Human Cytomegalovirus Infection Induces High Expression of Prolactin and Prolactin Receptors in Ovarian Cancer

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Increased PRL Transcript Levels in HCMV-Infected SKOV3 Cells
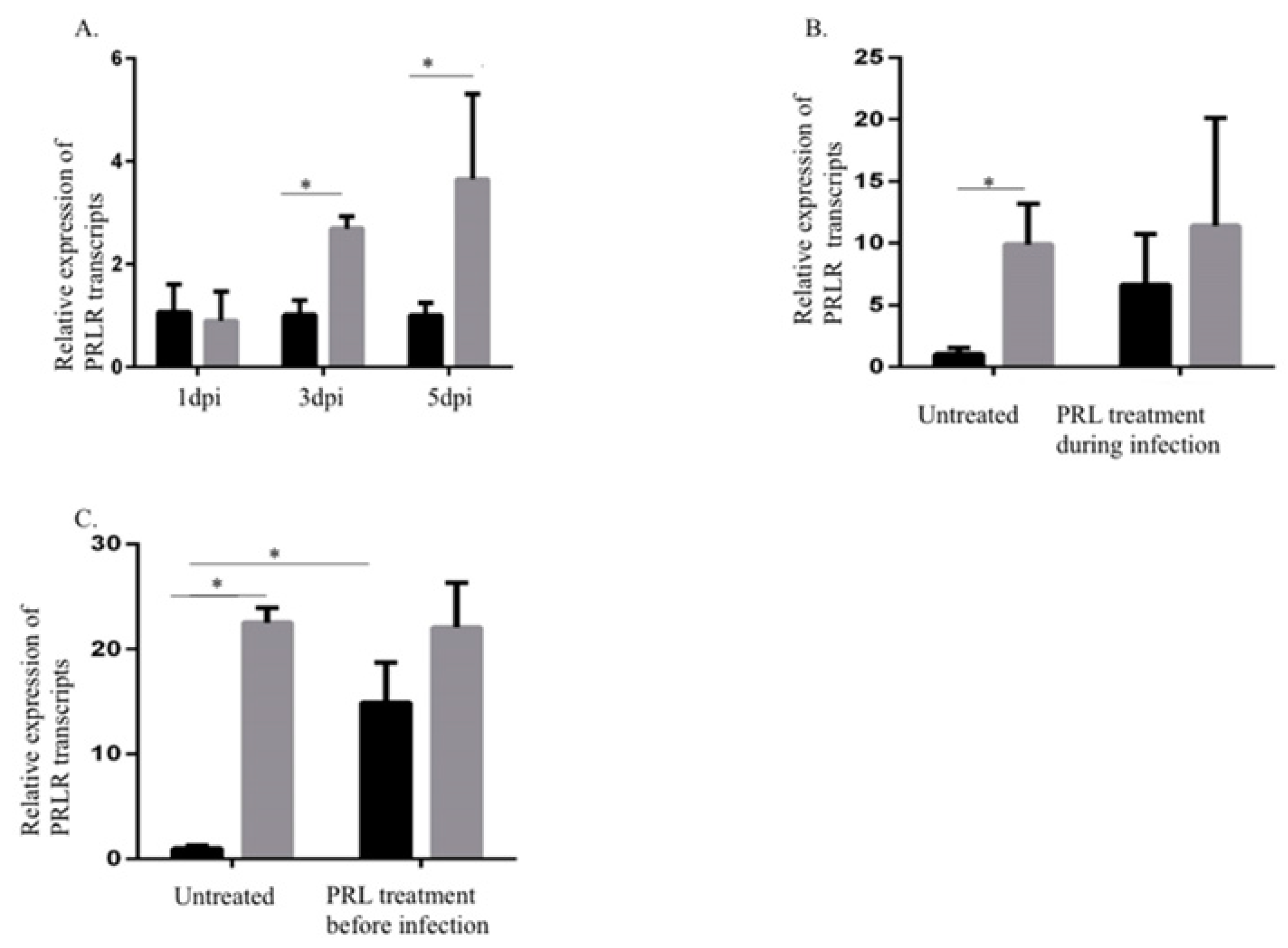
2.2. Increased PRLR Transcript Levels in HCMV-Infected SKOV3 Cells
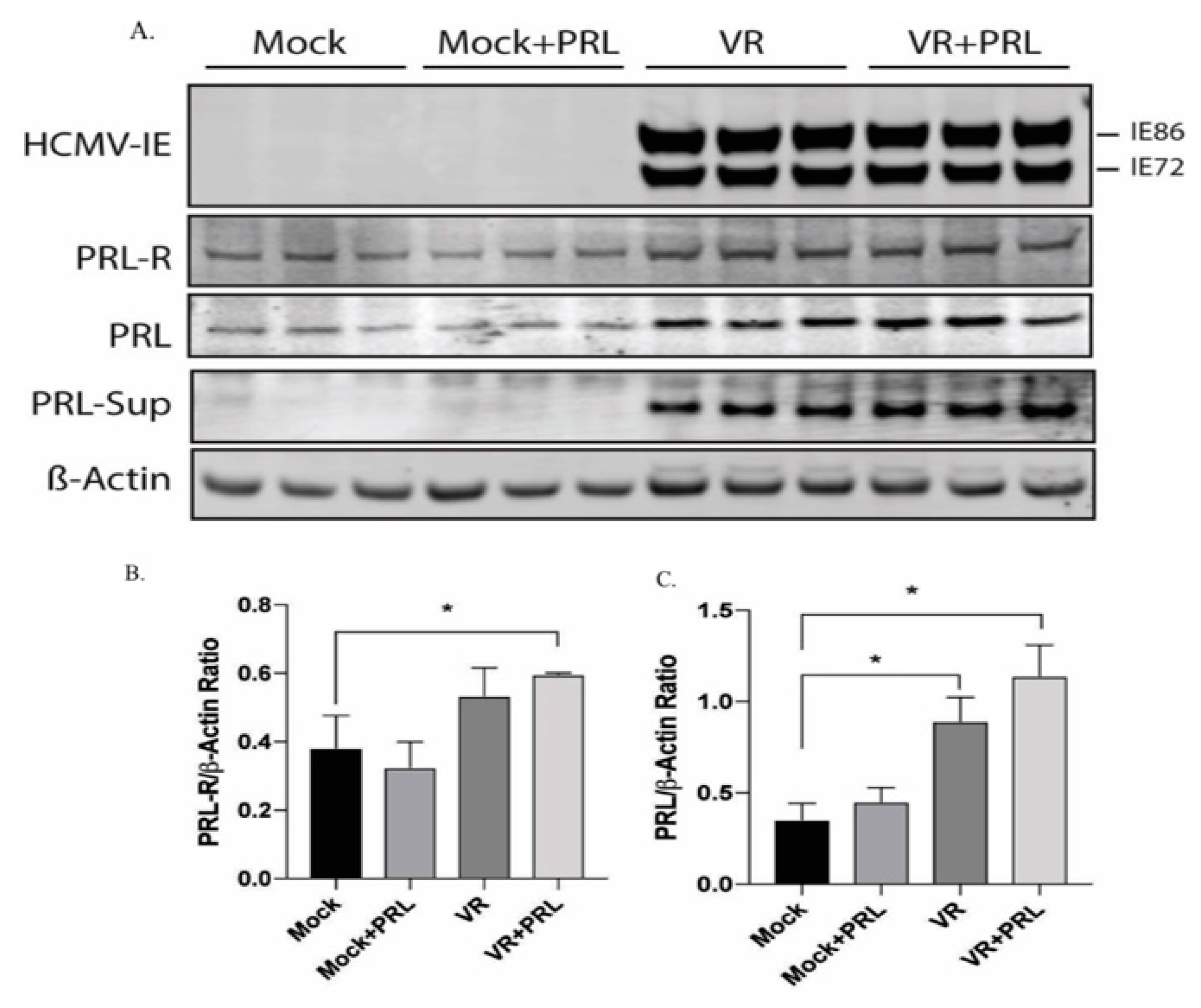
2.3. HCMV Induces PRL and PRLR Protein Expression in HCMV-Infected SKOV3 Cells
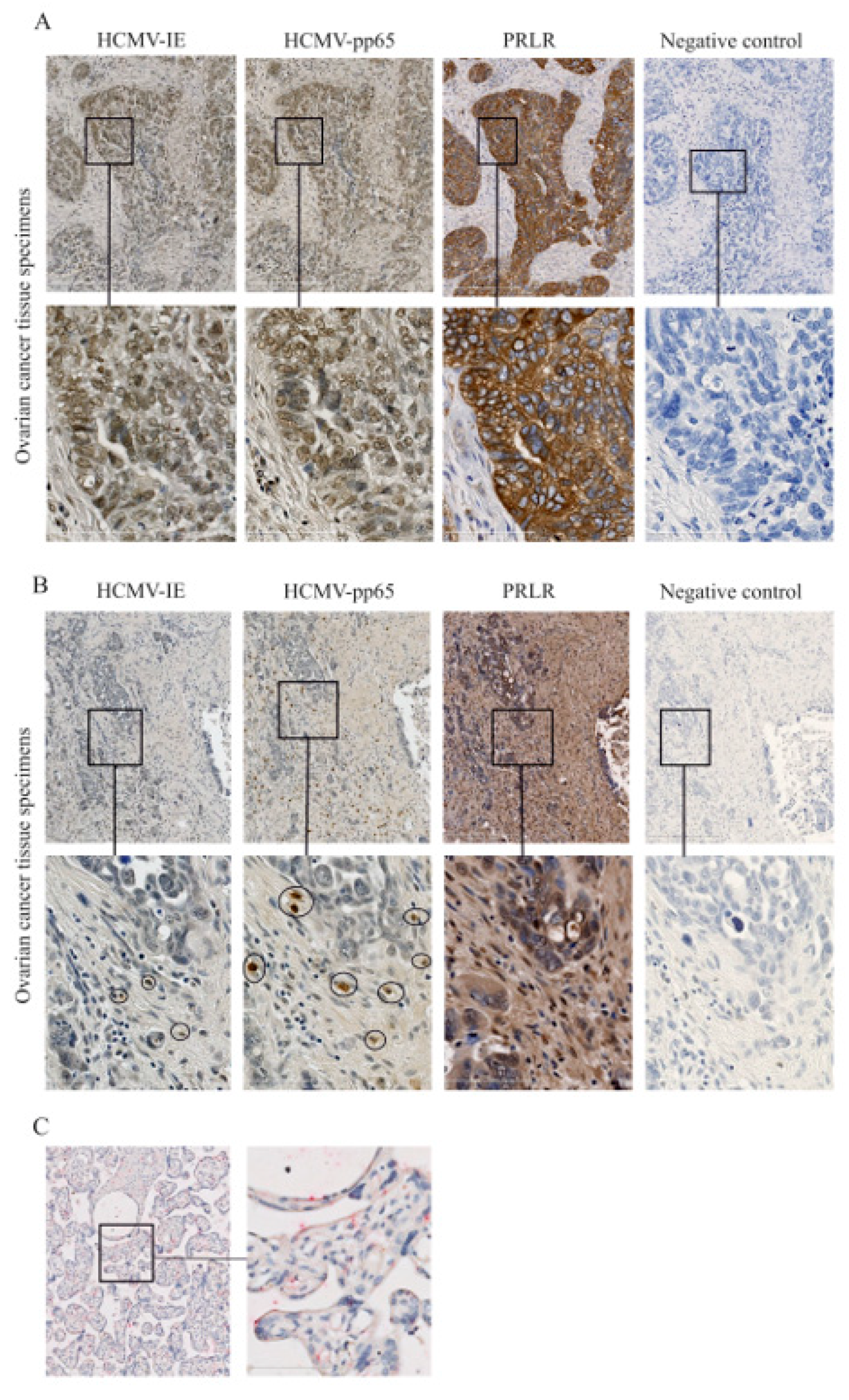
2.4. Expression of PRLR in HCMV-Infected Ovarian Cancer Tissues
3. Discussion
4. Materials and Methods
4.1. Cell Culture, HCMV Infection, and PRL Treatment
4.2. Western Blotting
4.3. RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.4. Immunohistochemical Staining
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Shin, H.-R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008, GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Calò, V.; Bruno, L.; Rizzo, S.; Bazan, V.; Di Fede, G. Hereditary ovarian cancer. Crit. Rev. Oncol. Hematol. 2009, 69, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://ocrahope.org/patients/about-ovarian-cancer/risk-factors/ (accessed on 5 November 2019).
- Brown, S.B.; Hankinson, S.E. Endogenous estrogens and the risk of breast, endometrial, and ovarian cancers. Steroids 2015, 99, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Taher, C.; de Boniface, J.; Mohammad, A.A.; Religa, P.; Hartman, J.; Yaiw, K.C.; Frisell, J.; Rahbar, A.; Söderberg-Naucler, C. High Prevalence of Human Cytomegalovirus Proteins and Nucleic Acids in Primary Breast Cancer and Metastatic Sentinel Lymph Nodes. PLoS ONE 2013, 8, e56795. [Google Scholar] [CrossRef] [PubMed]
- Schuessler, A.; Walker, D.G.; Khanna, R. Cytomegalovirus as a novel target for immunotherapy of glioblastoma multiforme. Front. Oncol. 2014, 4, 275. [Google Scholar] [CrossRef] [Green Version]
- Harkins, L.E.; Matlaf, L.A.; Soroceanu, L.; Klemm, K.; Britt, W.J.; Wang, W.; Bland, K.I.; Cobbs, C.S. Detection of human cytomegalovirus in normal and neoplastic breast epithelium. Herpesviridae 2010, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Cobbs, C.S.; Harkins, L.; Samanta, M.; Gillespie, G.Y.; Bharara, S.; King, P.H.; Nabors, L.B.; Cobbs, C.G.; Britt, W.J. Human Cytomegalovirus Infection and Expression in Human Malignant Glioma. Cancer Res. 2002, 62, 3347–3350. [Google Scholar]
- Tafvizi, F.; Fard, Z.T. Detection of human cytomegalovirus in patients with colorectal cancer by nested-PCR. Asian Pac. J. Cancer Prev. 2014, 15, 1453–1457. [Google Scholar] [CrossRef] [Green Version]
- Samanta, M.; Harkins, L.; Klemm, K.; Britt, W.J.; Cobbs, C.S. High prevalence of human cytomegalovirus in prostatic intraepithelial neoplasia and prostatic carcinoma. J. Urol. 2003, 170, 998–1002. [Google Scholar] [CrossRef]
- Lucas, K.G.; Bao, L.; Bruggeman, R.; Dunham, K.; Specht, C. The detection of CMV pp65 and IE1 in glioblastoma multiforme. J. Neuro-Oncol. 2011, 103, 231–238. [Google Scholar] [CrossRef]
- Baryawno, N.; Rahbar, A.; Wolmer-Solberg, N.; Taher, C.; Odeberg, J.; Darabi, A.; Khan, Z.; Sveinbjörnsson, B.; FuskevÅg, O.M.; Segerström, L.; et al. Detection of human cytomegalovirus in medulloblastomas reveals a potential therapeutic target. J. Clin. Investig. 2011, 121, 4043–4055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolmer-Solberg, N.; Baryawno, N.; Rahbar, A.; Fuchs, D.; Odeberg, J.; Taher, C.; Wilhelmi, V.; Milosevic, J.; Mohammad, A.A.; Martinsson, T.; et al. Frequent detection of human cytomegalovirus in neuroblastoma: A novel therapeutic target? Int. J. Cancer 2013, 133, 2351–2361. [Google Scholar] [CrossRef] [PubMed]
- Hahn, G.; Jores, R.; Mocarski, E.S. Cytomegalovirus remains latent in a common precursor of dendritic and myeloid cells. Proc. Natl. Acad. Sci. USA 1998, 95, 3937–3942. [Google Scholar] [CrossRef] [Green Version]
- Arvin, A.; Campadelli-Fiume, G.; Mocarski, E.; Moore, P.S.; Roizman, B.; Whitley, R.; Yamanishi, K. (Eds.) Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar] [CrossRef]
- Wille, P.T.; Wisner, T.W.; Ryckman, B.; Johnson, D.C. Human cytomegalovirus (HCMV) glycoprotein gB promotes virus entry in Trans acting as the viral fusion protein rather than as a receptor-binding protein. MBio 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söderberg-Nauclér, C.; Fish, K.N.; Nelson, J.A. Reactivation of Latent Human Cytomegalovirus by Allogeneic Stimulation of Blood Cells from Healthy Donors. Cell 1997, 91, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Mason, G.M.; Poole, E.; Sissons, J.G.P.; Wills, M.R.; Sinclair, J.H. Human cytomegalovirus latency alters the cellular secretome, inducing cluster of differentiation (CD)4+ T-cell migration and suppression of effector function. Proc. Natl. Acad. Sci. USA 2012, 109, 14538–14543. [Google Scholar] [CrossRef] [Green Version]
- Bodaghi, B.; Goureau, O.; Zipeto, D.; Laurent, L.; Virelizier, J.-L.; Michelson, S. Role of IFN-γ-Induced Indoleamine 2,3 Dioxygenase and Inducible Nitric Oxide Synthase in the Replication of Human Cytomegalovirus in Retinal Pigment Epithelial Cells. J. Immunol. 1999, 162, 957–964. [Google Scholar]
- Kotenko, S.V.; Saccani, S.; Izotova, L.S.; Mirochnitchenko, O.V.; Pestka, S. Human cytomegalovirus harbors its own unique IL-10 homolog (cmvIL-10). Proc. Natl. Acad. Sci. USA 2000, 97, 1695–1700. [Google Scholar] [CrossRef] [Green Version]
- Reeves, M.B.; Compton, T. Inhibition of inflammatory interleukin-6 activity via extracellular signal-regulated kinase-mitogen-activated protein kinase signaling antagonizes human cytomegalovirus reactivation from dendritic cells. J. Virol. 2011, 85, 12750–12758. [Google Scholar] [CrossRef] [Green Version]
- Benard, M.; Straat, K.; Omarsdottir, S.; Leghmari, K.; Bertrand, J.; Davrinche, C.; Duga-Neulat, I.; Söderberg-Nauclér, C.; Rahbar, A.; Casper, C. Human cytomegalovirus infection induces leukotriene B4 and 5-lipoxygenase expression in human placentae and umbilical vein endothelial cells. Placenta 2014, 35, 345–350. [Google Scholar] [CrossRef]
- Qiu, H.; Strååt, K.; Rahbar, A.; Wan, M.; Söderberg-Nauclér, C.; Haeggström, J.Z. Human CMV infection induces 5-lipoxygenase expression and leukotriene B4 production in vascular smooth muscle cells. J. Exp. Med. 2008, 205, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooks, J.J.; Chin, M.S.; Srinivasan, K.; Momma, Y.; Hooper, L.C.; Nagineni, C.N.; Chan, C.C.; Detrick, B. Human cytomegalovirus induced cyclooxygenase-2 in human retinal pigment epithelial cells augments viral replication through a prostaglandin pathway. Microbes Infect. 2006, 8, 2236–2244. [Google Scholar] [CrossRef] [PubMed]
- Antman, E.M.; DeMets, D.; Loscalzo, J. Cyclooxygenase Inhibition and Cardiovascular Risk. Circulation 2005, 112, 759–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.E.; Redeker, A.; Arens, R.; van Baarle, D.; van Den Berg, S.P.; Benedict, C.A.; Čičin-Šain, L.; Hill, A.B.; Wills, M.R. CMV immune evasion and manipulation of the immune system with aging. GeroScience 2017, 273–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, C.; DeFilippis, V.; Malouli, D.; Früh, K. Cytomegalovirus immune evasion. Curr. Top. Microbiol. Immunol. 2008, 333–359. [Google Scholar] [CrossRef]
- Abdullah, A.A.; Abdullah, R.; Nazariah, Z.A.; Balakrishnan, K.N.; Abdullah, F.F.J.; Bala, J.A.; Mohd-Lila, M.A. Cyclophilin a as a target in the treatment of cytomegalovirus infections. Antivir. Chem. Chemother. 2018. [Google Scholar] [CrossRef]
- Syed, F.; Rycyzyn, M.A.; Westgate, L.; Clevenger, C.V. A novel and functional interaction between cyclophilin A and prolactin receptor. Endocrine 2003, 20, 83–89. [Google Scholar] [CrossRef]
- Moussawi, F.A.; Kumar, A.; Pasquereau, S.; Tripathy, M.K.; Karam, W.; Diab-Assaf, M.; Herbein, G. The transcriptome of human mammary epithelial cells infected with the HCMV-DB strain displays oncogenic traits. Sci. Rep. 2018, 8, 12574. [Google Scholar] [CrossRef]
- Kumar, A.; Tripathy, M.K.; Pasquereau, S.; Al Moussawi, F.; Abbas, W.; Coquard, L.; Khan, K.A.; Russo, L.; Algros, M.P.; Valmary-Degano, S.; et al. The Human Cytomegalovirus Strain DB Activates Oncogenic Pathways in Mammary Epithelial Cells. EBioMedicine 2018, 30, 167–183. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, A.; Touma, J.; Costa, H.; Davoudi, B.; Bukholm, I.R.; Sauer, T.; Vetvik, K.; Geisler, J.; Söderberg-Naucler, C. Low Expression of Estrogen Receptor-α and Progesterone Receptor in Human Breast Cancer Tissues Is Associated with High-Grade Human Cytomegalovirus Protein Expression. Clin. Breast Cancer 2017, 17, 526–535.e1. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, T.; Rapp, F. Malignant transformation of hamster embryo fibroblasts following exposure to ultraviolet-irradiated human cytomegalovirus. Virology 1973, 55, 53–61. [Google Scholar] [CrossRef]
- Clendenen, T.V.; Arslan, A.A.; Lokshin, A.E.; Liu, M.; Lundin, E.; Koenig, K.L.; Berrino, F.; Hallmans, G.; Idahl, A.; Krogh, V.; et al. Circulating prolactin levels and risk of epithelial ovarian cancer. Cancer Causes Control 2013, 24, 741–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffion, A.; Al-Sakkaf, K.A.; Brown, B.L.; Eaton, C.L.; Hamdy, F.C.; Dobson, P.R.M. The Survival Effect of Prolactin on PC3 Prostate Cancer Cells. European Urology. Elsevier 2003, 43, 301–308. [Google Scholar] [CrossRef]
- Perks, C.M.; Keith, A.J.; Goodhew, K.L.; Savage, P.B.; Winters, Z.E.; Holly, J.M.P. Prolactin acts as a potent survival factor for human breast cancer cell lines. British journal of cancer. Nat. Publ. Group 2004, 91, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Ben-Jonathan, N.; Liby, K.; McFarland, M.; Zinger, M. Prolactin as an autocrine/paracrine growth factor in human cancer. Trends Endocrinol. Metab. 2016, 13, 245–250. [Google Scholar] [CrossRef]
- Levina, V.V.; Nolen, B.; Su, Y.; Godwin, A.K.; Fishman, D.; Liu, J.; Mor, G.; Maxwell, L.G.; Herberman, R.B.; Szczepanski, M.J.; et al. Biological Significance of Prolactin in Gynecological Cancers. Cancer Res. 2009, 69, 5226–5233. [Google Scholar] [CrossRef] [Green Version]
- Alkharusi, A.; Yu, S.; Landázuri, N.; Zadjali, F.; Davodi, B.; Nyström, T.; Gräslund, T.; Rahbar, A.; Norstedt, G. Stimulation of prolactin receptor induces STAT-5 phosphorylation and cellular invasion in glioblastoma multiforme. Oncotarget 2016, 7, 79572. [Google Scholar] [CrossRef]
- Goffin, V.; Binart, N.; Touraine, P.; Kelly, P.A. Prolactin: The new biology of an old hormone. Ann. Rev. Physiol. 2002, 64, 47–67. [Google Scholar] [CrossRef]
- Berwaer, M.; Martial, J.A.; Davis, J.R. Characterization of an up-stream promoter directing extrapituitary expression of the human prolactin gene. Mol. Endocrinol. 1994, 8, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Goffin, V.; Kelly, P.A. The prolactin/growth hormone receptor family: Structure/function relationships. J. Mammary Gland Biol. Neoplasia 1997, 2, 7–17. [Google Scholar] [CrossRef]
- Alkharusi, A.; Lesma, E.; Ancona, S.; Chiaramonte, E.; Nyström, T.; Gorio, A.; Norstedt, G. Role of Prolactin Receptors in Lymphangioleiomyomatosis. PLoS ONE 2016, 11, e0146653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Arav-Boger, R. New cell-signaling pathways for controlling cytomegalovirus replication. Am. J. Transplant. 2014, 14, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Hagemeier, C.; Walker, S.; Caswell, R.; Kouzarides, T.; Sinclair, J. The human cytomegalovirus 80-kilodalton but not the 72-kilodalton immediate-early protein transactivates heterologous promoters in a TATA box- dependent mechanism and interacts directly with TFIID. J. Virol. 1992, 66, 4452–4456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, J.W.; Rådestad, A.F.; Söderberg-Naucler, C.; Rahbar, A. Human cytomegalovirus in high grade serous ovarian cancer possible implications for patients survival. Medicine 2018, 97, e9685. [Google Scholar] [CrossRef] [PubMed]
- Rådestad, A.F.; Estekizadeh, A.; Cui, H.L.; Kostopoulou, O.N.; Davoudi, B.; Hirschberg, A.L.; Carlson, J.; Rahbar, A.; Söderberg-Naucler, C. Impact of Human Cytomegalovirus Infection and its Immune Response on Survival of Patients with Ovarian Cancer. Transl. Oncol. 2018, 11, 1292–1300. [Google Scholar] [CrossRef]
- Estekizadeh, A.; Landázuri, N.; Pantalone, M.R.; Davoudi, B.; Hu, L.F.; Nawaz, I.; Stragliotto, G.; Ekström, T.J.; Rahbar, A. 5-Azacytidine treatment results in nuclear exclusion of DNA methyltransferase-1, as well as reduced proliferation and invasion in human cytomegalovirus-infected glioblastoma cells. Oncol. Rep. 2019, 41, 2927–2936. [Google Scholar] [CrossRef]
- Liu, X.; Cohen, J.I. The role of PI3K/Akt in human herpesvirus infection: From the bench to the bedside. Virology 2015, 479–480, 568–577. [Google Scholar] [CrossRef] [Green Version]
- Brisken, C.; Ayyannan, A.; Nguyen, C.; Heineman, A.; Reinhardt, F.; Jan, T.; Dey, S.K.; Dotto, G.P.; Weinberg, R.A. IGF-2 Is a Mediator of Prolactin-Induced Morphogenesis in the Breast. Devel. Cell 2002, 3, 877–887. [Google Scholar] [CrossRef] [Green Version]
- Clevenger, C.V.; Kline, J.B. Prolactin receptor signal transduction. Lupus 2001, 10, 706–718. [Google Scholar] [CrossRef]
- Plotnikov, A.; Li, Y.; Tran, T.H.; Tang, W.; Palazzo, J.P.; Rui, H.; Fuchs, S.Y. Oncogene-Mediated Inhibition of Glycogen Synthase Kinase 3β Impairs Degradation of Prolactin Receptor. Cancer Res. 2008, 68, 1354–1361. [Google Scholar] [CrossRef] [Green Version]
- Solana, R.; Tarazona, R.; Aiello, A.E.; Akbar, A.N.; Appay, V.; Beswick, M.; Bosch, J.A.; Campos, C.; Cantisán, S.; Cicin-Sain, L. CMV and Immunosenescence: From basics to clinics. Immun. Ageing 2012, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R.; Janeway, C. Innate Immunity. N. Engl. J. Med. 2000, 343, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Raspollini, M.R.; Amunni, G.; Villanucci, A.; Boddi, V.; Baroni, G.; Taddei, A.; Taddei, G.L. Expression of inducible nitric oxide synthase and cyclooxygenase-2 in ovarian cancer: Correlation with clinical outcome. Gynecol. Oncol. 2004, 92, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, X.; Sun, D.; Jia, X.; Xu, L.; Qiao, Y.; Jin, Y. COX-2 expression in ovarian cancer: An updated meta-analysis. Oncotarget 2017, 8, 88152–88162. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Comaills, V.; Burr, R.; Boulay, G.; Miyamoto, D.T.; Wittner, B.S.; Emmons, E.; Sil, S.; Koulopoulos, M.W.; Broderick, K.T.; et al. COX-2 mediates tumor-stromal prolactin signaling to initiate tumorigenesis. Proc. Natl. Acad. Sci. 2019, 116, 5223–5232. [Google Scholar] [CrossRef] [Green Version]
- Bonneterre, J.; Mauriac, L.; Weber, B.; Roche, H.; Fargeot, P.; Tubiana-Hulin, M.; Sevin, M.; Chollet, P.; Cappelaere, P. Tamoxifen plus bromocriptine versus tamoxifen plus placebo in advanced breast cancer: Results of a double blind multicentre clinical trial. Eur. J. Cancer Clin. Oncol. 1988, 24, 1851–1853. [Google Scholar] [CrossRef]
- Kelly, M.P.; Hickey, C.; Makonnen, S.; Coetzee, S.; Jalal, S.; Wang, Y.; Delfino, F.; Shan, J.; Potocky, T.B.; Chatterjee, I.; et al. Preclinical Activity of the Novel Anti-Prolactin Receptor (PRLR) Antibody–Drug Conjugate REGN2878-DM1 in PRLR-Positive Breast Cancers. Mol. Cancer Ther. 2017, 16, 1299–1311. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Alkharusi, A.; Norstedt, G.; Gräslund, T. An in vivo half-life extended prolactin receptor antagonist can prevent STAT5 phosphorylation. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Stegmann, C.; Hochdorfer, D.; Lieber, D.; Subramanian, N.; Stoehr, D.; Sampaio, K.L.; Sinzger, C. A derivative of platelet-derived growth factor receptor alpha binds to the trimer of human cytomegalovirus and inhibits entry into fibroblasts and endothelial cells. PLoS Pathog. 2017, 13. [Google Scholar] [CrossRef]
- Kim, J.H.; Collins-McMillen, D.; Buehler, J.C.; Goodrum, F.D.; Yurochko, A.D. Human Cytomegalovirus Requires Epidermal Growth Factor Receptor Signaling To Enter and Initiate the Early Steps in the Establishment of Latency in CD34 + Human Progenitor Cells. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Huong, S.M.; Chiu, M.L.; Raab-Traub, N.; Huang, E.S. Epidermal growth factor receptor is a cellular receptor for human cytomegalovirus. Nature 2003, 424, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Collins-McMillen, D.; Stevenson, E.V.; Kim, J.H.; Lee, B.J.; Cieply, S.J.; Nogalski, M.T.; Chan, G.C.; Frost, R.W.; Spohn, C.R.; Yurochko, A.D. Human Cytomegalovirus Utilizes a Nontraditional Signal Transducer and Activator of Transcription 1 Activation Cascade via Signaling through Epidermal Growth Factor Receptor and Integrins to Efficiently Promote the Motility, Differentiation, and Polarizati. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, G.; Nogalski, M.T.; Bentz, G.L.; Smith, M.S.; Parmater, A.; Yurochko, A.D. PI3K-Dependent Upregulation of Mcl-1 by Human Cytomegalovirus Is Mediated by Epidermal Growth Factor Receptor and Inhibits Apoptosis in Short-Lived Monocytes. J. Immunol. 2010, 184, 3213–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaiw, K.C.; Mohammad, A.A.; Costa, H.; Taher, C.; Badrnya, S.; Assinger, A.; Wilhelmi, V.; Ananthaseshan, S.; Estekizadeh, A.; Davoudi, B.; et al. Human cytomegalovirus up-regulates endothelin receptor type B: Implication for vasculopathies? Open Forum Infectious Diseases. 2015, 2, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teicher, B.A.; Doroshow, J.H. The Promise of Antibody–Drug Conjugates. N. Engl. J. Med. 2012, 367, 1847–1848. [Google Scholar] [CrossRef] [Green Version]
- Söderberg-Nauclér, C.; Rahbar, A.; Stragliotto, G. Survival in patients with glioblastoma receiving valganciclovir. N. Engl. J. Med. 2013, 985–986. [Google Scholar] [CrossRef] [Green Version]
- Stragliotto, G.; Rahbar, A.; Solberg, N.W.; Lilja, A.; Taher, C.; Orrego, A.; Bjurman, B.; Tammik, C.; Skarman, P.; Peredo, I.; et al. Effects of valganciclovir as an add-on therapy in patients with cytomegalovirus-positive glioblastoma: A randomized, double-blind, hypothesis-generating study. Int. J. Cancer 2013, 133, 1204–1213. [Google Scholar] [CrossRef] [Green Version]
- Rahbar, A.; Orrego, A.; Peredo, I.; Dzabic, M.; Wolmer-Solberg, N.; Strååt, K.; Stragliotto, G.; Söderberg-Nauclér, C. Human cytomegalovirus infection levels in glioblastoma multiforme are of prognostic value for survival. J. Clin. Virol. 2016, 57, 36–42. [Google Scholar] [CrossRef]
- Batich, K.A.; Reap, E.A.; Archer, G.E.; Sanchez-Perez, L.; Nair, S.K.; Schmittling, R.J.; Norberg, P.; Xie, W.; Herndon, J.E.; Healy, P.; et al. Long-term survival in glioblastoma with cytomegalovirus pp65-targeted vaccination. Clin. Cancer Res. 2017, 23, 1898–1909. [Google Scholar] [CrossRef] [Green Version]
- Estekizadeh, A.; Landázur, N.; Bartek, J.; Beltoft Brøchner, C.; Davoudi, B.; Broholm, H.; Karimi, M.; Ekström, T.J.; Rahbar, A. Increased cytomegalovirus replication by 5-Azacytidine and viral-induced cytoplasmic expression of DNMT-1 in medulloblastoma and endothelial cells. Int. J. Oncol. 2018, 52, 1317–1327. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | Ovarian Cancer | HCMV-IE | HCMV-pp65 | PRLR | OS (m) |
|---|---|---|---|---|---|
| 1 | Other histotypes | 0 | 0 | 1 | 68.5 |
| 2 | Serous adenocarcinoma | 4 | 3 | 4 | 40 |
| 3 | Serous adenocarcinoma | 0 | 1 | 3 | 42.5 |
| 4 | Other histotypes | 3 | 2 | 4 | 67 |
| 5 | Other histotypes | 4 | 4 | 4 | 41 |
| 6 | Other histotypes | 1 | 1 | 4 | 66 |
| 7 | Serous adenocarcinoma | 4 | 2 | 3 | 48 |
| 8 | Serous adenocarcinoma | 2 | 1 | 3 | 28 |
| 9 | Serous adenocarcinoma | 4 | 4 | 4 | 31 |
| 10 | Serous adenocarcinoma | 1 | 2 | 4 | 32 |
| Patient Characteristics | |
|---|---|
| Ovarian Cancer (n = 10) | |
| Median age, years (range) | 65 (42–74) |
| Stage | |
| IA, IC | n = 3 |
| IIIC | n = 6 |
| IV | n = 1 |
| Median BMI (range) | 26.5 (20–26.9) |
| Median CA125 level | |
| Initial | 436 U/mL |
| After treatment | 17.5 U/mL |
| Surgery | |
| R0 | n = 6 (60%) |
| R1 | n = 4 (40%) |
| Neoadjuvant chemotherapy before surgery | |
| Yes | n = 3 (30%) |
| No | n = 7 (70%) |
| Adjuvant chemotherapy after surgery | |
| Yes | n = 9 (90%) |
| No | n = 1 (10%) |
| Dead at study closure | n = 5 (50%) |
| Alive at study closure | n = 5 (50%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahbar, A.; AlKharusi, A.; Costa, H.; Pantalone, M.R.; Kostopoulou, O.N.; Cui, H.L.; Carlsson, J.; Rådestad, A.F.; Söderberg-Naucler, C.; Norstedt, G. Human Cytomegalovirus Infection Induces High Expression of Prolactin and Prolactin Receptors in Ovarian Cancer. Biology 2020, 9, 44. https://doi.org/10.3390/biology9030044
Rahbar A, AlKharusi A, Costa H, Pantalone MR, Kostopoulou ON, Cui HL, Carlsson J, Rådestad AF, Söderberg-Naucler C, Norstedt G. Human Cytomegalovirus Infection Induces High Expression of Prolactin and Prolactin Receptors in Ovarian Cancer. Biology. 2020; 9(3):44. https://doi.org/10.3390/biology9030044
Chicago/Turabian StyleRahbar, Afsar, Amira AlKharusi, Helena Costa, Mattia Russel Pantalone, Ourania N. Kostopoulou, Huanhuan L. Cui, Joseph Carlsson, Angelique Flöter Rådestad, Cecilia Söderberg-Naucler, and Gunnar Norstedt. 2020. "Human Cytomegalovirus Infection Induces High Expression of Prolactin and Prolactin Receptors in Ovarian Cancer" Biology 9, no. 3: 44. https://doi.org/10.3390/biology9030044