Transcriptome Analysis Reveals That Abeliophyllum distichum Nakai Extract Inhibits RANKL-Mediated Osteoclastogenensis Mainly through Suppressing Nfatc1 Expression

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Extracts of Abeliophyllum distichum
2.2. Osteoclast Differentiation Assay
2.3. Cell Proliferation Assay
2.4. RNA-seq Data Analysis
2.5. Real-Time Quantitative PCR (qRT-PCR)
2.6. Immunoblotting
2.7. Reporter Gene Assay
2.8. Micro-Computed Tomography (Micro-CT) Analysis
2.9. Bone Histomorphometry
2.10. Zebrafish Maintenance
2.11. Whole-Mount Skeletal Staining
2.12. Statistical Analysis
2.13. Accession Numbers
3. Results
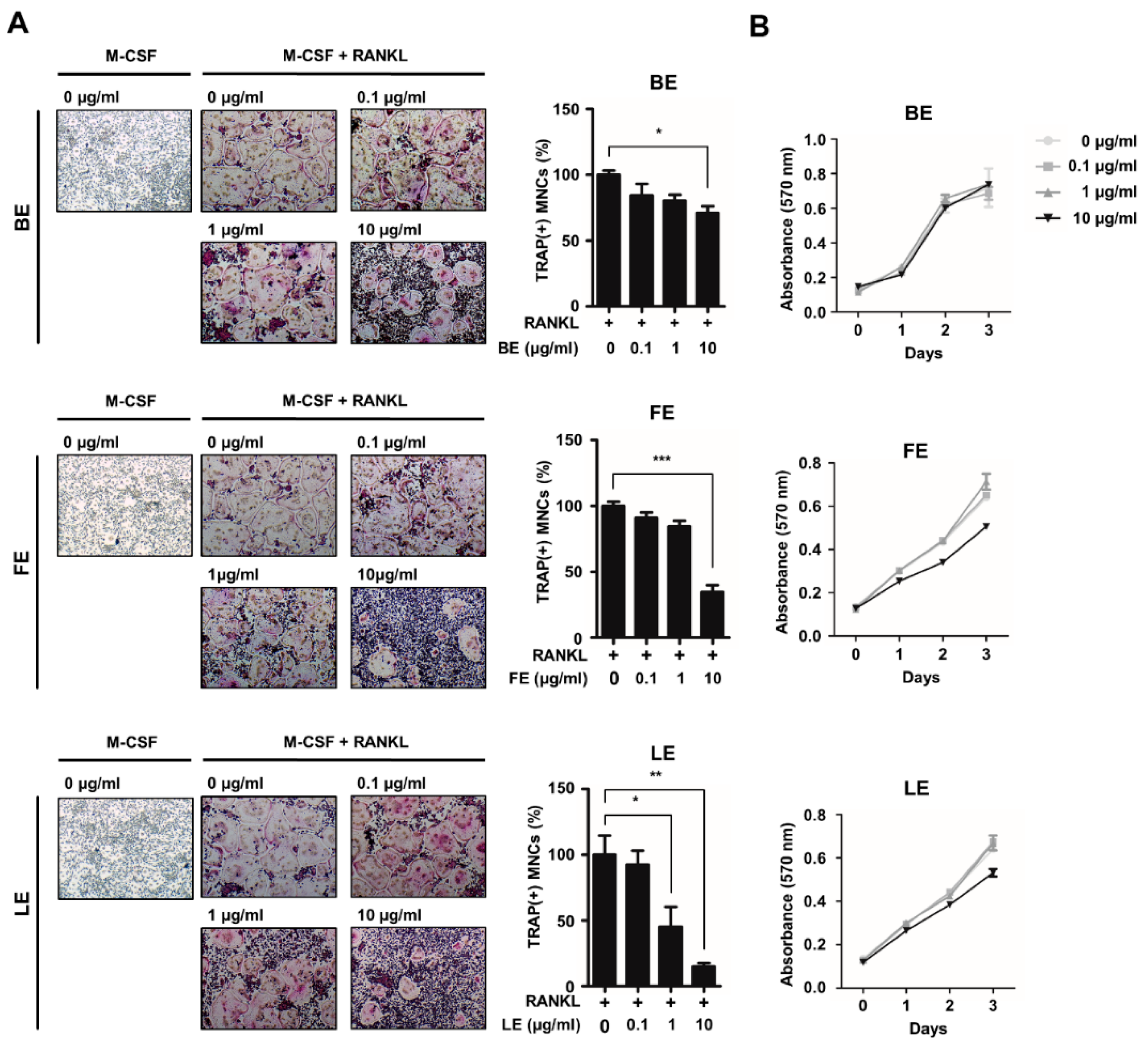
3.1. Leaf Extract of Abeliophyllum distichum Inhibits Osteoclast Differentiation
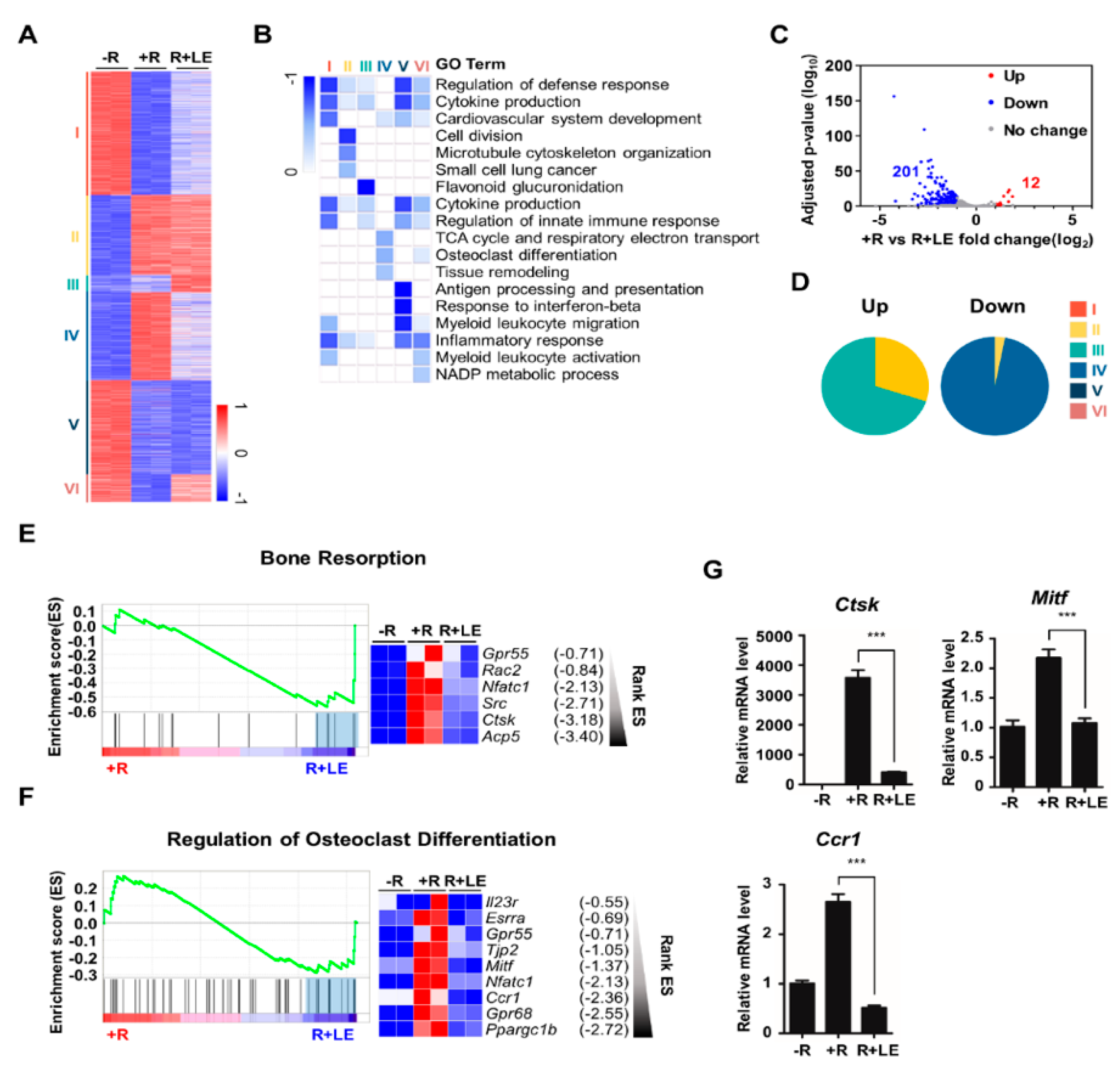
3.2. Leaf Extract of Abeliophyllum distichum Regulates the Expression of a Set of Osteoclastogenic Genes during Osteoclastogenesis
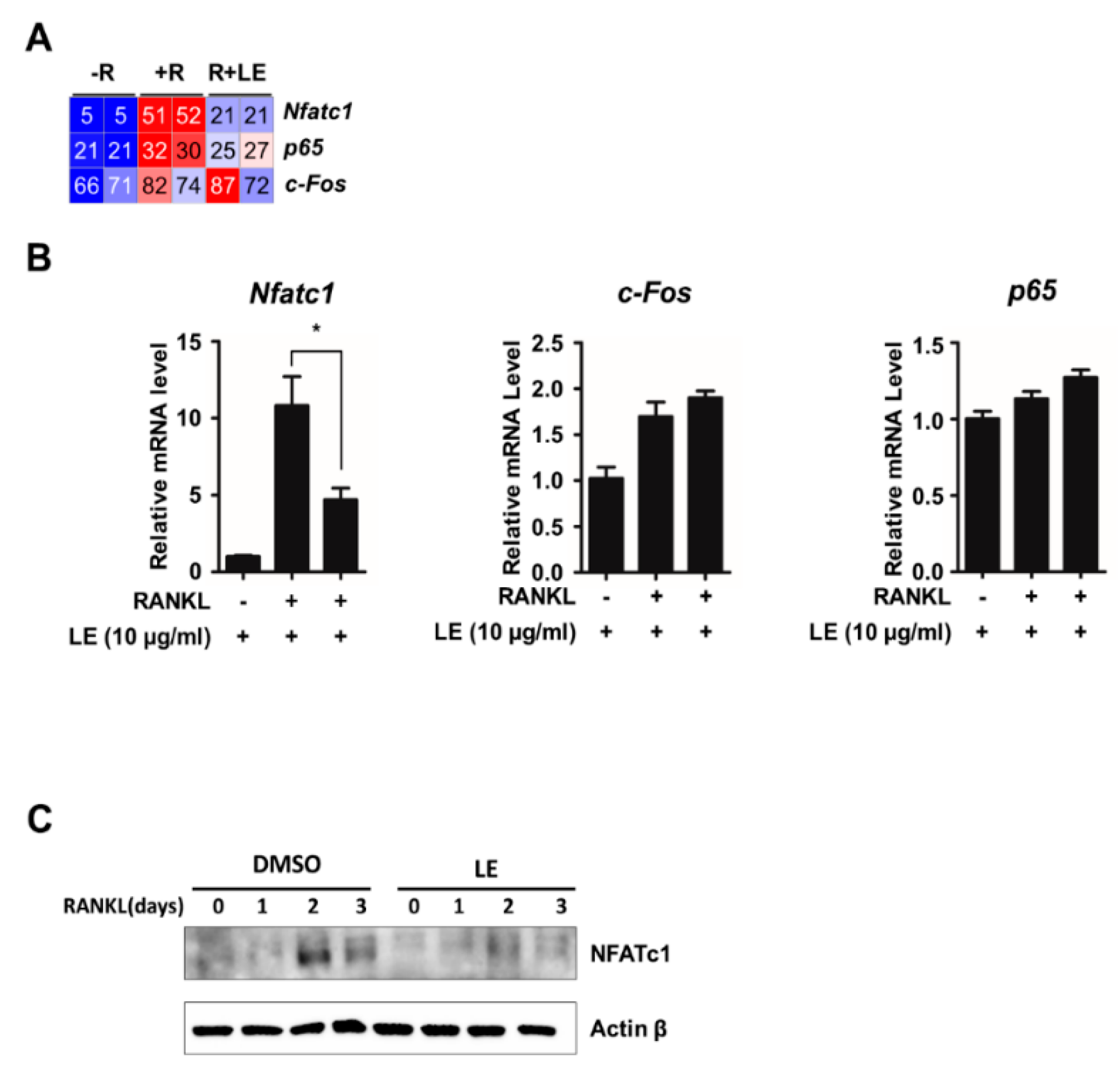
3.3. Leaf Extract of Abeliophyllum distichum Decreases RANKL-Induced NFATc1 Expression
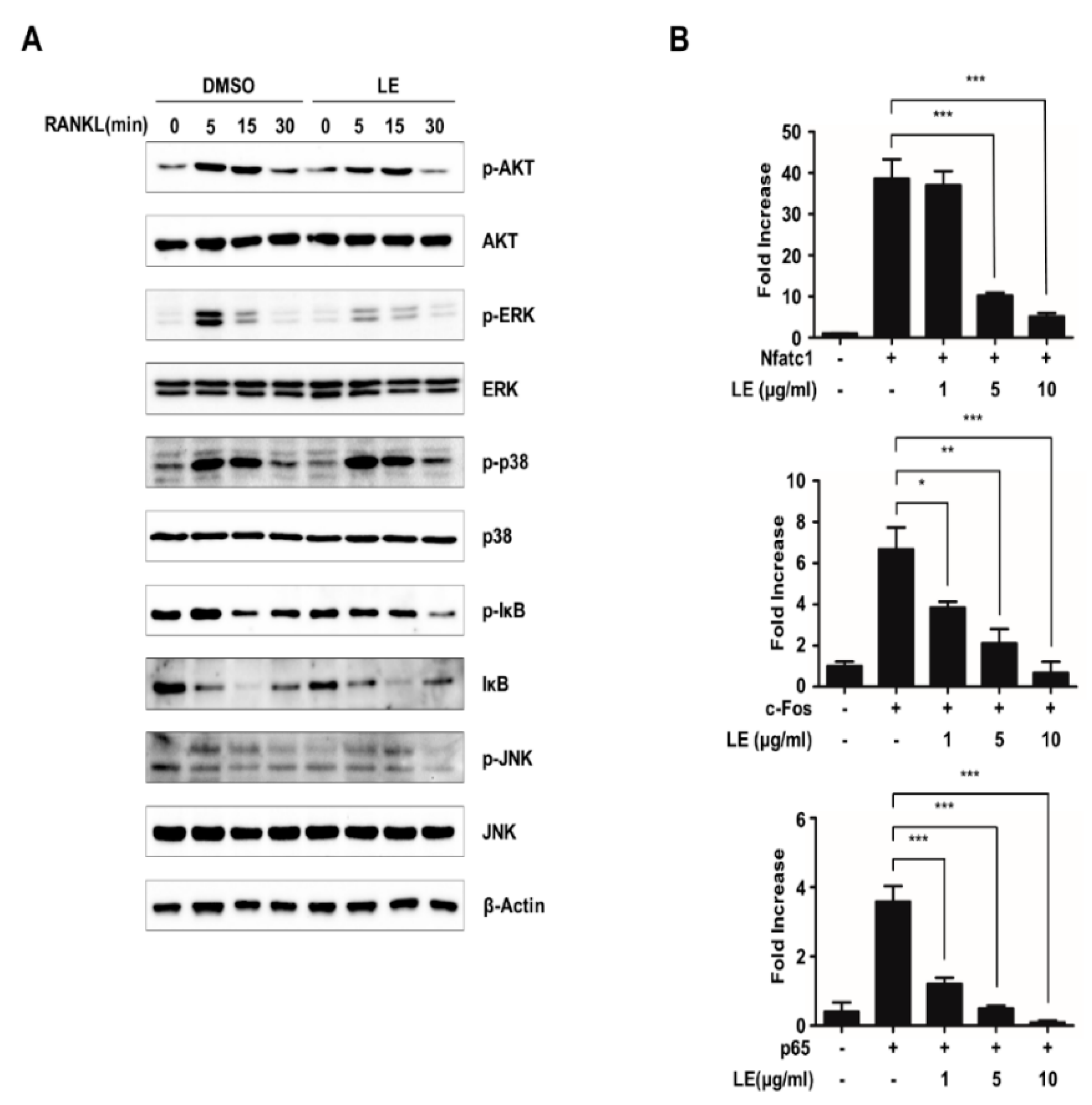
3.4. Leaf Extract of Abeliophyllum distichum Suppresses NFATc1 Expression by the Inhibition of Phosphorylation of ERK and the Inhibition of Transcriptional Activities of NFATc1, c-FOS, and p65
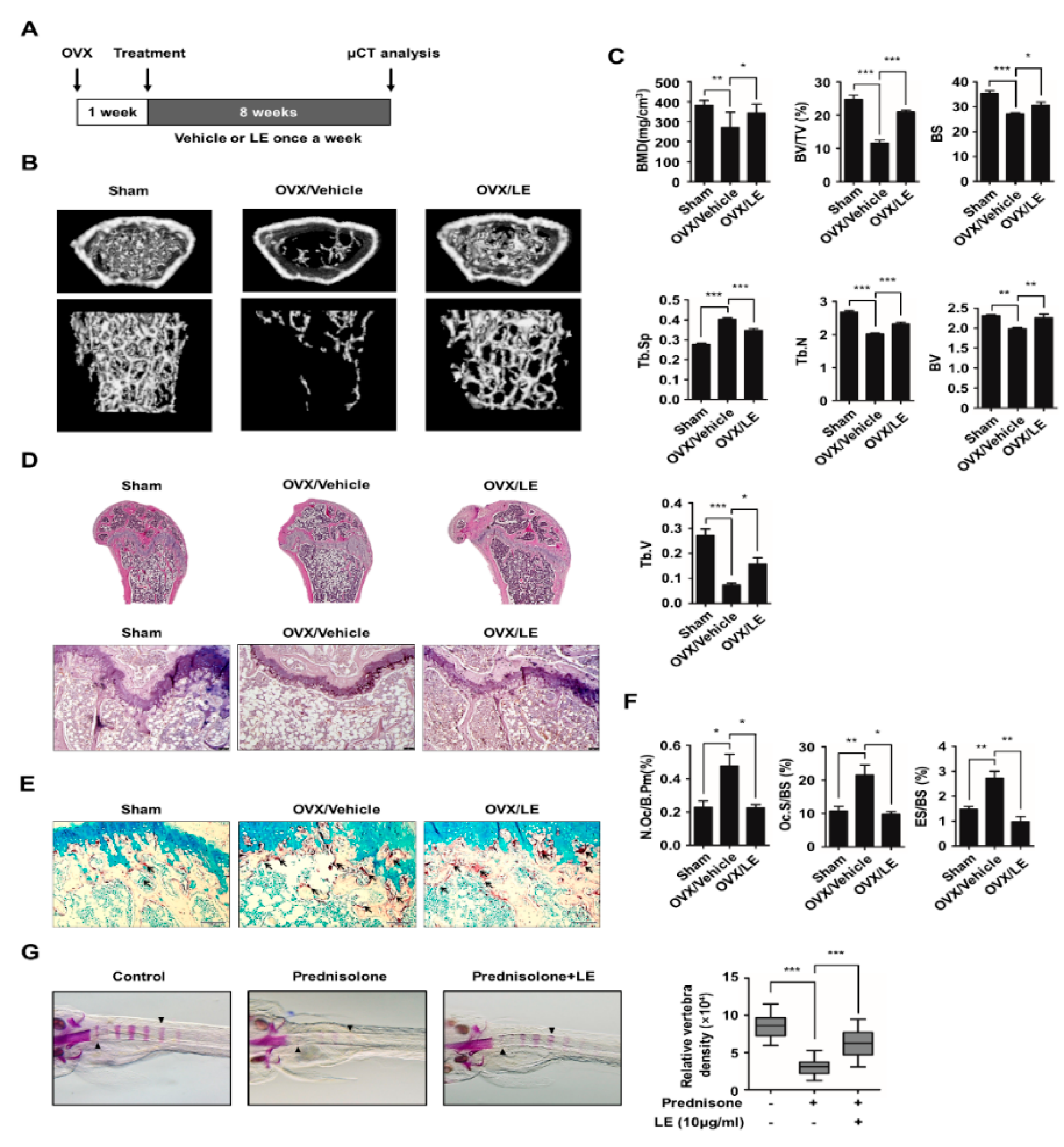
3.5. Leaf Extract of Abeliophyllum distichum Prevents Bone Loss in Both Ovariectomized Mice and Prednisolone-Treated Zebrafish
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RANKL | Receptor activator of the NF-κB ligand |
| NFATc1 | Nuclear factor of activated T-cells, cytoplasmic 1 |
| ERK | Extracellular-signal-regulated kinase |
| BMM | Bone marrow-derived macrophage |
| GIO | Glucocorticoid-induced osteoporosis |
| TRAP | Tartrate resistant acid phosphatase |
References
- Yi, S.J.; Lee, H.; Lee, J.; Lee, K.; Kim, J.; Kim, Y.; Park, J.I.; Kim, K. Bone Remodeling: Histone Modifications as Fate Determinants of Bone Cell Differentiation. Int. J. Mol. Sci. 2019, 20, 3147. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; McDonald, J.M. Disorders of bone remodeling. Annu. Rev. Pathol. 2011, 6, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Glaser, D.L.; Kaplan, F.S. Osteoporosis. Definition and clinical presentation. Spine 1997, 22, 12S–16S. [Google Scholar] [CrossRef] [PubMed]
- Compston, J.E.; McClung, M.R.; Leslie, W.D. Osteoporosis. Lancet 2019, 393, 364–376. [Google Scholar] [CrossRef]
- Cauley, J.A. Public health impact of osteoporosis. J. Gerontol. Ser. A Biol. Sci. Med Sci. 2013, 68, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Dobbs, M.B.; Buckwalter, J.; Saltzman, C. Osteoporosis: The increasing role of the orthopaedist. Iowa Orthop. J. 1999, 19, 43–52. [Google Scholar]
- Mazziotti, G.; Angeli, A.; Bilezikian, J.P.; Canalis, E.; Giustina, A. Glucocorticoid-induced osteoporosis: An update. Trends Endocrinol. Metab. 2006, 17, 144–149. [Google Scholar] [CrossRef]
- Hsu, H.; Lacey, D.L.; Dunstan, C.R.; Solovyev, I.; Colombero, A.; Timms, E.; Tan, H.L.; Elliott, G.; Kelley, M.J.; Sarosi, I.; et al. Tumor necrosis factor receptor family member RANK mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc. Natl. Acad. Sci. USA 1999, 96, 3540–3545. [Google Scholar] [CrossRef]
- Liu, C.; Walter, T.S.; Huang, P.; Zhang, S.; Zhu, X.; Wu, Y.; Wedderburn, L.R.; Tang, P.; Owens, R.J.; Stuart, D.I.; et al. Structural and functional insights of RANKL-RANK interaction and signaling. J. Immunol. 2010, 184, 6910–6919. [Google Scholar] [CrossRef]
- Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; Yasuda, H.; Yano, K.; Morinaga, T.; Higashio, K. RANK is the essential signaling receptor for osteoclast differentiation factor in osteoclastogenesis. Biochem. Biophys. Res. Commun. 1998, 253, 395–400. [Google Scholar] [CrossRef]
- Lee, N.K.; Choi, Y.G.; Baik, J.Y.; Han, S.Y.; Jeong, D.W.; Bae, Y.S.; Kim, N.; Lee, S.Y. A crucial role for reactive oxygen species in RANKL-induced osteoclast differentiation. Blood 2005, 106, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, N. Regulation of NFATc1 in Osteoclast Differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Yamashita, T.; Yao, Z.Q.; Zhang, Q.; Li, F.; Xing, L.P. Roles for NF-kappa B and c-Fos in osteoclasts. J. Bone Miner. Metab. 2005, 23, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Seo, E.-J.; Sung, J.; Choi, K.M.; Kim, H.; Kim, J.-S.; Lee, J.; Efferth, T.; Hyun, T.K. Polyphenolic compounds, antioxidant and anti-inflammatory effects of Abeliophyllum distichum Nakai extract. J. Appl. Bot. Food Qual. 2017, 90. [Google Scholar] [CrossRef]
- Li, H.M.; Kim, J.K.; Jang, J.M.; Cui, C.B.; Lim, S.S. Analysis of the inhibitory activity of Abeliophyllum distichum leaf constituents against aldose reductase by using high-speed counter current chromatography. Arch. Pharmacal. Res. 2013, 36, 1104–1112. [Google Scholar] [CrossRef]
- Oh, H.; Kang, D.G.; Kwon, T.O.; Jang, K.K.; Chai, K.Y.; Yun, Y.G.; Chung, H.T.; Lee, H.S. Four glycosides from the leaves of Abeliophyllum distichum with inhibitory effects on angiotensin converting enzyme. Phytother. Res. 2003, 17, 811–813. [Google Scholar] [CrossRef]
- Park, G.H.; Park, J.H.; Eo, H.J.; Song, H.M.; Woo, S.H.; Kim, M.K.; Lee, J.W.; Lee, M.H.; Lee, J.R.; Koo, J.S.; et al. The induction of activating transcription factor 3 (ATF3) contributes to anti-cancer activity of Abeliophyllum distichum Nakai in human colorectal cancer cells. BMC Complement. Altern. Med. 2014, 14, 487. [Google Scholar] [CrossRef]
- Lee, H.; Lee, K.; Lee, S.; Lee, J.; Jeong, W.T.; Lim, H.B.; Hyun, T.K.; Yi, S.J.; Kim, K. Ethyl Acetate Fraction of Aqueous Extract of Lentinula edodes Inhibits Osteoclastogenesis by Suppressing NFATc1 Expression. Int. J. Med. Sci. 2020, 17, 21–32. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- An, D.; Kim, K.; Lu, W. Defective entry into mitosis 1 (Dim1) negatively regulates osteoclastogenesis by inhibiting the expression of nuclear factor of activated T-cells, cytoplasmic, calcineurin-dependent 1 (NFATc1). J. Biol. Chem. 2014, 289, 24366–24373. [Google Scholar] [CrossRef] [PubMed]
- Dempster, D.W.; Compston, J.E.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R.; Parfitt, A.M. Standardized nomenclature, symbols, and units for bone histomorphometry: A 2012 update of the report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone Miner. Res. 2013, 28, 2–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, J.; Lee, H.; Shin, W.R.; Lee, S.; Lee, J.; Park, J.I.; Jhun, B.H.; Kim, Y.H.; Yi, S.J.; et al. Tetracycline Analogs Inhibit Osteoclast Differentiation by Suppressing MMP-9-Mediated Histone H3 Cleavage. Int. J. Mol. Sci. 2019, 20, 4038. [Google Scholar] [CrossRef] [PubMed]
- Abu-Amer, Y. NF-kappaB signaling and bone resorption. Osteoporos. Int. 2013, 24, 2377–2386. [Google Scholar] [CrossRef]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Lee, K.; Seo, I.; Choi, M.H.; Jeong, D. Roles of Mitogen-Activated Protein Kinases in Osteoclast Biology. Int. J. Mol. Sci. 2018, 19, 3004. [Google Scholar] [CrossRef]
- Monje, P.; Hernandez-Losa, J.; Lyons, R.J.; Castellone, M.D.; Gutkind, J.S. Regulation of the transcriptional activity of c-Fos by ERK. A novel role for the prolyl isomerase PIN1. J. Biol. Chem. 2005, 280, 35081–35084. [Google Scholar] [CrossRef]
- Kim, K.; Kim, J.H.; Lee, J.; Jin, H.M.; Kook, H.; Kim, K.K.; Lee, S.Y.; Kim, N. MafB negatively regulates RANKL-mediated osteoclast differentiation. Blood 2007, 109, 3253–3259. [Google Scholar] [CrossRef]
- Zhao, B.; Ivashkiv, L.B. Negative regulation of osteoclastogenesis and bone resorption by cytokines and transcriptional repressors. Arthritis Res. Ther. 2011, 13, 234. [Google Scholar] [CrossRef]
- Barrett, R.; Chappell, C.; Quick, M.; Fleming, A. A rapid, high content, in vivo model of glucocorticoid-induced osteoporosis. Biotechnol. J. 2006, 1, 651–655. [Google Scholar] [CrossRef]
- He, H.; Wang, C.; Tang, Q.; Yang, F.; Xu, Y. Possible mechanisms of prednisolone-induced osteoporosis in zebrafish larva. Biomed. Pharmacother. 2018, 101, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Compston, J.; Cooper, A.; Cooper, C.; Gittoes, N.; Gregson, C.; Harvey, N.; Hope, S.; Kanis, J.A.; McCloskey, E.V.; Poole, K.E.S.; et al. UK clinical guideline for the prevention and treatment of osteoporosis. Arch. Osteoporos. 2017, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Cheon, Y.H.; Baek, J.M.; Park, S.H.; Ahn, S.J.; Lee, M.S.; Oh, J.; Kim, J.Y. Stauntonia hexaphylla (Lardizabalaceae) leaf methanol extract inhibits osteoclastogenesis and bone resorption activity via proteasome-mediated degradation of c-Fos protein and suppression of NFATc1 expression. BMC Complement. Altern. Med. 2015, 15, 280. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.H.; Jang, S.A.; Kim, T.; Ha, H. Forsythia suspensa Protects against Bone Loss in Ovariectomized Mice. Nutrients 2019, 11, 1831. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, V.; Sarkar, M.; Chaubey, P.; Khan, T.; Sherje, A.; Patel, K.; Dravyakar, B. Bone Health and Natural Products—An Insight. Front. Pharmacol. 2018, 9, 981. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Kim, J.H.; Kim, M.; Park, J.H.; Sohn, Y.; Jung, H.S. Abeliophyllum distichum Nakai alleviates postmenopausal osteoporosis in ovariectomized rats and prevents RANKL-induced osteoclastogenesis In Vitro. J. Ethnopharmacol. 2020, 257, 112828. [Google Scholar] [CrossRef] [PubMed]
- Yoo, T.K.; Kim, J.S.; Hyun, T.K. Polyphenolic Composition and Anti-Melanoma Activity of White Forsythia (Abeliophyllum distichum Nakai) Organ Extracts. Plants 2020, 9, 757. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, K.S.; Yi, S.H.; Kook, S.H.; Lee, J.C. Acteoside suppresses RANKL-mediated osteoclastogenesis by inhibiting c-Fos induction and NF-kappaB pathway and attenuating ROS production. PLoS ONE 2013, 8, e80873. [Google Scholar] [CrossRef]
- Ang, E.S.; Pavlos, N.J.; Chai, L.Y.; Qi, M.; Cheng, T.S.; Steer, J.H.; Joyce, D.A.; Zheng, M.H.; Xu, J. Caffeic acid phenethyl ester, an active component of honeybee propolis attenuates osteoclastogenesis and bone resorption via the suppression of RANKL-induced NF-kappaB and NFAT activity. J. Cell. Physiol. 2009, 221, 642–649. [Google Scholar] [CrossRef]
- Kwak, S.C.; Lee, C.; Kim, J.Y.; Oh, H.M.; So, H.S.; Lee, M.S.; Rho, M.C.; Oh, J. Chlorogenic acid inhibits osteoclast differentiation and bone resorption by down-regulation of receptor activator of nuclear factor kappa-B ligand-induced nuclear factor of activated T cells c1 expression. Biol. Pharm. Bull. 2013, 36, 1779–1786. [Google Scholar] [CrossRef]
- Doss, H.M.; Samarpita, S.; Ganesan, R.; Rasool, M. Ferulic acid, a dietary polyphenol suppresses osteoclast differentiation and bone erosion via the inhibition of RANKL dependent NF-kappaB signalling pathway. Life Sci. 2018, 207, 284–295. [Google Scholar] [CrossRef] [PubMed]
- La, V.D.; Tanabe, S.; Grenier, D. Naringenin inhibits human osteoclastogenesis and osteoclastic bone resorption. J. Periodontal Res. 2009, 44, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Masuhara, M.; Tsukahara, T.; Tomita, K.; Furukawa, M.; Miyawaki, S.; Sato, T. A relation between osteoclastogenesis inhibition and membrane-type estrogen receptor GPR30. Biochem. Biophys. Rep. 2016, 8, 389–394. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kyung, T.W.; Lee, J.E.; Shin, H.H.; Choi, H.S. Rutin inhibits osteoclast formation by decreasing reactive oxygen species and TNF-alpha by inhibiting activation of NF-kappaB. Exp. Mol. Med. 2008, 40, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.A.; Adachi, J.D. Glucocorticoid-induced osteoporosis: Treatment update and review. Ther. Adv. Musculoskelet. Dis. 2009, 1, 71–85. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.; Jang, Y.-J.; Lee, H.; Kim, E.; Kim, Y.; Yoo, T.-K.; Hyun, T.K.; Park, J.-I.; Yi, S.-J.; Kim, K. Transcriptome Analysis Reveals That Abeliophyllum distichum Nakai Extract Inhibits RANKL-Mediated Osteoclastogenensis Mainly through Suppressing Nfatc1 Expression. Biology 2020, 9, 212. https://doi.org/10.3390/biology9080212
Lee K, Jang Y-J, Lee H, Kim E, Kim Y, Yoo T-K, Hyun TK, Park J-I, Yi S-J, Kim K. Transcriptome Analysis Reveals That Abeliophyllum distichum Nakai Extract Inhibits RANKL-Mediated Osteoclastogenensis Mainly through Suppressing Nfatc1 Expression. Biology. 2020; 9(8):212. https://doi.org/10.3390/biology9080212
Chicago/Turabian StyleLee, Kyubin, You-Jee Jang, Hyerim Lee, Eunbin Kim, Yeojin Kim, Tong-Kewn Yoo, Tae Kyung Hyun, Jae-Il Park, Sun-Ju Yi, and Kyunghwan Kim. 2020. "Transcriptome Analysis Reveals That Abeliophyllum distichum Nakai Extract Inhibits RANKL-Mediated Osteoclastogenensis Mainly through Suppressing Nfatc1 Expression" Biology 9, no. 8: 212. https://doi.org/10.3390/biology9080212
APA StyleLee, K., Jang, Y.-J., Lee, H., Kim, E., Kim, Y., Yoo, T.-K., Hyun, T. K., Park, J.-I., Yi, S.-J., & Kim, K. (2020). Transcriptome Analysis Reveals That Abeliophyllum distichum Nakai Extract Inhibits RANKL-Mediated Osteoclastogenensis Mainly through Suppressing Nfatc1 Expression. Biology, 9(8), 212. https://doi.org/10.3390/biology9080212