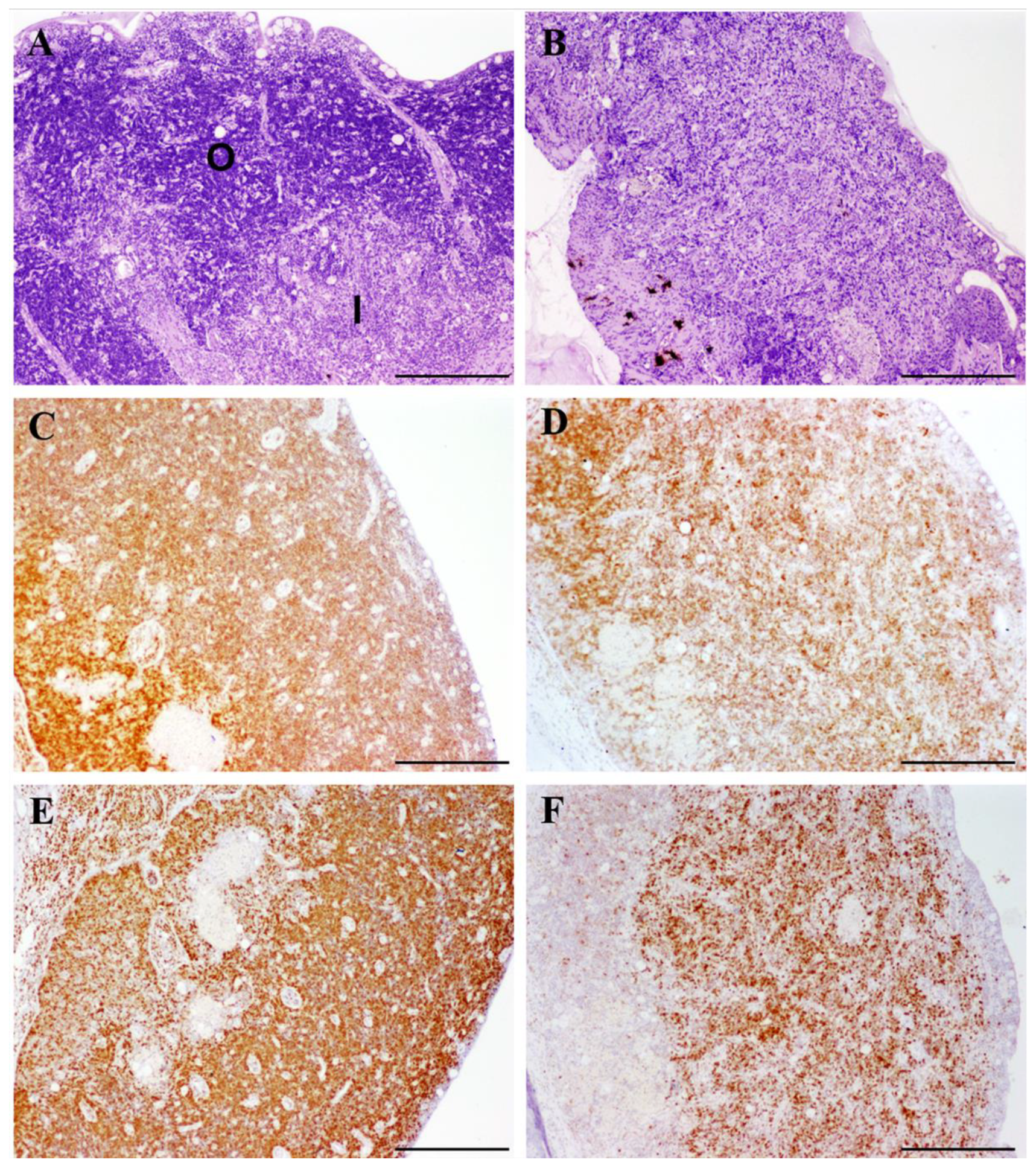
3.1. Histopathological Evaluation of Thymuses
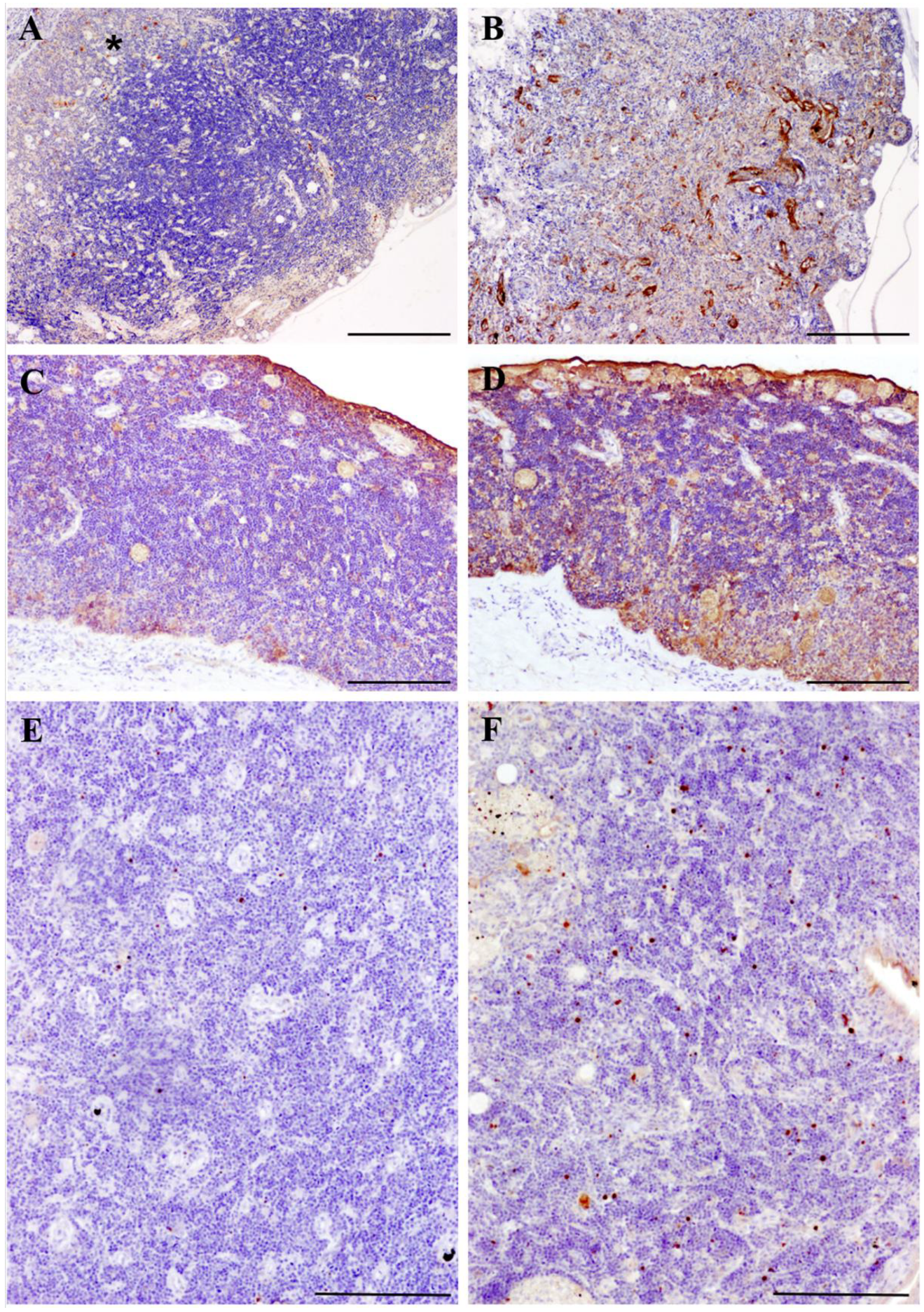
3.2. Immunohistochemistry
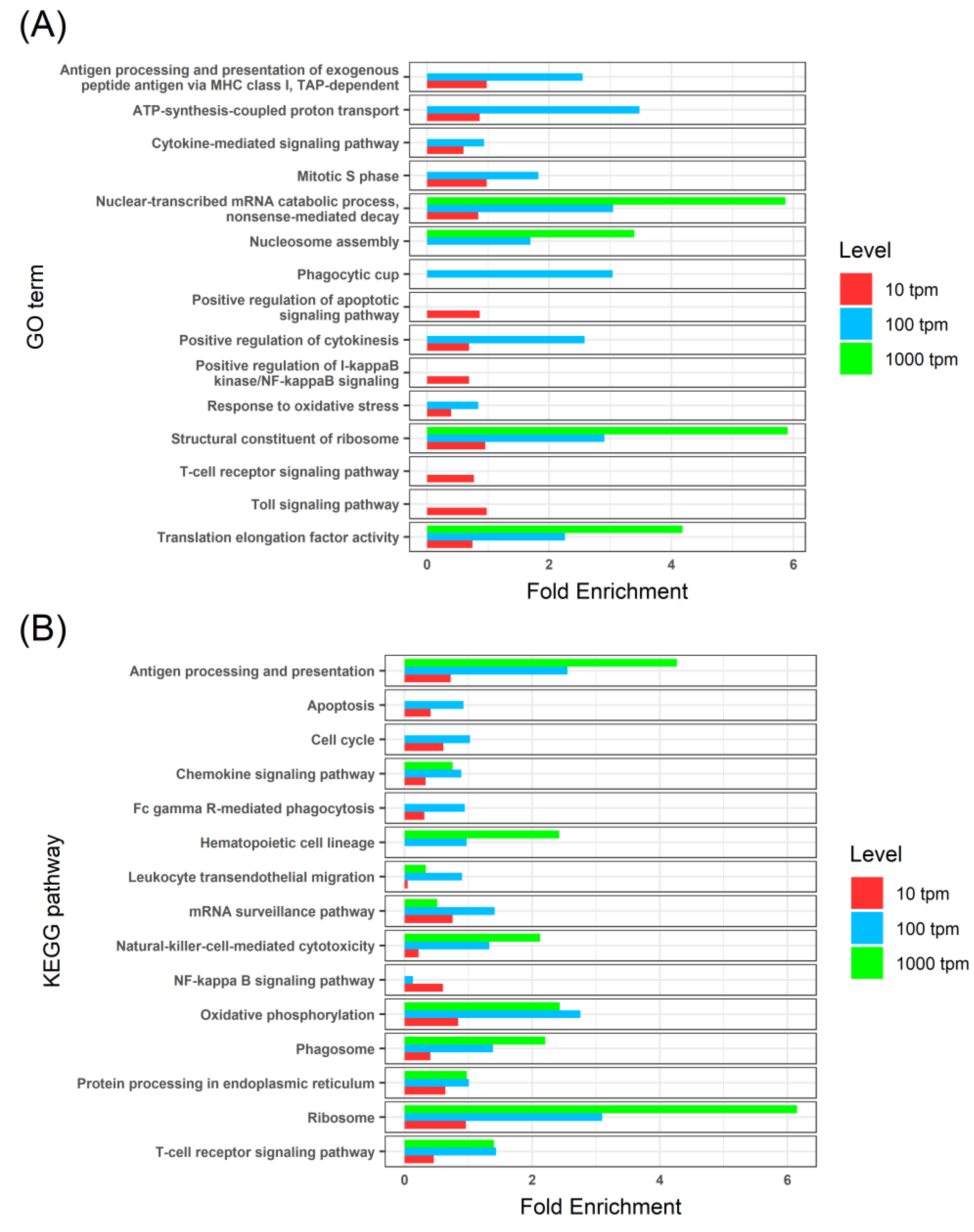
3.3. Thymus Transcriptome of Healthy Fish
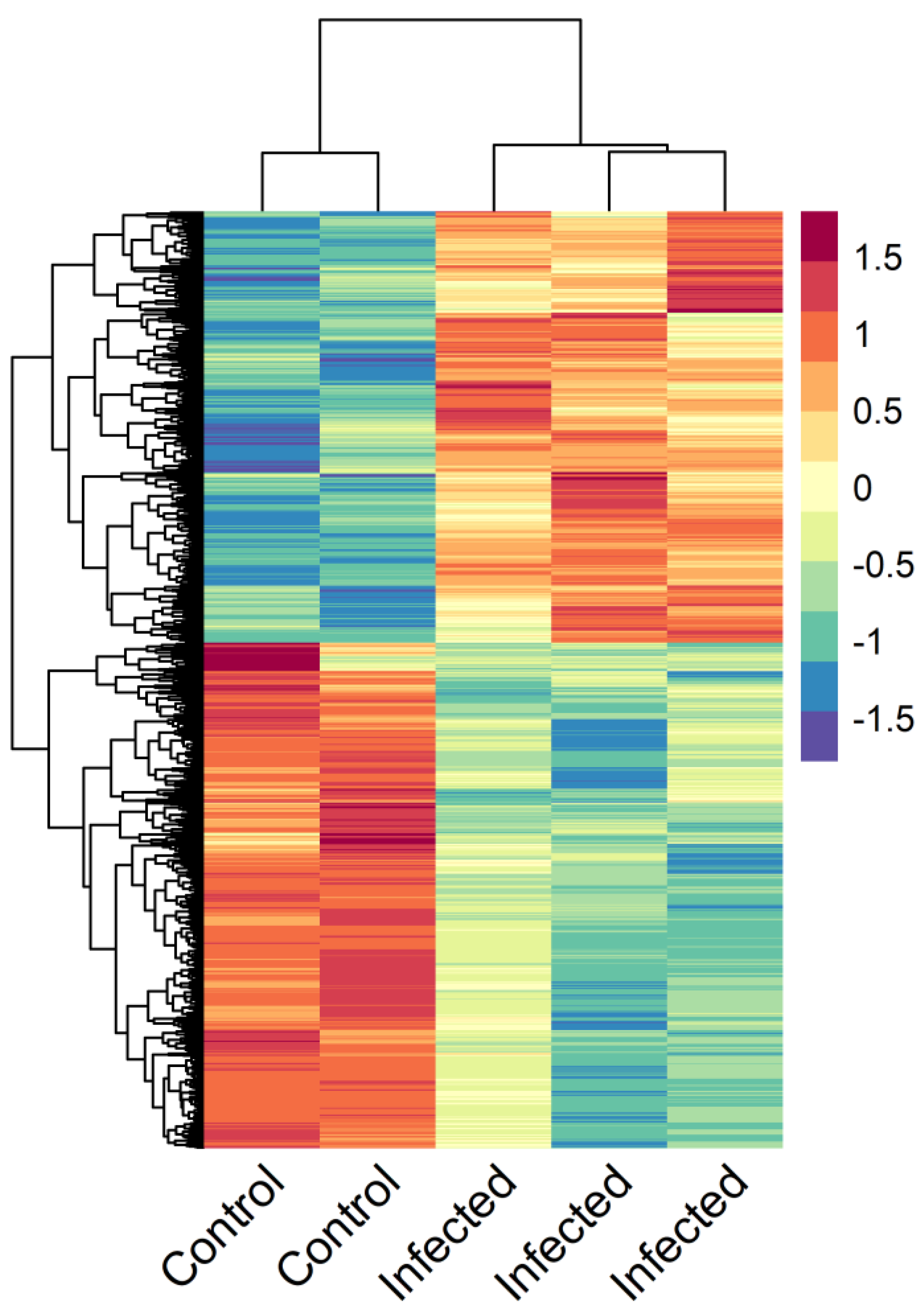
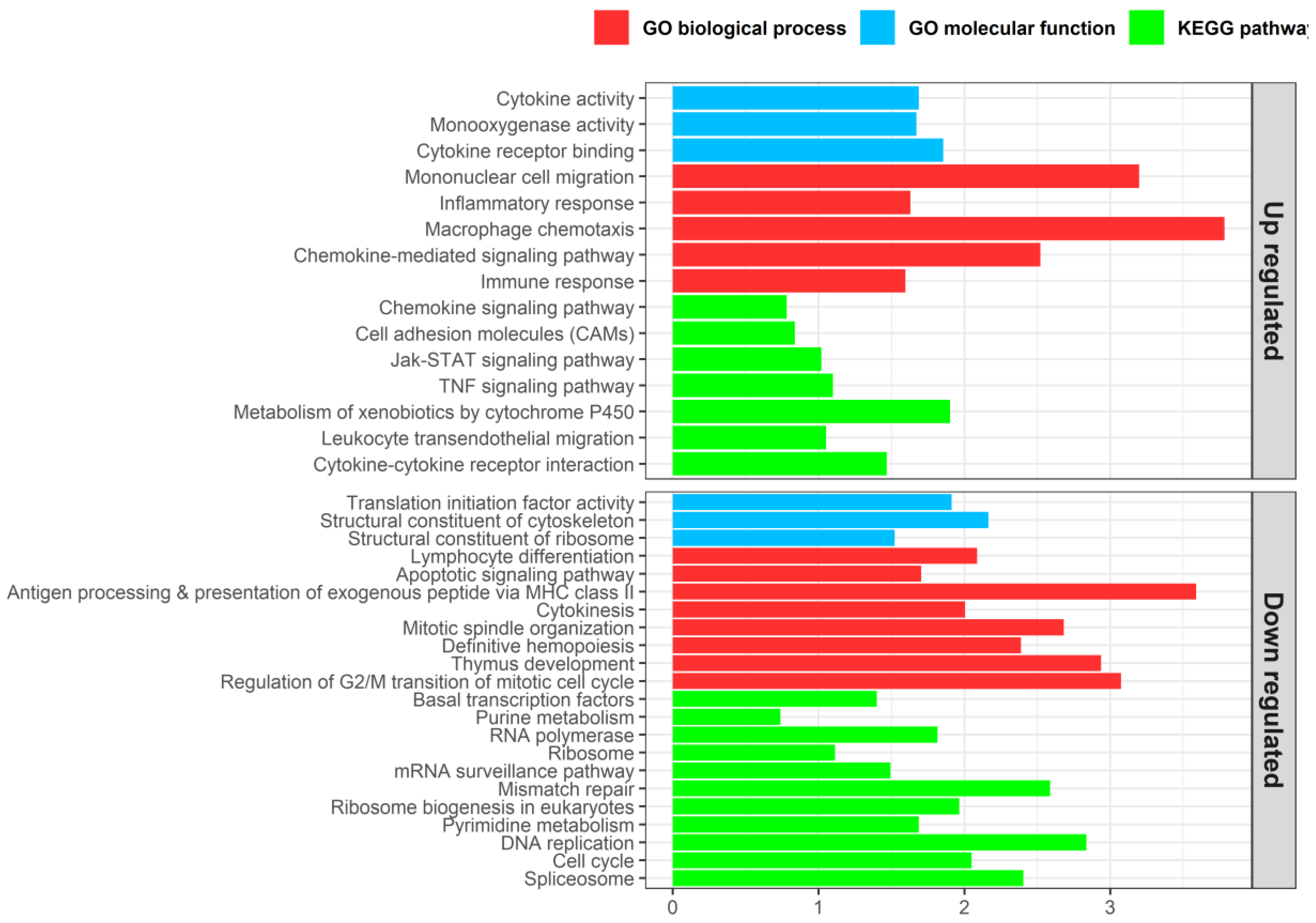
3.4. Enteromyxosis-Induced Transcriptomic Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miller, J.F.A.P. The golden anniversary of the thymus. Nat. Rev. Immunol. 2011, 11, 489–495. [Google Scholar] [CrossRef]
- Lancaster, J.N.; Li, Y.; Ehrlich, L.I. Chemokine-mediated choreography of thymocyte development and selection. Trends Immunol. 2018, 39, 86–98. [Google Scholar] [CrossRef]
- Savino, W.; Mendes-da-Cruz, D.A.; Smaniotto, S.; Silva-Monteiro, E.; Villa-Verde, D.M.S. Molecular mechanisms governing thymocyte migration: Combined role of chemokines and extracellular matrix. J. Leukoc. Biol. 2004, 75, 951–961. [Google Scholar] [CrossRef]
- Takaba, H.; Takayanagi, H. The mechanisms of T cell selection in the thymus. Trends Immunol. 2017, 38, 805–816. [Google Scholar] [CrossRef]
- Klein, L.; Kyewski, B.; Allen, P.M.; Hogquist, K.A. Positive and negative selection of the T cell repertoire: What thymocytes see (and don’t see). Nat. Rev. Immunol. 2014, 14, 377–391. [Google Scholar] [CrossRef]
- Bowden, T.J.; Cook, P.; Rombout, J.H. Development and function of the thymus in teleosts. Fish Shellfish Immunol. 2005, 19, 413–427. [Google Scholar] [CrossRef]
- Nakanishi, T.; Shibasaki, Y.; Matsuura, Y. T cells in fish. Biology 2015, 4, 640–663. [Google Scholar] [CrossRef]
- Carneiro-Sampaio, M. Thymus: Still an underexplored organ in medical practice. Rev. Med. São Paulo 2016, 95, 76–83. [Google Scholar] [CrossRef]
- Nunes-Alves, C.; Nobrega, C.; Behar, S.M.; Correia-Neves, M. Tolerance has its limits: How the thymus copes with infection. Trends Immunol. 2013, 34, 502–510. [Google Scholar] [CrossRef]
- Valli, V.E.O.; Kiupel, M.; Bienzle, D.; Wood, R.D. Chapter 2—Hematopoietic System. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: London, UK, 2016; Volume 3, pp. 102–268.e1. [Google Scholar]
- Savino, W.; Dardenne, M.; Velloso, L.A.; Silva-Barbosa, S.D. The thymus is a common target in malnutrition and infection. Br. J. Nutr. 2007, 98, S11–S16. [Google Scholar] [CrossRef]
- Majumdar, S.; Nandi, D. Thymic atrophy: Experimental studies and therapeutic interventions. Scand. J. Immunol. 2018, 87, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Losada-Barragán, M.; Umaña-Pérez, A.; Cuervo-Escobar, S.; Berbert, L.R.; Porrozzi, R.; Morgado, F.N.; Mendes-da-Cruz, D.A.; Savino, W.; Sánchez-Gómez, M.; Cuervo, P. Protein malnutrition promotes dysregulation of molecules involved in T cell migration in the thymus of mice infected with Leishmania infantum. Sci. Rep. 2017, 7, 45991. [Google Scholar] [CrossRef]
- Losada-Barragán, M.; Umaña-Pérez, A.; Durães, J.; Cuervo-Escobar, S.; Rodríguez-Vega, A.; Ribeiro-Gomes, F.L.; Berbert, L.R.; Morgado, F.; Porrozzi, R.; Mendes-da-Cruz, D.A.; et al. Thymic microenvironment is modified by malnutrition and Leishmania infantum infection. Front. Cell. Infect. Microbiol. 2019, 9, 252. [Google Scholar] [CrossRef] [PubMed]
- Langenau, D.M.; Zon, L.I. The zebrafish: A new model of T-cell and thymic development. Nat. Rev. Immunol. 2005, 5, 307–317. [Google Scholar] [CrossRef]
- Vigliano, F.A.; Losada, A.P.; Castello, M.; Bermúdez, R.; Quiroga, M.I. Morphological and immunohistochemical characterisation of the thymus in juvenile turbot (Psetta maxima, L.). Cell Tissue Res. 2011, 346, 407–416. [Google Scholar] [CrossRef]
- Padrós, F.; Crespo, S. Ontogeny of the lymphoid organs in the turbot Scophthalmus maximus: A light and electron microscope study. Aquaculture 1996, 144, 1–16. [Google Scholar] [CrossRef]
- Ronza, P.; Robledo, D.; Bermúdez, R.; Losada, A.P.; Pardo, B.G.; Martínez, P.; Quiroga, M.I. Integrating genomic and morphological approaches in fish pathology research: The case of turbot (Scophthalmus maximus) enteromyxosis. Front. Genet. 2019, 10, 26. [Google Scholar] [CrossRef]
- Bermúdez, R.; Losada, A.P.; Vázquez, S.; Redondo, M.J.; Álvarez-Pellitero, P.; Quiroga, M.I. Light and electron microscopic studies on turbot Psetta maxima infected with Enteromyxum scophthalmi: Histopathology of turbot enteromyxosis. Dis. Aquat. Organ. 2010, 89, 209–221. [Google Scholar] [CrossRef]
- Robledo, D.; Ronza, P.; Harrison, P.W.; Losada, A.P.; Bermúdez, R.; Pardo, B.G.; Redondo, M.J.; Sitjà-Bobadilla, A.; Quiroga, M.I.; Martínez, P. RNA-seq analysis reveals significant transcriptome changes in turbot (Scophthalmus maximus) suffering severe enteromyxosis. BMC Genom. 2014, 15, 1149. [Google Scholar] [CrossRef]
- Ronza, P.; Robledo, D.; Bermudez, R.; Losada, A.P.; Pardo, B.G.; Sitja-Bobadilla, A.; Quiroga, M.I.; Martinez, P. RNA-seq analysis of early enteromyxosis in turbot (Scophthalmus maximus): New insights into parasite invasion and immune evasion strategies. Int. J. Parasitol. 2016, 46, 507–517. [Google Scholar] [CrossRef]
- Boardman, T.; Warner, C.; Ramirez-Gomez, F.; Matrisciano, J.; Bromage, E. Characterization of an anti-rainbow trout (Oncorhynchus mykiss) CD3ε monoclonal antibody. Vet. Immunol. Immunop. 2012, 145, 511–515. [Google Scholar] [CrossRef]
- Losada, A.P.; Ronza, P.; Bermúdez, R.; Castrillo, A.; de Azevedo, A.M.; Quiroga, M.I. Proliferative Cell Nuclear Antigen (PCNA) Expression in the Intestine of Scophthalmus maximus Experimentally Infected with Enteromyxum scophthalmi (Myxozoa). In Proceedings of the 18th International Conference on Diseases of Fish and Shellfish, Belfast, UK, 4–8 September 2017. [Google Scholar]
- Losada, A.P.; Bermúdez, R.; Faílde, L.D.; de Ocenda, M.V.R.; Quiroga, M.I. Study of the distribution of active caspase-3-positive cells in turbot, Scophthalmus maximus (L.), enteromyxosis. J. Fish Dis. 2014, 37, 21–32. [Google Scholar] [CrossRef]
- Losada, A.P.; Bermúdez, R.; Faílde, L.D.; Quiroga, M.I. Quantitative and qualitative evaluation of iNOS expression in turbot (Psetta maxima) infected with Enteromyxum scophthalmi. Fish Shellfish Immunol. 2012, 32, 243–248. [Google Scholar] [CrossRef]
- Ronza, P.; Losada, A.P.; Villamarín, A.; Bermúdez, R.; Quiroga, M.I. Immunolocalisation of tumor necrosis factor alpha in turbot (Scophthalmus maximus, L.) tissues. Fish Shellfish Immunol. 2015, 45, 470–476. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Figueras, A.; Robledo, D.; Corvelo, A.; Hermida, M.; Pereiro, P.; Rubiolo, J.A.; Gomez-Garrido, J.; Carrete, L.; Bello, X.; Gut, M.; et al. Whole genome sequencing of turbot (Scophthalmus maximus; Pleuronectiformes): A fish adapted to demersal life. DNA Res. 2016, 23, 181–192. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2013, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef]
- Griffith, A.V.; Venables, T.; Shi, J.; Farr, A.; van Remmen, H.; Szweda, L.; Fallahi, M.; Rabinovitch, P.; Petrie, H.T. Metabolic damage and premature thymus aging caused by stromal catalase deficiency. Cell Rep. 2015, 12, 1071–1079. [Google Scholar] [CrossRef]
- Palmer, D. The effect of age on thymic function. Front. Immunol. 2013, 4, 316. [Google Scholar] [CrossRef]
- Shanley, D.P.; Aw, D.; Manley, N.R.; Palmer, D.B. An evolutionary perspective on the mechanisms of immunosenescence. Trends Immunol. 2009, 30, 374–381. [Google Scholar] [CrossRef]
- Torroba, M.; Zapata, A.G. Aging of the vertebrate immune system. Microsc. Res. Techniq. 2003, 62, 477–481. [Google Scholar] [CrossRef]
- Ansari, A.R.; Liu, H. Acute thymic involution and mechanisms for recovery. Arch. Immunol. Ther. Exp. 2017, 65, 401–420. [Google Scholar] [CrossRef]
- Ronza, P.; Bermudez, R.; Losada, A.P.; Sitja-Bobadilla, A.; Pardo, B.G.; Quiroga, M.I. Immunohistochemical detection and gene expression of TNFalpha in turbot (Scophthalmus maximus) enteromyxosis. Fish Shellfish Immunol. 2015, 47, 368–376. [Google Scholar] [CrossRef]
- Borges, M.; Barreira-Silva, P.; Flórido, M.; Jordan, M.B.; Correia-Neves, M.; Appelberg, R. Molecular and cellular mechanisms of Mycobacterium avium-induced thymic atrophy. J. Immunol. 2012, 189, 3600–3608. [Google Scholar] [CrossRef]
- Pérez, A.R.; Berbert, L.R.; Lepletier, A.; Revelli, S.; Bottasso, O.; Silva-Barbosa, S.D.; Savino, W. TNF-α is involved in the abnormal thymocyte migration during experimental Trypanosoma cruzi infection and favors the export of immature cells. PLoS ONE 2012, 7, e34360. [Google Scholar] [CrossRef]
- Lima, A.C.D.; Francelin, C.; Ferrucci, D.L.; Stach-Machado, D.R.; Verinaud, L. Thymic alterations induced by Plasmodium berghei: Expression of matrix metalloproteinases and their tissue inhibitors. Cell. Immunol. 2012, 279, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Suganami, T.; Kim-Saijo, M.; Toda, C.; Tsuiji, M.; Ochi, K.; Kamei, Y.; Minokoshi, Y.; Ogawa, Y. Role of central leptin signaling in the starvation-induced alteration of B-cell development. J. Neurosci. 2011, 31, 8373–8380. [Google Scholar] [CrossRef] [PubMed]
- Prentice, A.M. The thymus: A barometer of malnutrition. Br. J. Nutr. 1999, 81, 345–347. [Google Scholar] [CrossRef] [PubMed]
- Velloso, L.A.; Savino, W.; Mansour, E. Leptin action in the thymus. Ann. N. Y. Acad. Sci. 2009, 1153, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Procaccini, C.; La Rocca, C.; Carbone, F.; De Rosa, V.; Galgani, M.; Matarese, G. Leptin as immune mediator: Interaction between neuroendocrine and immune system. Dev. Comp. Immunol. 2017, 66, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Ceccarelli, G.; Caprera, C.; Caccia, E.; Baldassini, M.R.; Marino, G. Apoptosis in thymus of teleost fish. Fish Shellfish Immunol. 2013, 35, 589–594. [Google Scholar] [CrossRef]
- Bermúdez, R.; Vigliano, F.; Marcaccini, A.; Sitjà-Bobadilla, A.; Quiroga, M.I.; Nieto, J.M. Response of Ig-positive cells to Enteromyxum scophthalmi (Myxozoa) experimental infection in turbot, Scophthalmus maximus (L.): A histopathological and immunohistochemical study. Fish Shellfish Immunol. 2006, 21, 501–512. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Redondo, M.J.; Bermúdez, R.; Palenzuela, O.; Ferreiro, I.; Riaza, A.; Quiroga, I.; Nieto, J.M.; Alvarez-Pellitero, P. Innate and adaptive immune responses of turbot, Scophthalmus maximus (L.), following experimental infection with Enteromyxum scophthalmi (Myxosporea: Myxozoa). Fish Shellfish Immunol. 2006, 21, 485–500. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Description | Working Conditions | Reference |
|---|---|---|---|
| CD3ε (Donated by Dr. Erin Bromage) | Chain of T-cell co-receptor | 1:500 No AR required | [22] |
| PCNA (M0879, Dako) | Cofactor of DNA polymerase delta | 1:500 dilution AR: HUP in pH 6 buffer | [23] |
| Active caspase-3 (G7481, Promega) | Effector protease in apoptosis | 1:200 dilution AR: HUP in pH 8 buffer | [24] |
| Inos (RB-1605, Thermo Fisher Scientific) | Nitric oxide synthase | 1:5000 dilution No AR required | [25] |
| TNF-α (ab6671, Abcam) | Proinflammatory cytokine | 1:600 dilution AR: HUP in pH 6 buffer | [26] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronza, P.; Robledo, D.; Losada, A.P.; Bermúdez, R.; Pardo, B.G.; Martínez, P.; Quiroga, M.I. The Teleost Thymus in Health and Disease: New Insights from Transcriptomic and Histopathological Analyses of Turbot, Scophthalmus maximus. Biology 2020, 9, 221. https://doi.org/10.3390/biology9080221
Ronza P, Robledo D, Losada AP, Bermúdez R, Pardo BG, Martínez P, Quiroga MI. The Teleost Thymus in Health and Disease: New Insights from Transcriptomic and Histopathological Analyses of Turbot, Scophthalmus maximus. Biology. 2020; 9(8):221. https://doi.org/10.3390/biology9080221
Chicago/Turabian StyleRonza, Paolo, Diego Robledo, Ana Paula Losada, Roberto Bermúdez, Belén G. Pardo, Paulino Martínez, and María Isabel Quiroga. 2020. "The Teleost Thymus in Health and Disease: New Insights from Transcriptomic and Histopathological Analyses of Turbot, Scophthalmus maximus" Biology 9, no. 8: 221. https://doi.org/10.3390/biology9080221
APA StyleRonza, P., Robledo, D., Losada, A. P., Bermúdez, R., Pardo, B. G., Martínez, P., & Quiroga, M. I. (2020). The Teleost Thymus in Health and Disease: New Insights from Transcriptomic and Histopathological Analyses of Turbot, Scophthalmus maximus. Biology, 9(8), 221. https://doi.org/10.3390/biology9080221