
Characterisation of Arctic Bacterial Communities in the Air above Svalbard

Abstract
:1. Introduction
2. Materials and Methods
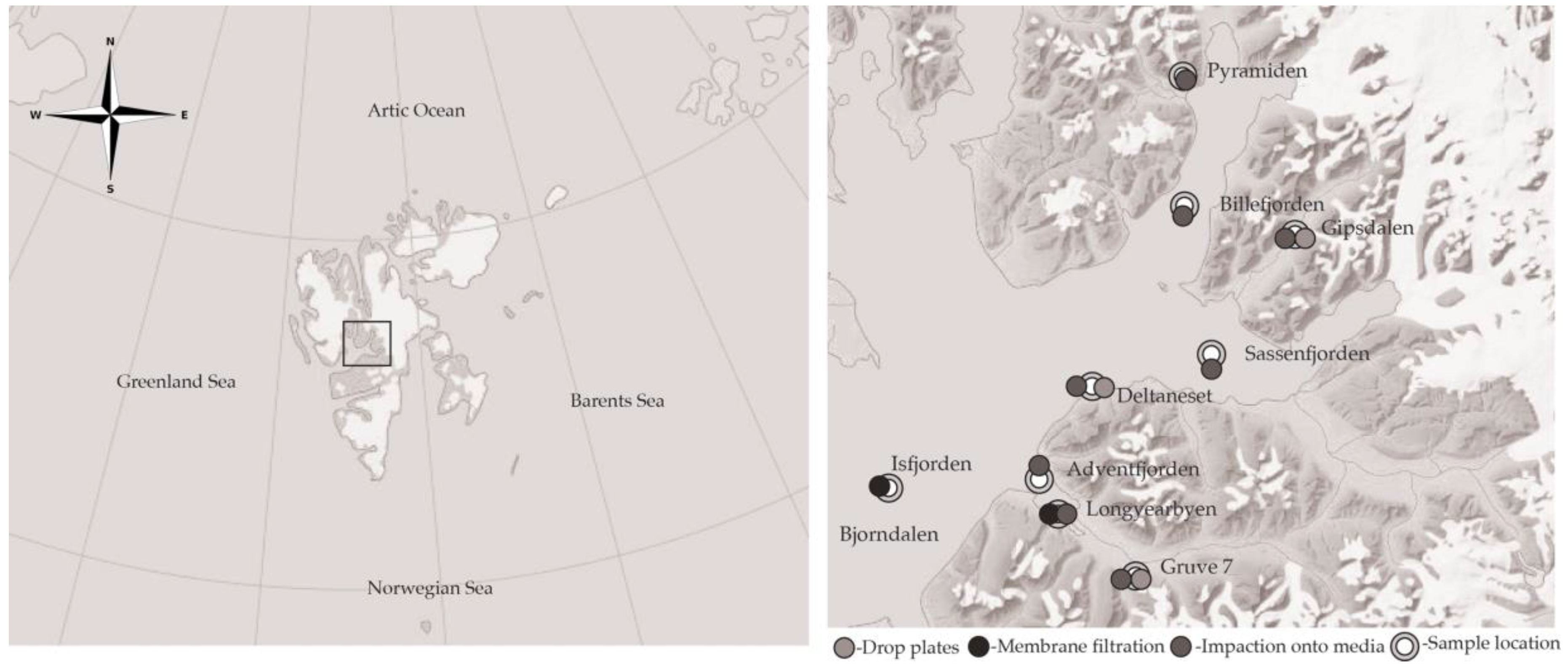
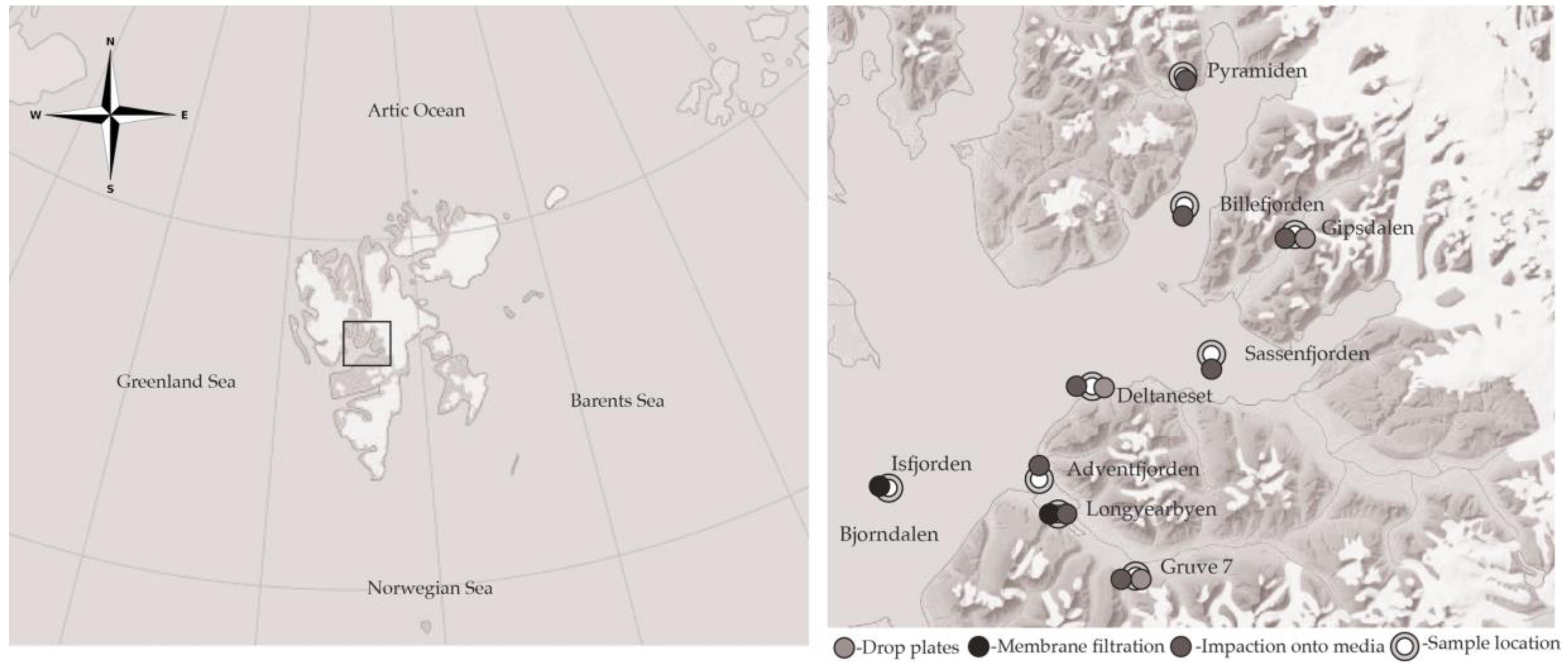
2.1. Site Description
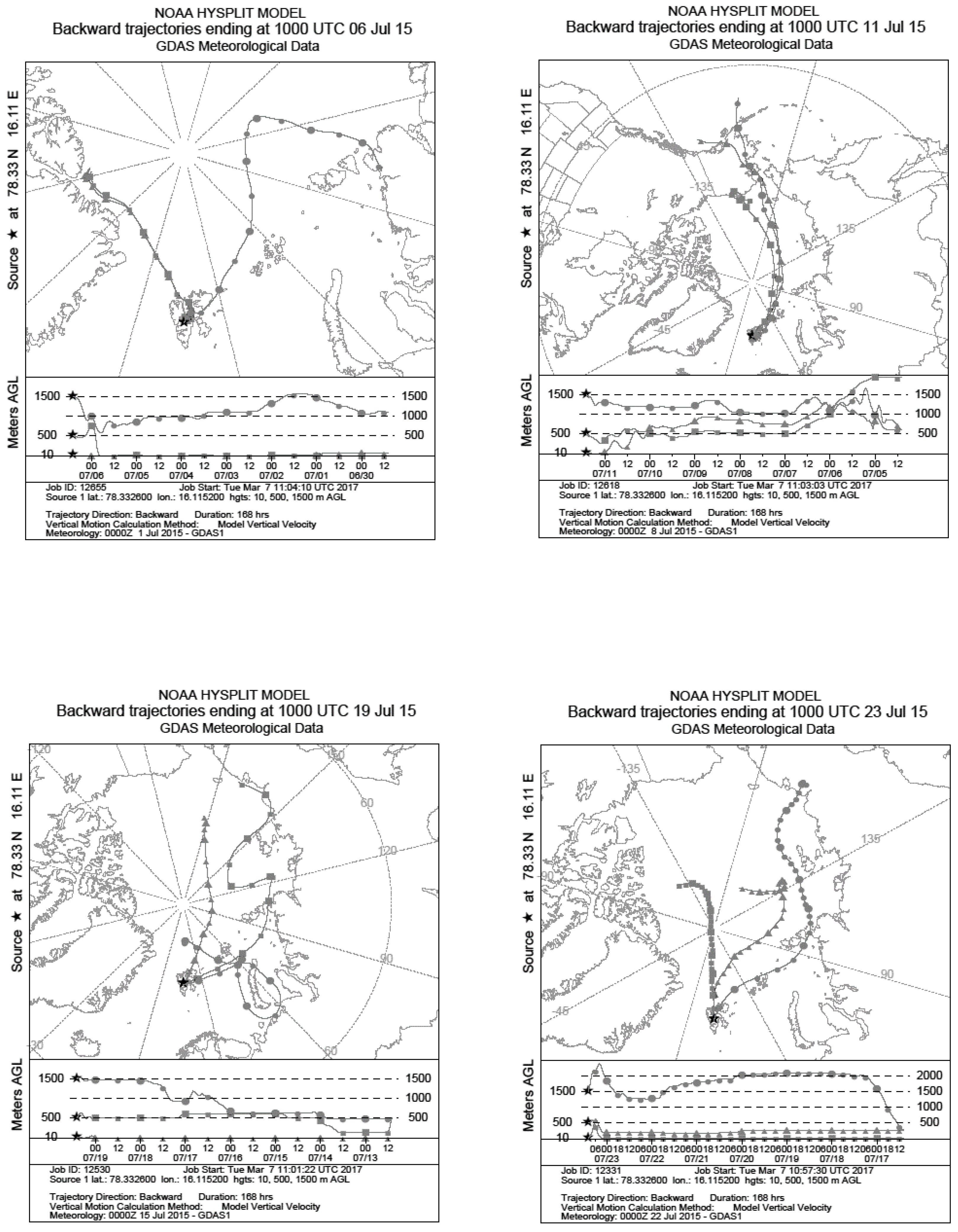
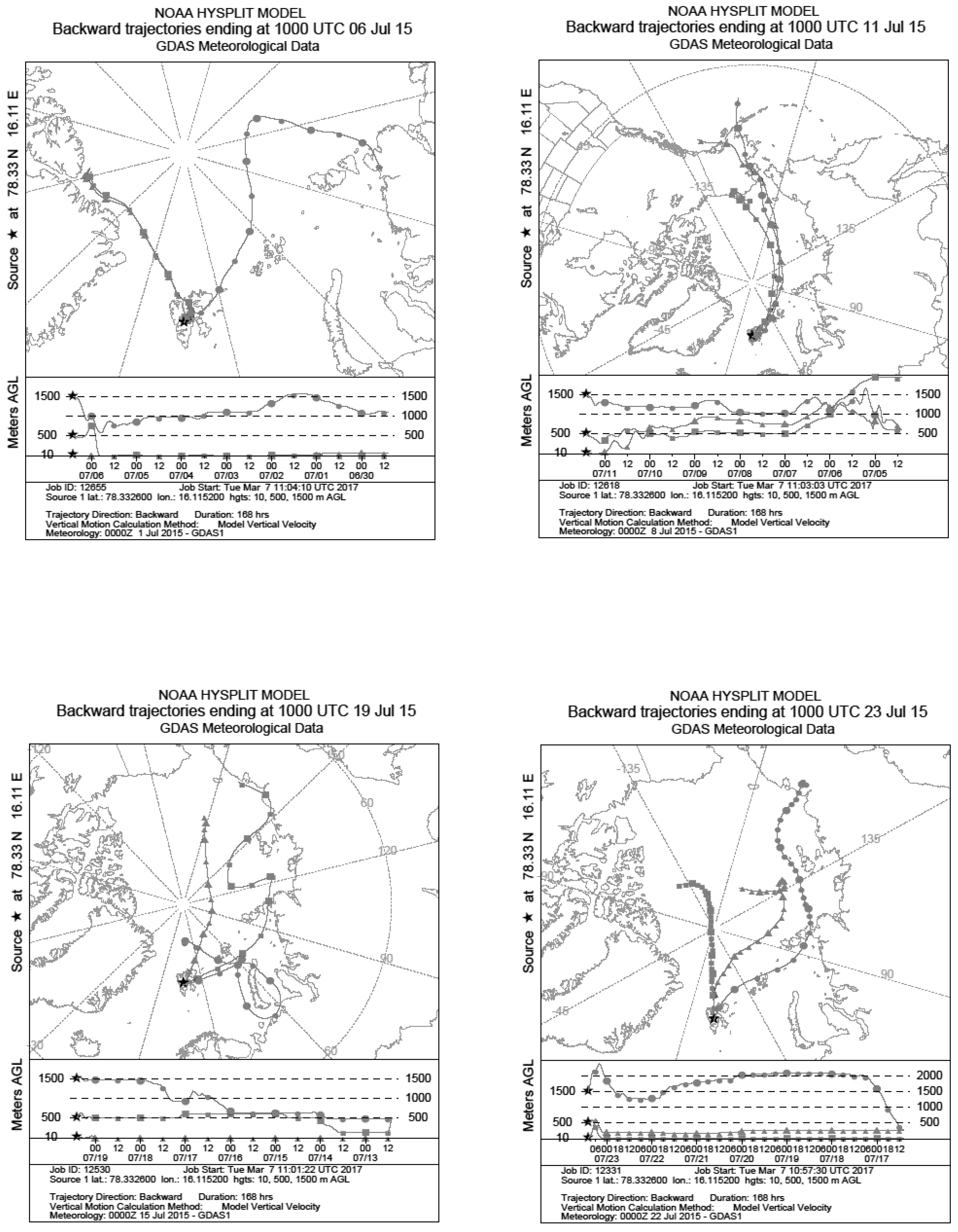
2.2. Meteorological Data
2.3. Culture Dependent
2.4. Culture Independent
2.5. Statistical Analysis
3. Results
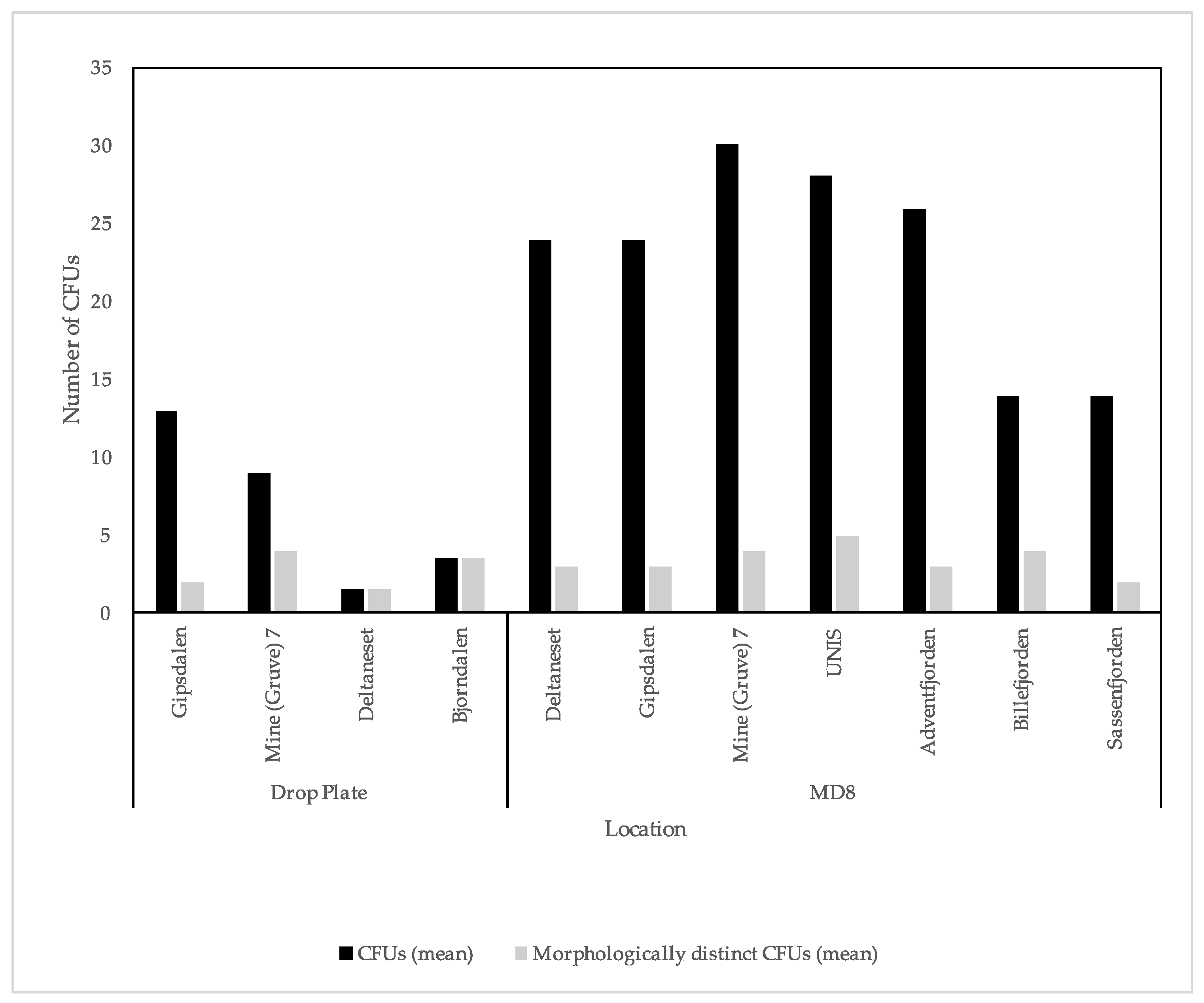
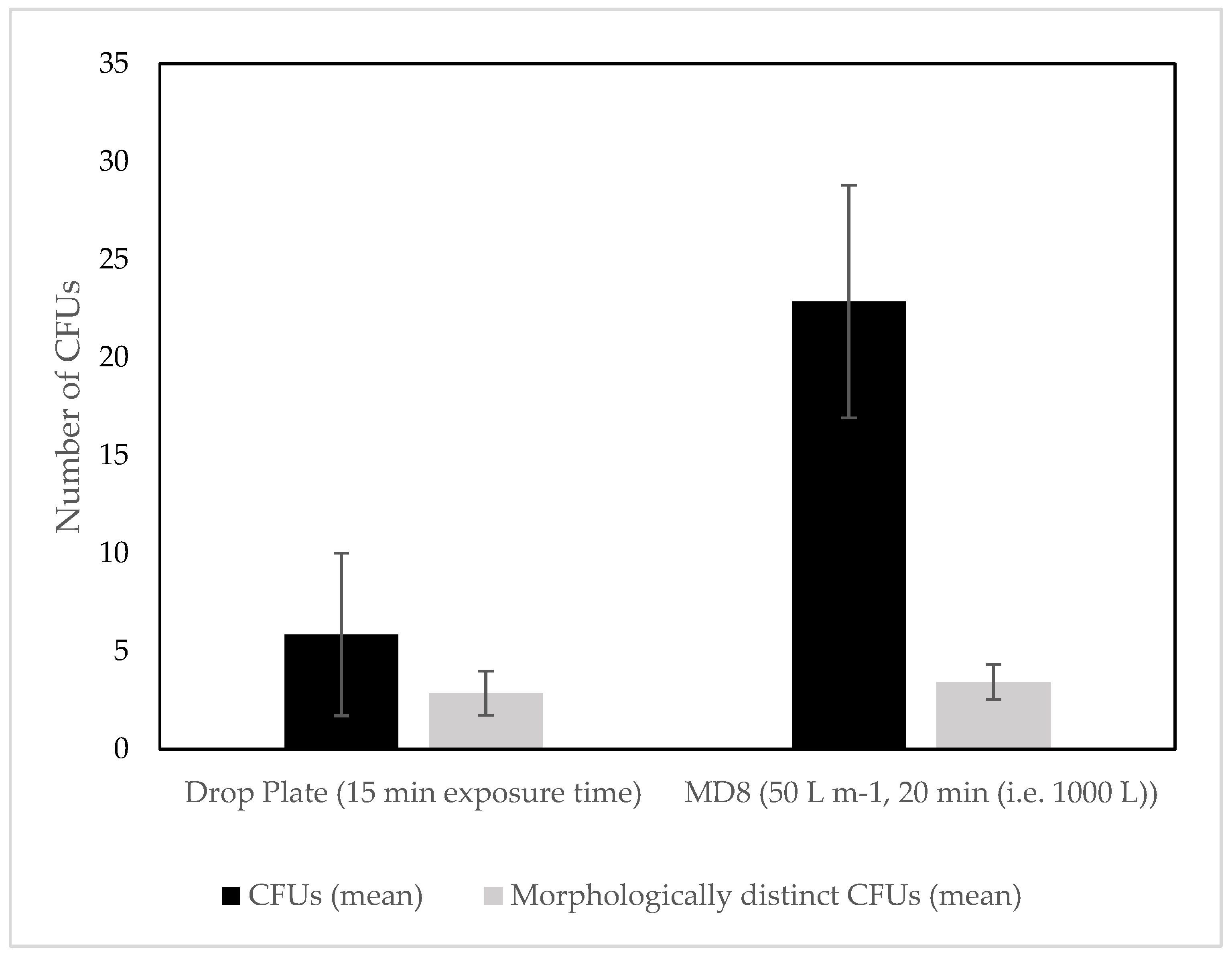
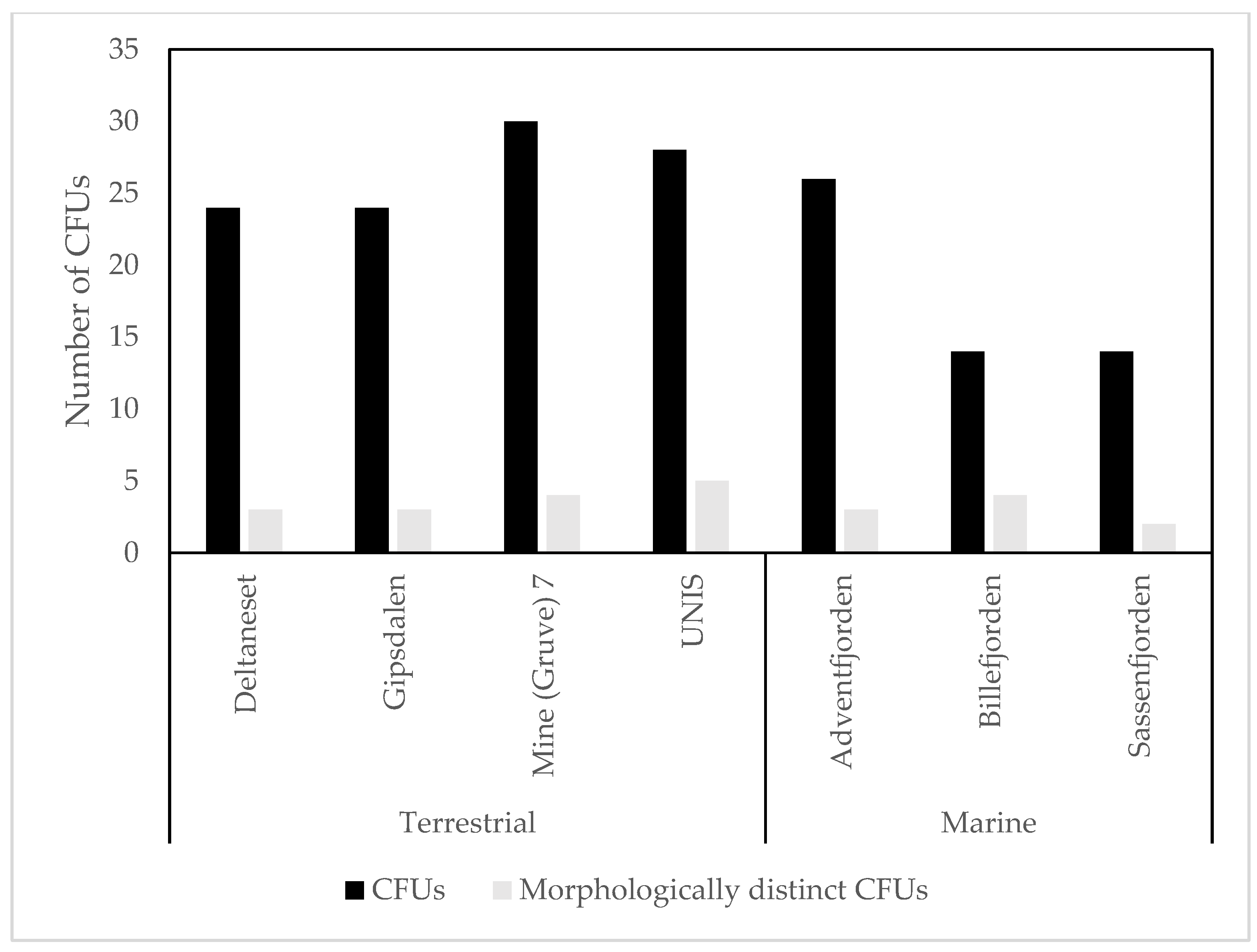
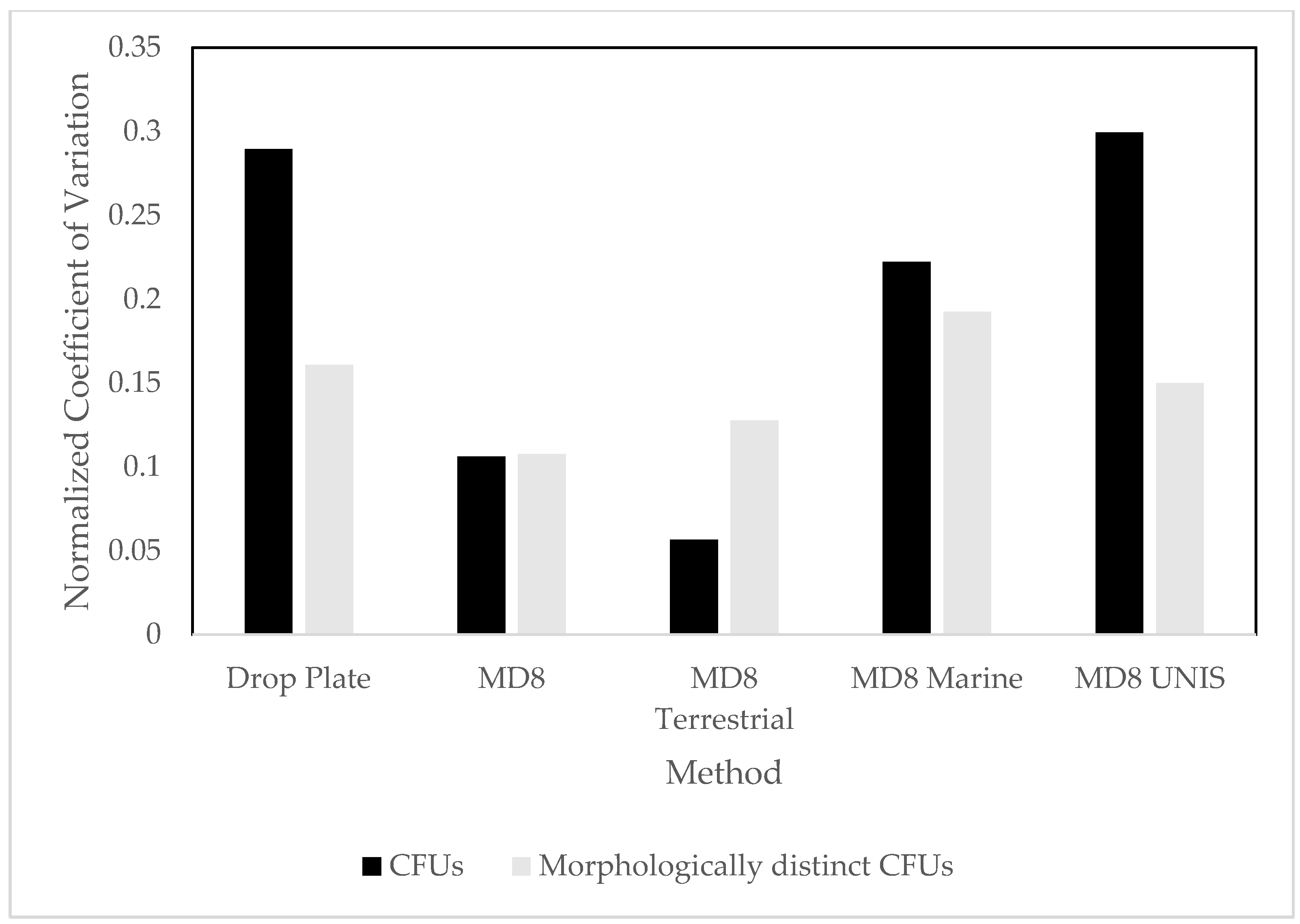
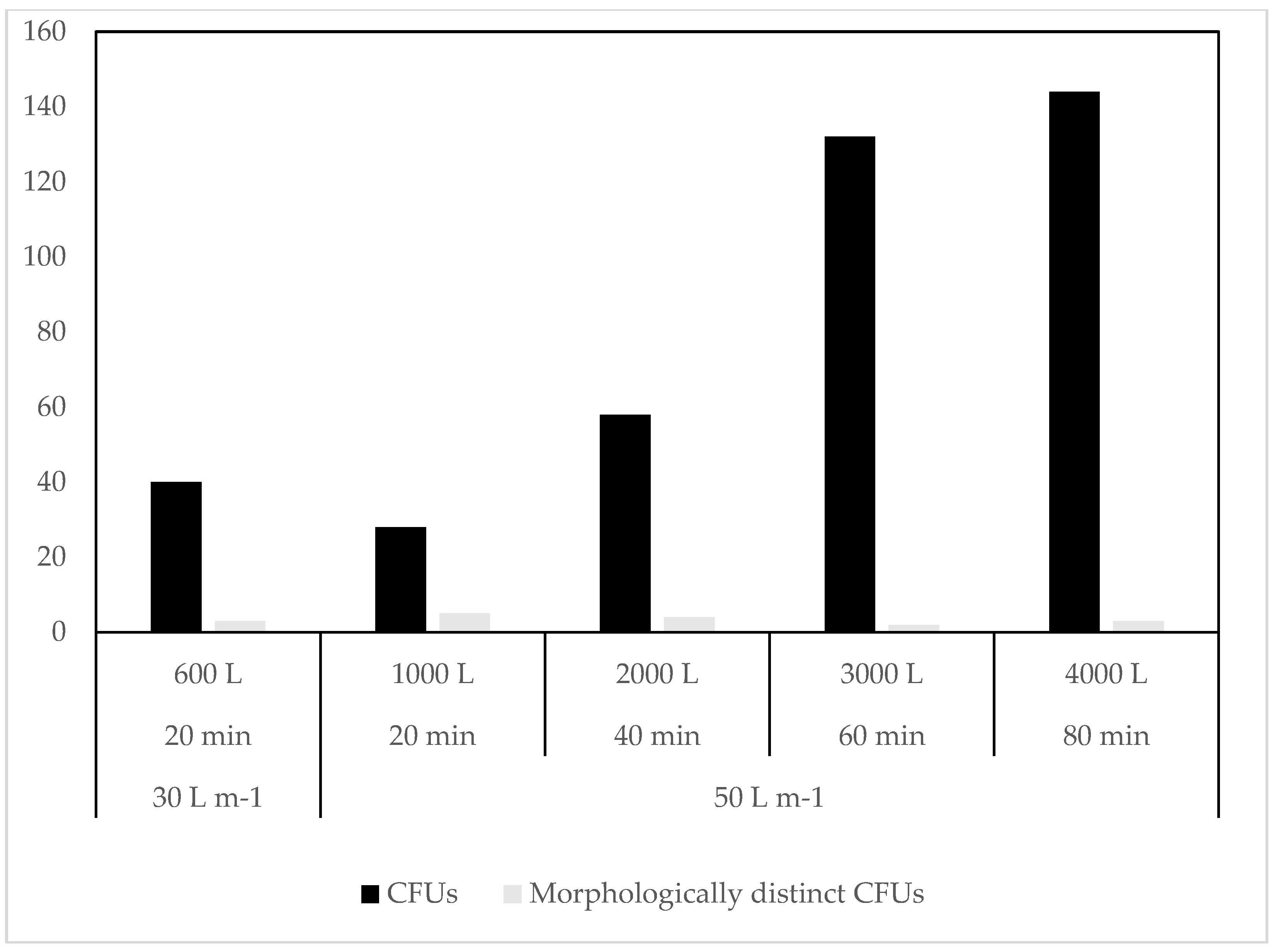
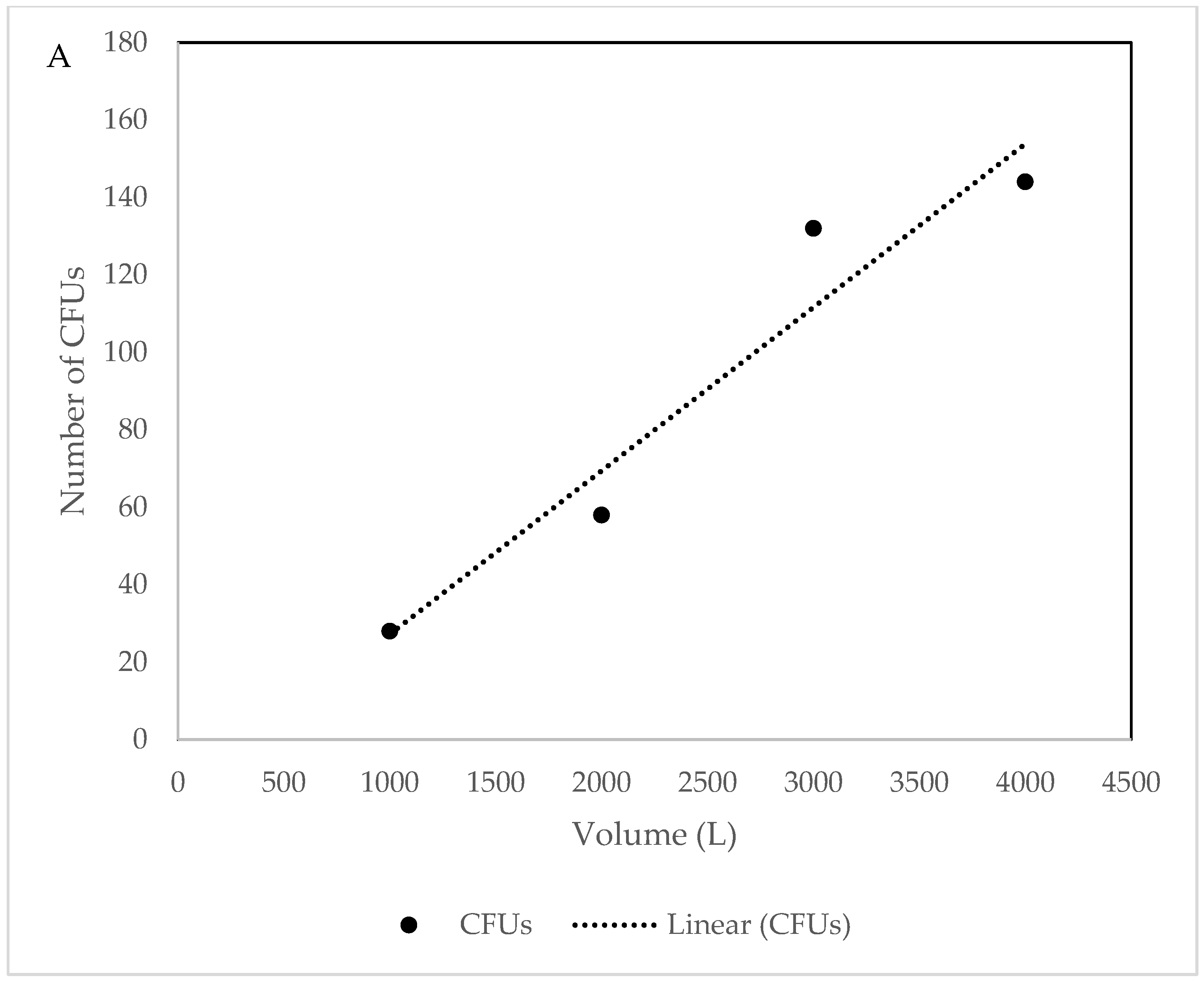
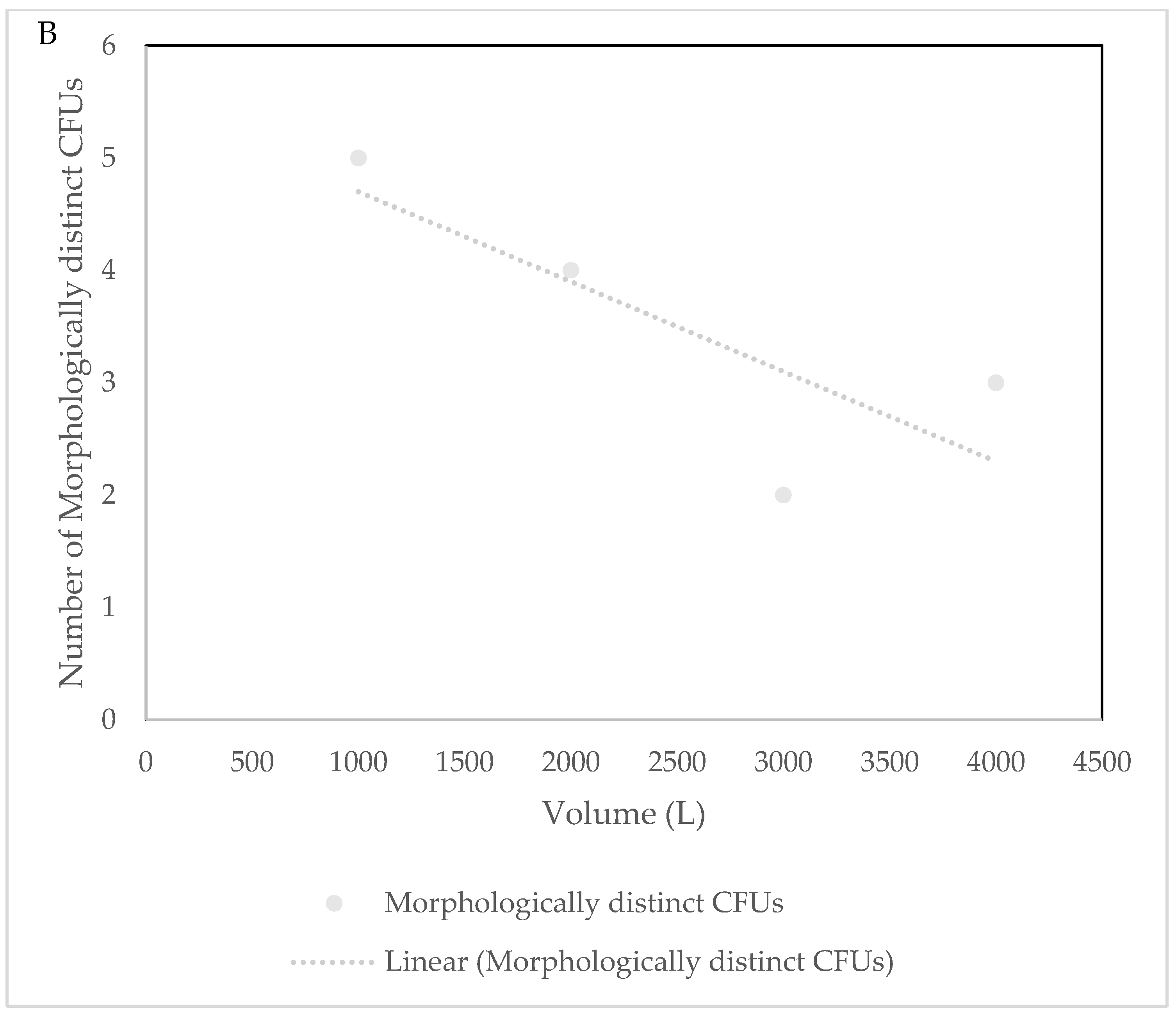
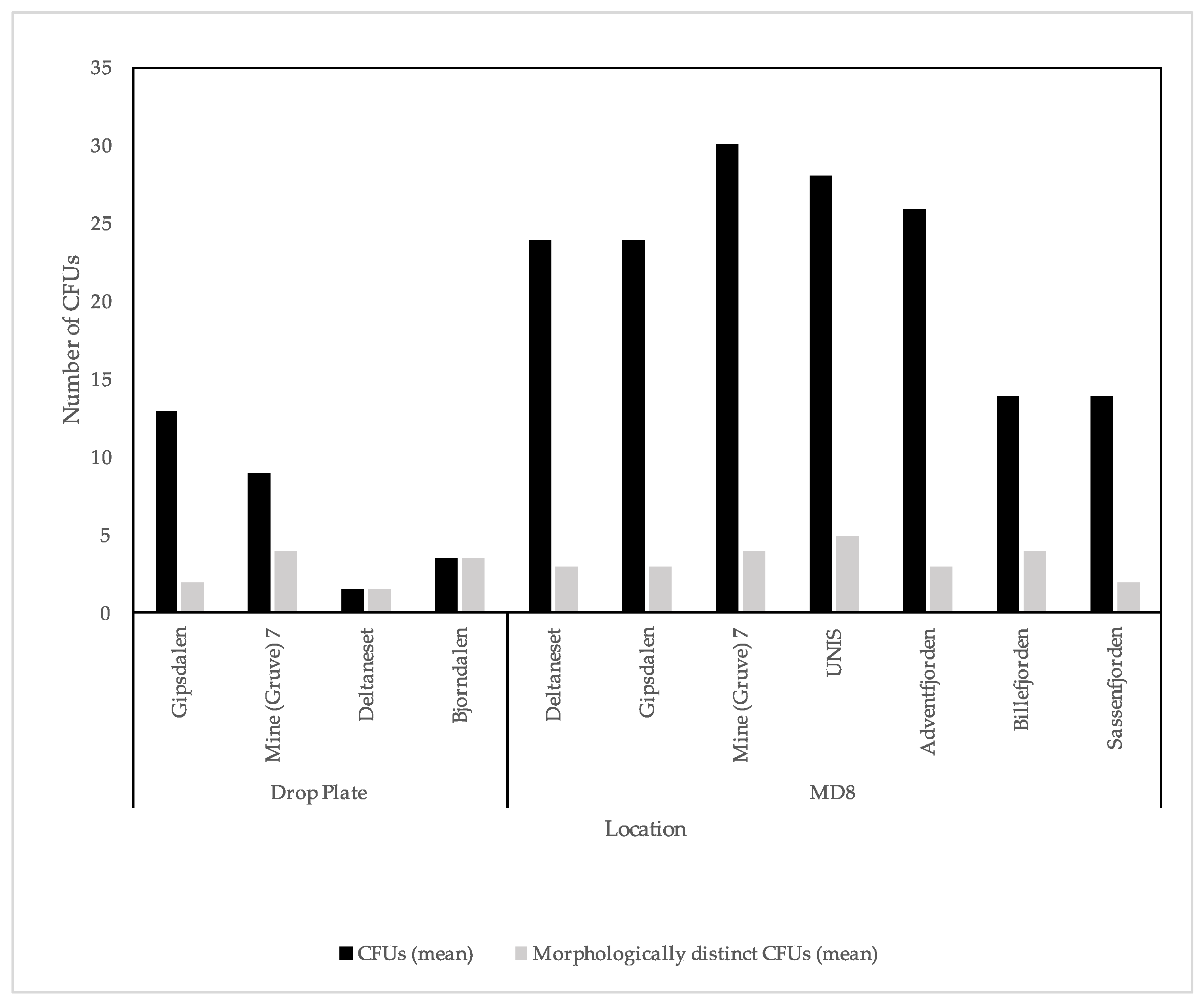
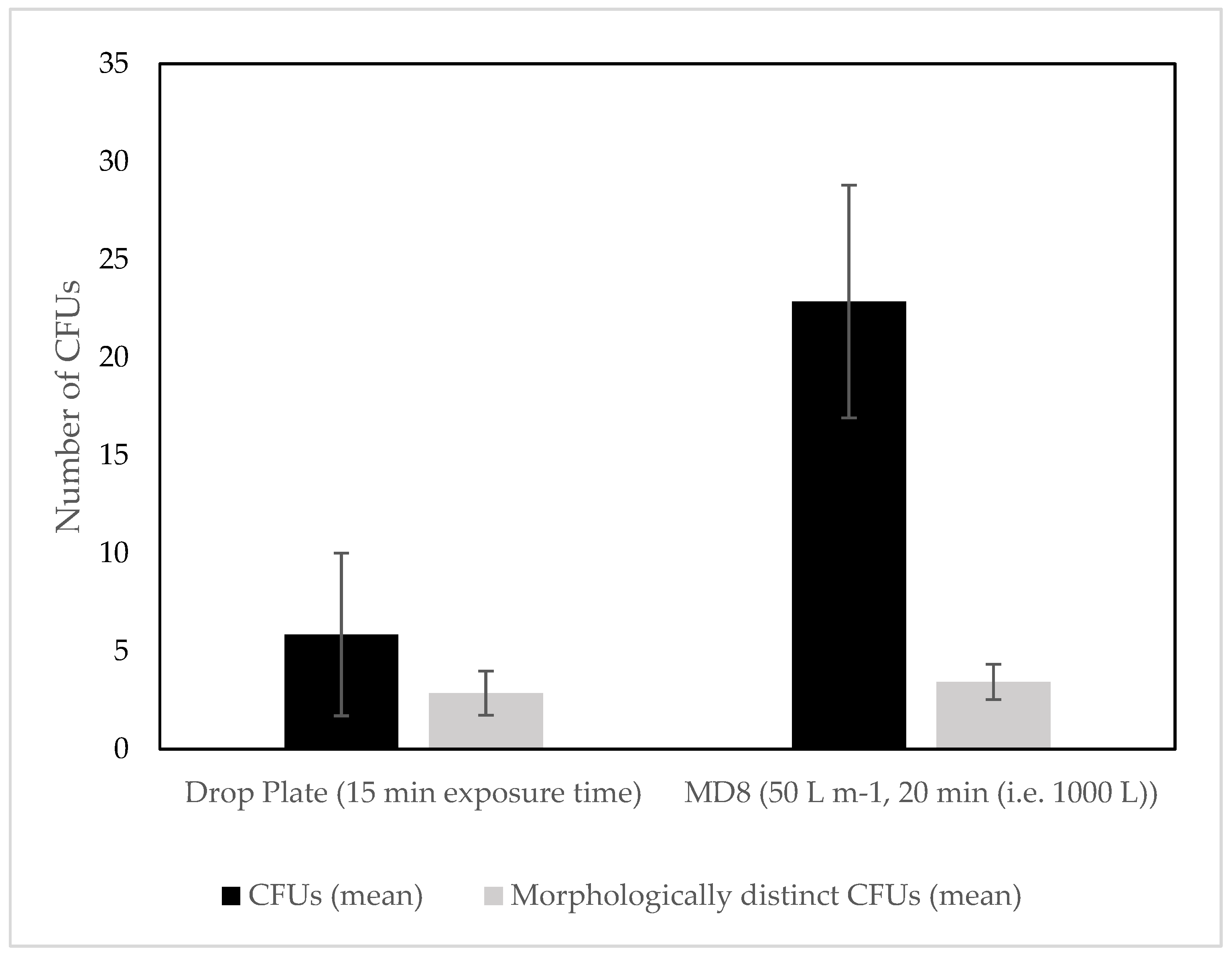
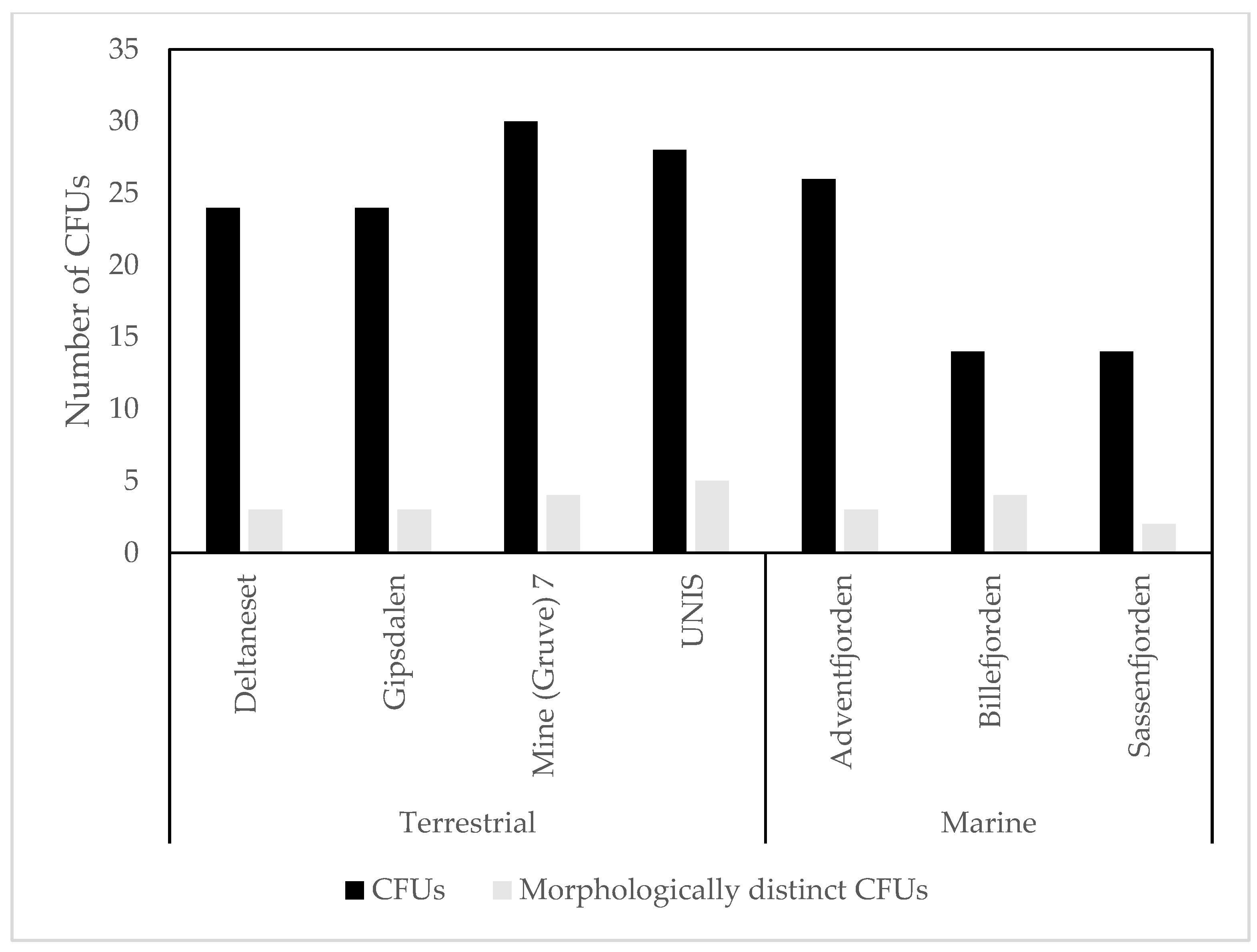
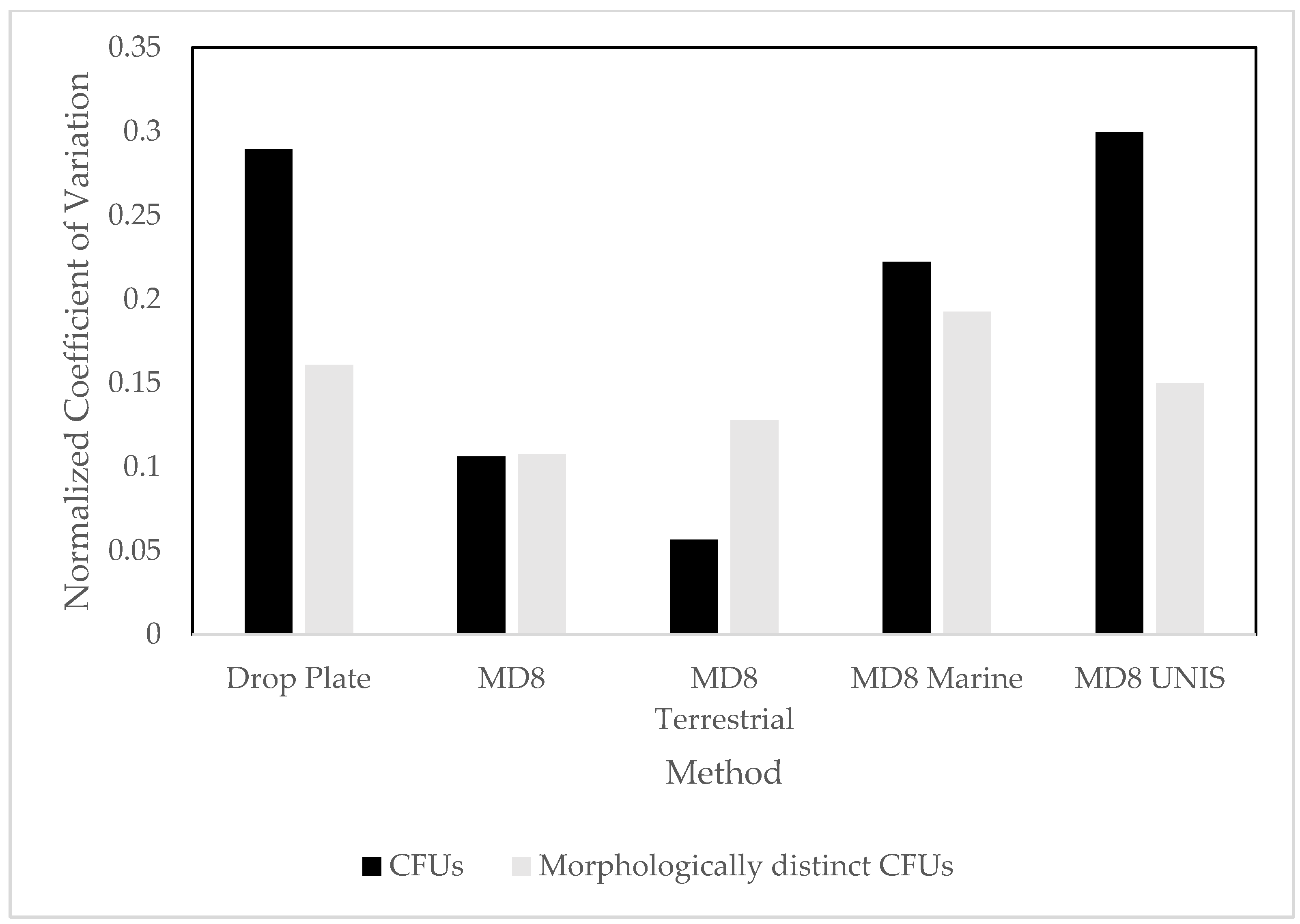
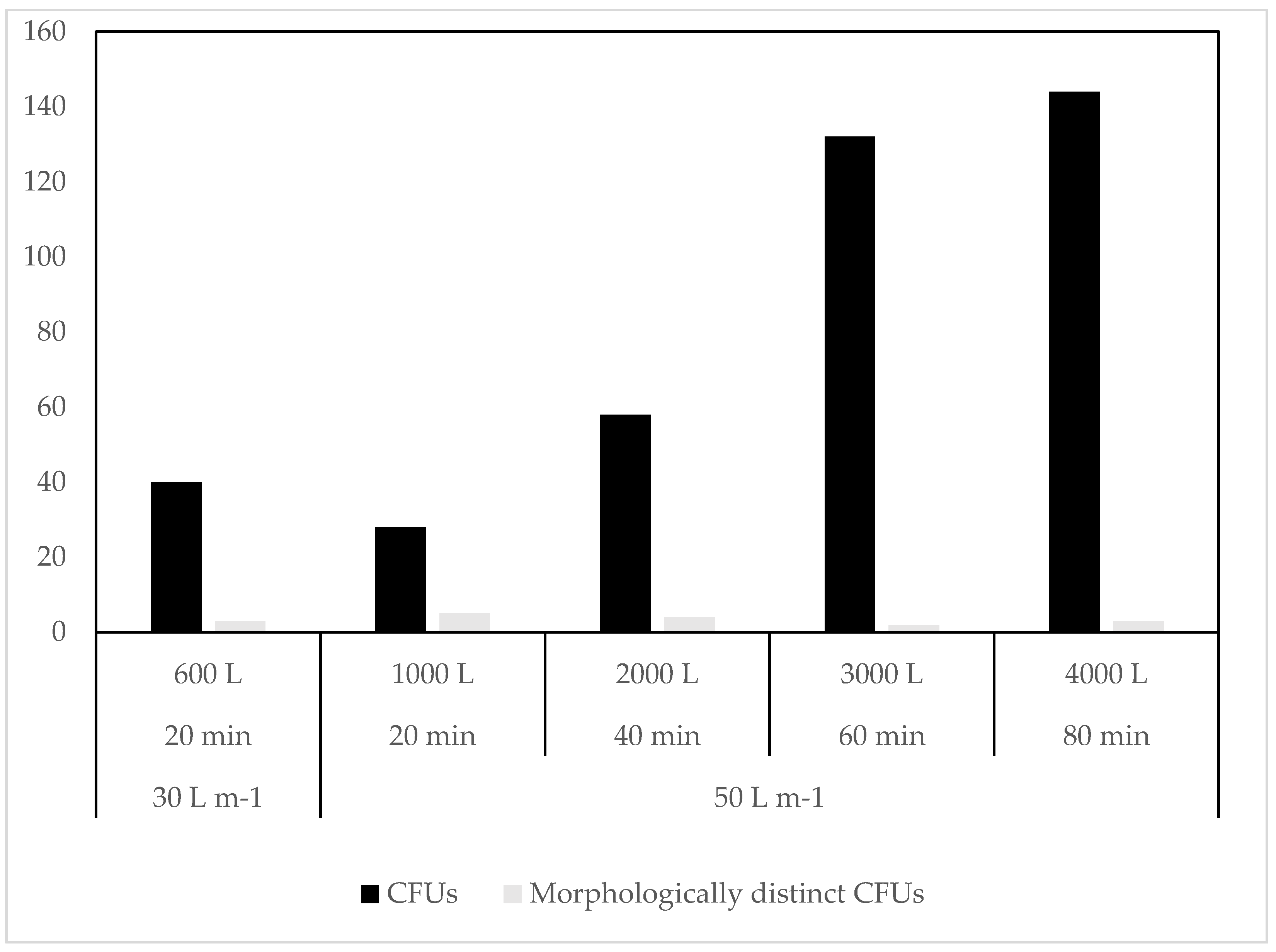
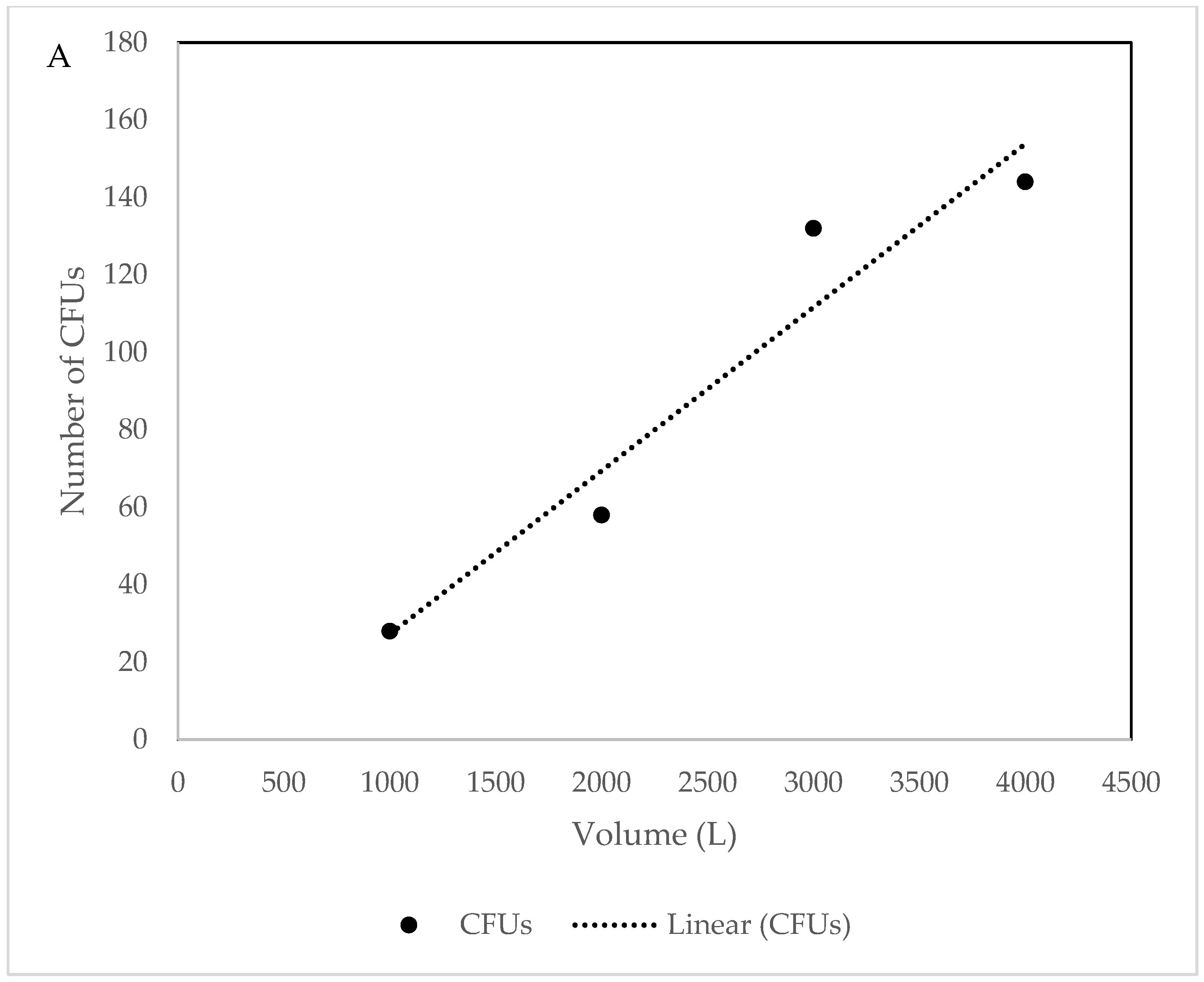
3.1. Culture Dependent
3.2. Culture Independent
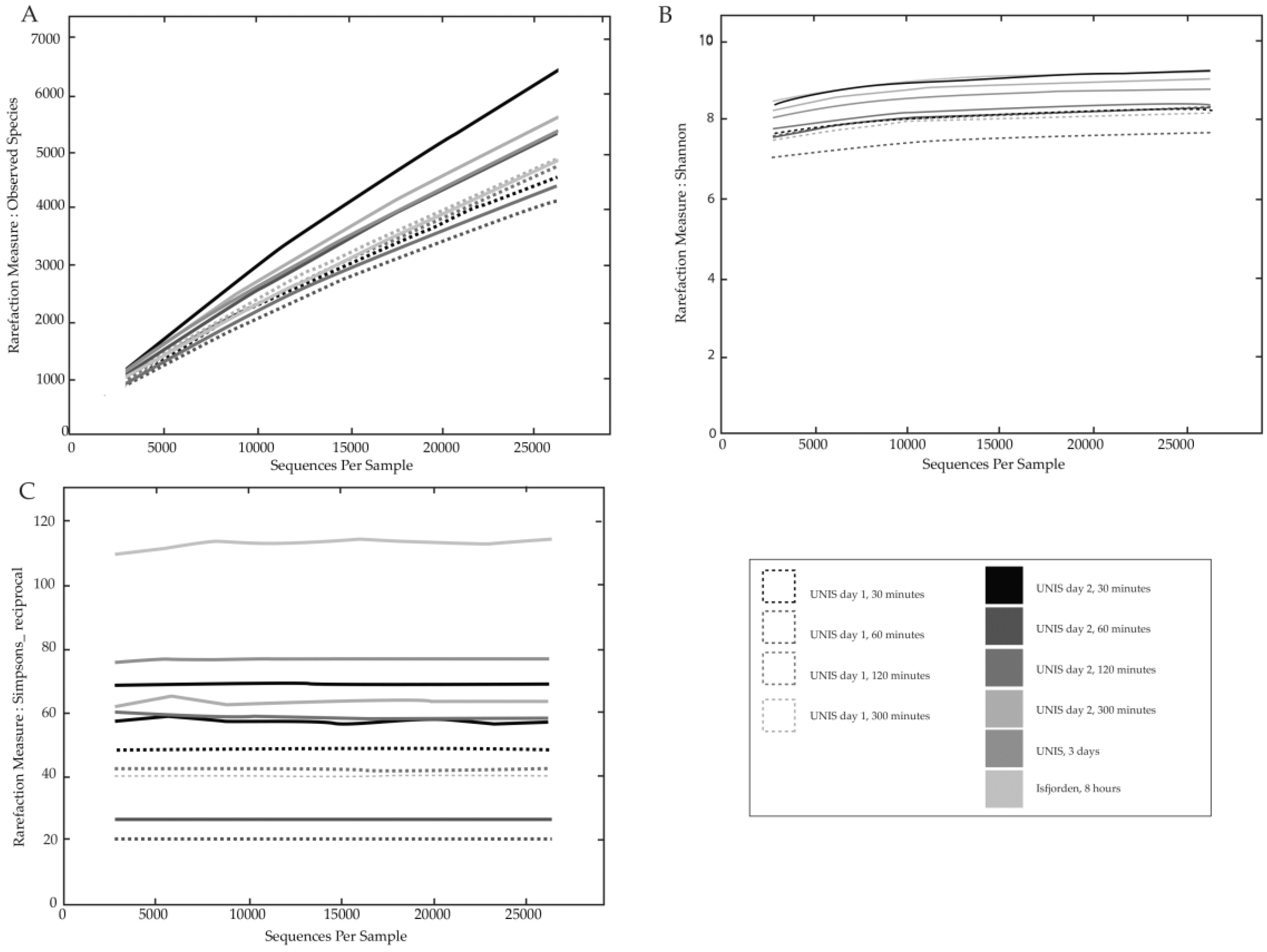
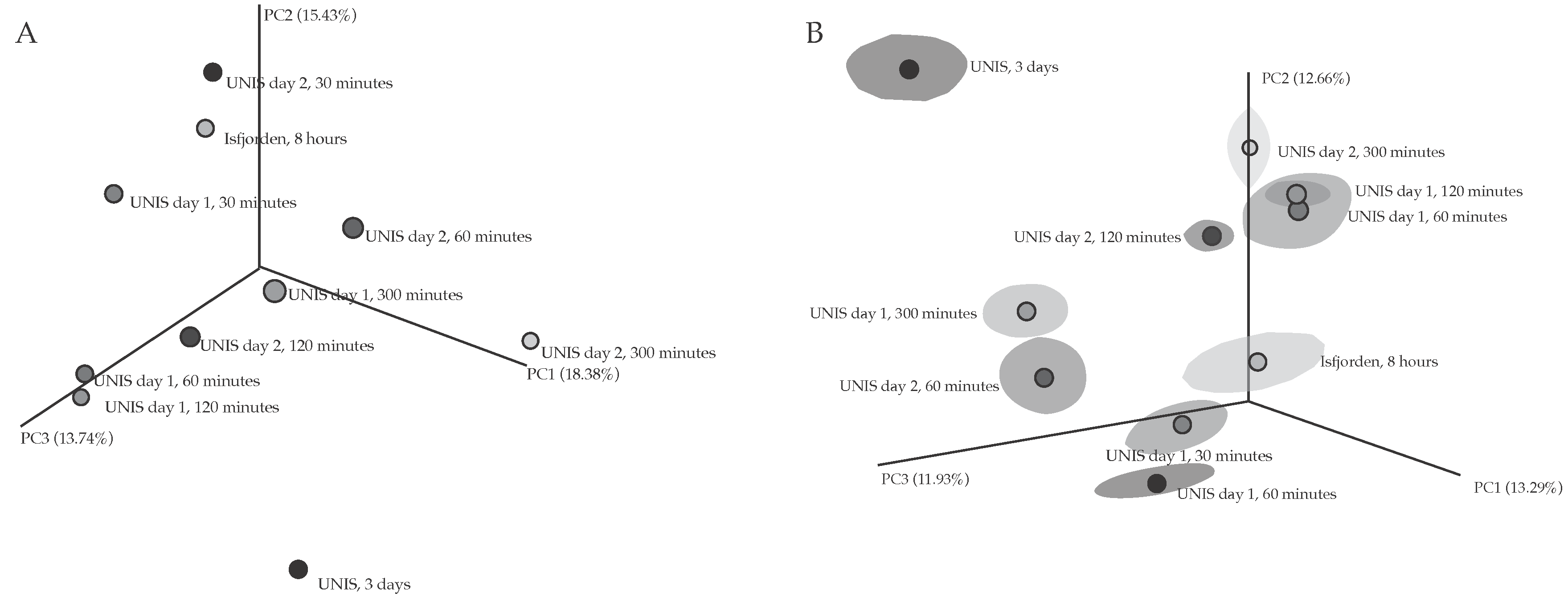
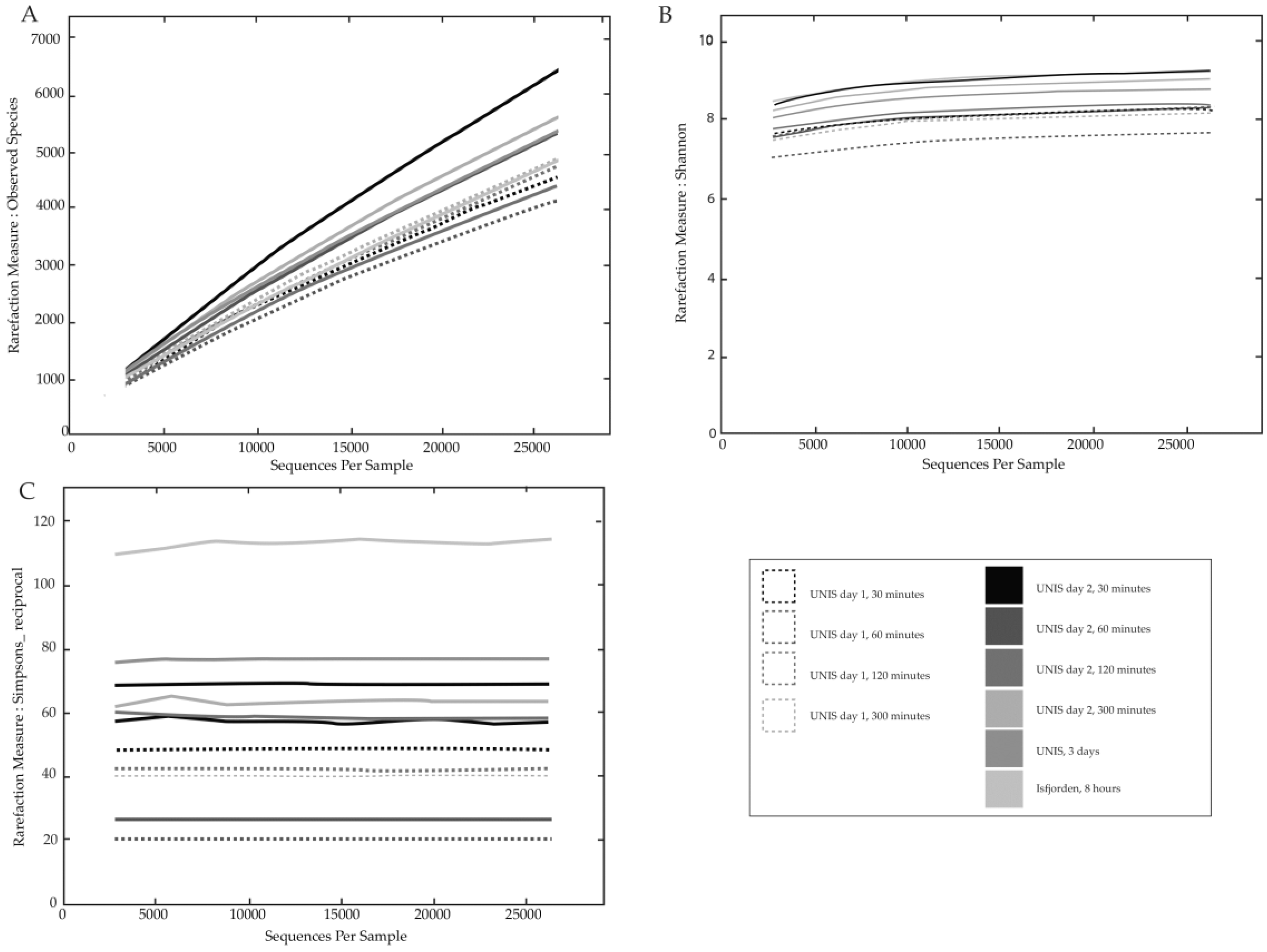
3.2.1. Bacterial Diversity
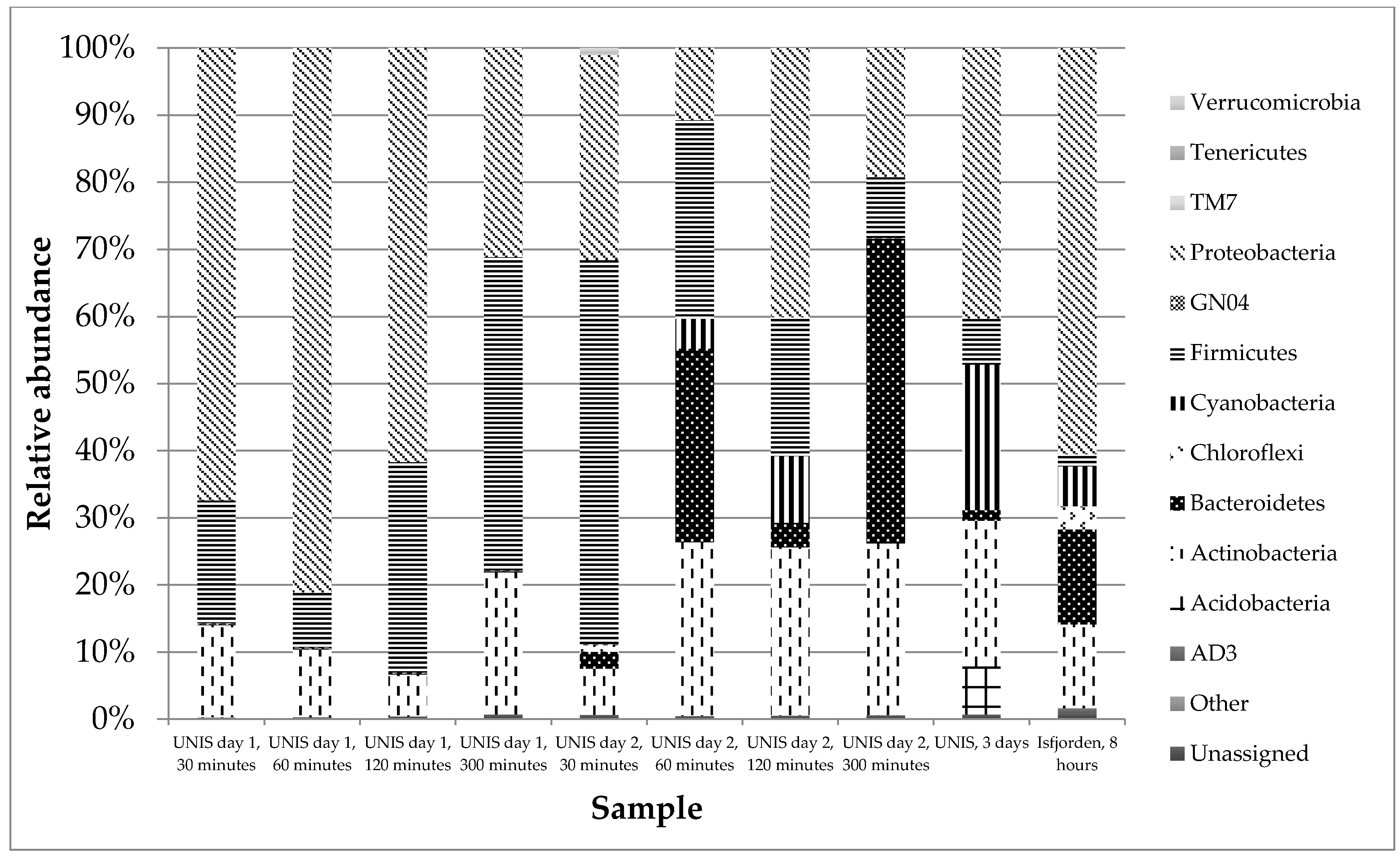
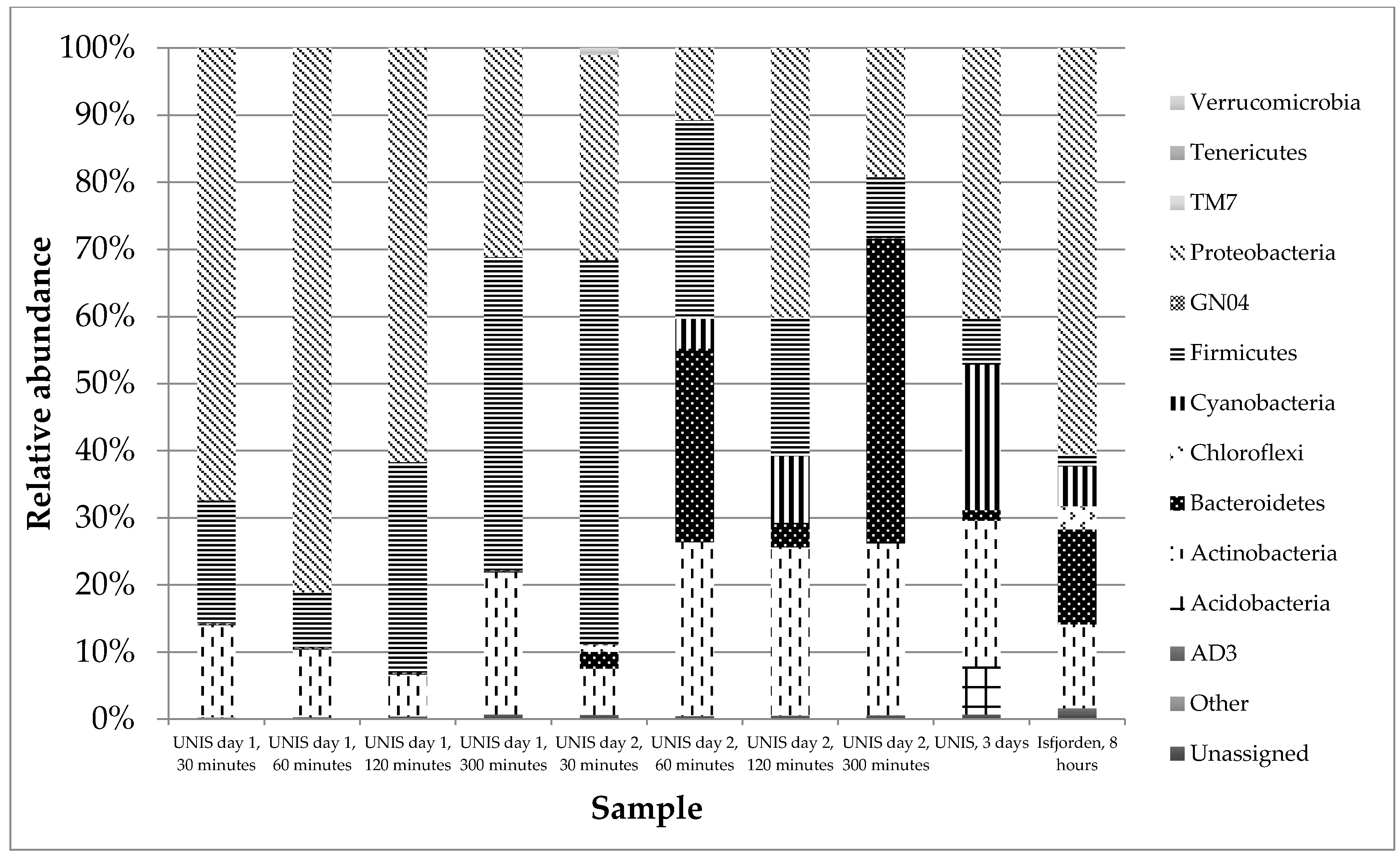
3.2.2. Taxonomy
4. Discussion
4.1. Culture Dependent
4.2. Culture Independent
4.2.1. Diversity
4.2.2. Taxonomy
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pearce, D.A.; Hughes, K.A.; Lachlan-Cope, T.; Harangozo, S.A.; Jones, A.E. Biodiversity of air-borne microorganisms at halley station, Antarctica. Extremophiles 2010, 14, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.W. Atmospheric movement of microorganisms in clouds of desert dust and implications for human health. Clin. Microbiol. Rev. 2007, 20, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Stewart, F.J. Where the genes flow. Nat. Geosci. 2013, 6, 688–690. [Google Scholar] [CrossRef]
- Weber, C.F.; Werth, J.T. Is the lower atmosphere a readily accessible reservoir of culturable, antimicrobial compound-producing Actinomycetales? Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N. Microbial biogeography: Patterns in microbial diversity across space and time. In Accessing Uncultivated Microorganisms; Zengler, K., Ed.; ASM Press: Washington, DC, USA, 2008. [Google Scholar]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Vali, G.; Christensen, M.; Fresh, R.W.; Galyan, E.L.; Maki, L.R.; Schnell, R.C. Biogenic Ice Nuclei. Part II: Bacterial Sources. J. Atmos. Sci. 1976, 33, 1565–1570. [Google Scholar] [CrossRef]
- Deguillaume, L.; Leriche, M.; Amato, P.; Ariya, P.A.; Delort, A.M.; Pöschl, U.; Chaumerliac, N.; Bauer, H.; Flossmann, A.I.; Morris, C.E. Microbiology and atmospheric processes: Chemical interactions of primary biological aerosols. Biogeosciences 2008, 5, 1073–1084. [Google Scholar] [CrossRef]
- Hill, K.A.; Shepson, P.B.; Galbavy, E.S.; Anastasio, C.; Kourtev, P.S.; Konopka, A.; Stirm, B.H. Processing of atmospheric nitrogen by clouds above a forest environment. J. Geophys. Res. 2007. [Google Scholar] [CrossRef]
- Ariya, P.A.; Nepotchatykh, O.; Ignatova, O.; Amyot, M. Microbiological degradation of atmospheric organic compounds. Geophys. Res. Lett. 2002, 29, 34-1–34-4. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F.; Marx, J.C.; Gerday, C. Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin, Germany, 2008; pp. 1–462. [Google Scholar]
- Giddings, S.N.; MacCready, P.; Hickey, B.M.; Banas, N.S.; Davis, K.A.; Siedlecki, S.A.; Trainer, V.L.; Kudela, R.M.; Pelland, N.A.; Connolly, T.P. Hindcasts of potential harmful algal bloom transport pathways on the pacific northwest coast. J. Geophys. Res. Oceans 2014, 119, 2439–2461. [Google Scholar] [CrossRef]
- Chen, P.S.; Tsai, F.T.; Lin, C.K.; Yang, C.Y.; Chan, C.C.; Young, C.Y.; Lee, C.H. Ambient influenza and avian influenza virus during dust storm days and background days. Environ. Health Perspect. 2010, 118, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Litchman, E. Invisible invaders: Non-pathogenic invasive microbes in aquatic and terrestrial ecosystems. Ecol. Lett. 2010, 13, 1560–1572. [Google Scholar] [CrossRef] [PubMed]
- Shinn, E.A.; Smith, G.W.; Prospero, J.M.; Betzer, P.; Hayes, M.L.; Garrison, V.; Barber, R.T. African dust and the demise of caribbean coral reefs. Geophys. Res. Lett. 2000, 27, 3029–3032. [Google Scholar] [CrossRef]
- Smith, D.J.; Griffin, D.W.; Jaffe, D.A. The high life: Transport of microbes in the atmosphere. Eos Trans. Am. Geophys. Union 2011, 92, 249–250. [Google Scholar] [CrossRef]
- Lighthart, B. Mini-review of the concentration variations found inthe alfresco atmospheric bacterial populations. Aerobiologia 2000, 16, 7–16. [Google Scholar] [CrossRef]
- Bowers, R.M.; McCubbin, I.B.; Hallar, A.G.; Fierer, N. Seasonal variability in airborne bacterial communities at a high-elevation site. Atmos. Environ. 2012, 50, 41–49. [Google Scholar] [CrossRef]
- Burrows, S.M.; Elbert, W.; Lawrence, M.G.; Pöschl, U. Bacteria in the global atmosphere–Part 1: Review and synthesis of literature data for different ecosystems. Atmos. Chem. Phys. 2009, 9, 9263–9280. [Google Scholar] [CrossRef]
- DeLeon-Rodriguez, N.; Lathem, T.L.; Rodriguez-R, L.M.; Barazesh, J.M.; Anderson, B.E.; Beyersdorf, A.J.; Ziemba, L.D.; Bergin, M.; Nenes, A.; Konstantinidis, K.T. Microbiome of the upper troposphere: Species composition and prevalence, effects of tropical storms, and atmospheric implications. Proc. Natl. Acad. Sci. USA 2013, 110, 2575–2580. [Google Scholar] [CrossRef] [PubMed]
- Fahlgren, C.; Hagstrom, A.; Nilsson, D.; Zweifel, U.L. Annual variations in the diversity, viability, and origin of airborne bacteria. Appl. Environ. Microbiol. 2010, 76, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Nonnenmann, M.W.; Bextine, B.; Dowd, S.E.; Gilmore, K.; Levin, J.L. Culture-independent characterization of bacteria and fungi in a poultry bioaerosol using pyrosequencing: A new approach. J. Occup. Environ. Hyg. 2010, 7, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.M.; Zervas, A.; Tendal, K.; Nielsen, J.L. Microbial diversity in bioaerosol samples causing ODTS compared to reference bioaerosol samples as measured using illumina sequencing and MALDI-TOF. Environ. Res. 2015, 140, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, A.; Medvedeva, S.; Shmakov, S.; Logacheva, M.D.; Krylenkov, V.; Severinov, K. Metagenomic analysis of bacterial communities of antarctic surface snow. Front. Microbiol. 2016, 7, 398. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Ladau, J.; Leff, J.W.; Pollard, K.S.; Menninger, H.L.; Dunn, R.R.; Fierer, N. Continental-scale distributions of dust-associated bacteria and fungi. Proc. Natl. Acad. Sci. USA 2015, 112, 5756–5761. [Google Scholar] [CrossRef] [PubMed]
- Seifried, J.S.; Wichels, A.; Gerdts, G. Spatial distribution of marine airborne bacterial communities. Microbiol. Open 2015, 4, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.H.; Bohannan, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Timonen, H.J.; Jaffe, D.A.; Griffin, D.W.; Birmele, M.N.; Perry, K.D.; Ward, P.D.; Roberts, M.S. Intercontinental dispersal of bacteria and archaea by transpacific winds. Appl. Environ. Microbiol. 2013, 79, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, C.; Vincent, W.F.; Bonilla, S.; Roy, S.; Martineau, M.-J.; Terrado, R.; Potvin, M.; Massana, R.; Pedrós-Alió, C. Distribution, phylogeny, and growth of cold-adapted picoprasinophytes in Arctic seas. J. Phycol. 2007, 43, 78–89. [Google Scholar] [CrossRef]
- Womack, A.M.; Bohannan, B.J.M.; Green, J.L. Biodiversity and biogeography of the atmosphere. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 3645–3653. [Google Scholar] [CrossRef] [PubMed]
- Population of Svalbard, 1 January 2015. Avaliable online: https://www.ssb.no/en/befolkning/statistikker/befsvalbard/halvaar/2015-04-09 (accessed on 5 March 2017).
- Polunin, N.; Pady, S.M.; Kelly, C.D. Arctic aerobiology. Nature 1947, 160, 876–877. [Google Scholar] [CrossRef] [PubMed]
- Harding, T.; Jungblut, A.D.; Lovejoy, C.; Vincent, W.F. Microbes in High Arctic snow and implications for the cold biosphere. Appl. Environ. Microbiol. 2011, 77, 3234–3243. [Google Scholar] [CrossRef] [PubMed]
- Amato, P.; Hennebelle, R.; Magand, O.; Sancelme, M.; Delort, A.-M.; Barbante, C.; Boutron, C.; Ferrari, C. Bacterial characterization of the snow cover at Spitzberg, Svalbard. FEMS Microbiol. Ecol. 2007, 59, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.K.; Søborg, D.A.; Al-Soud, W.A.; Sørensen, S.J.; Kroer, N. Bacterial community structure in high-arctic snow and freshwater as revealed by pyrosequencing of 16s rrna genes and cultivation. J. Pol. Res. 2013, 32. [Google Scholar] [CrossRef]
- Herbold, C.W.; Lee, C.K.; McDonald, I.R.; Cary, S.C. Evidence of global-scale aeolian dispersal and endemism in isolated geothermal microbial communities of Antarctica. Nat. Commun. 2014, 5, 3875. [Google Scholar] [CrossRef] [PubMed]
- Pearce, D.A.; Alekhina, I.A.; Terauds, A.; Wilmotte, A.; Quesada, A.; Edwards, A.; Dommergue, A.; Sattler, B.; Adams, B.J.; Magalhães, C.; et al. Aerobiology over Antarctica–A new initiative for atmospheric ecology. Front. Microbiol. 2016, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Liu, Z.; Rodríguez-Hernández, M.; Knight, R.; Henn, M.; Hernandez, M.T. Short-term temporal variability in airborne bacterial and fungal populations. Appl. Environ. Microbiol. 2008, 74, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.M.; McLetchie, S.; Knight, R.; Fierer, N. Spatial variability in airborne bacterial communities across land-use types and their relationship to the bacterial communities of potential source environments. ISME J. 2011, 5, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Brodie, E.L.; DeSantis, T.Z.; Parker, J.P.; Zubietta, I.X.; Piceno, Y.M.; Andersen, G.L. Urban aerosols harbor diverse and dynamic bacterial populations. Proc. Nat. Acad. Sci. USA 2007, 104, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.F.; Draxler, R.R.; Rolph, G.D.; Stunder, B.J.; Cohen, M.D.; Ngan, F. NOAA’S hysplit atmospheric transport and dispersion modeling system. Bull. Am. Meteorol. Soc. 2015, 96, 2059–2077. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than blast. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16s rRNA gene database and workbench compatible with Arb. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. Pynast: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. Fasttree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Hammer Oyvind, D.H. , Paul Ryan. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Grzesiak, J.; Górniak, D.; Świątecki, A.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Zdanowski, M.K. Microbial community development on the surface of Hans and Werenskiold glaciers (Svalbard, Arctic): A comparison. Extremophiles Life Extreme Cond. 2015, 19, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.H.; Hou, S.G.; Yang, G.L.; Wang, J.H. Bacterial community in the East Rongbuk Glacier, Mt. Qomolangma (Everest) by culture and culture-independent methods. Microbiol. Res. 2010, 165, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Singh, S.M.; Roy, U. Taxonomic characterization and the bio-potential of bacteria isolated from glacier ice cores in the High Arctic. J. Basic Microbiol. 2016, 56, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Salustiano, V.C.; Andrade, N.J.; Brandão, S.C.C.; Azeredo, R.M.C.; Lima, S.A.K. Microbiological air quality of processing areas in a dairy plant as evaluated by the sedimentation technique and a one-stage air sampler. Braz. J. Microbiol. 2003, 34, 255–259. [Google Scholar] [CrossRef]
- Li, M.; Qi, J.; Zhang, H.; Huang, S.; Li, L.; Gao, D. Concentration and size distribution of bioaerosols in an outdoor environment in the qingdao coastal region. Sci. Total Environ. 2011, 409, 3812–3819. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, R.; Kozłowska, K.; Szpakowska, M.; Trafny, E.A. Evaluation of applicability of the sartorius airport MD8 sampler for detection of bacillus endospores in indoor air. Environ. Monit. Assess. 2013, 185, 3517–3526. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.L.; Grinshpun, S.A.; Willeke, K.; Terzieva, S.; Ulevicius, V.; Donnelly, J. Effect of impact stress on microbial recovery on an agar surface. Appl. Environ. Microbiol. 1995, 61, 1232–1239. [Google Scholar] [PubMed]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. Strategies for culture of ‘unculturable’ bacteria. FEMS Microbiol. Lett. 2010, 309, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pankhurst, L.J.; Whitby, C.; Pawlett, M.; Larcombe, L.D.; McKew, B.; Deacon, L.J.; Morgan, S.L.; Villa, R.; Drew, G.H.; Tyrrel, S.; et al. Temporal and spatial changes in the microbial bioaerosol communities in green-waste composting. FEMS Microbiol. Ecol. 2012, 79, 229. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.; Saetnan, E.R.; Scullion, J.; Gwynn-Jones, D.; Ostle, N.; Edwards, A. Temporal and spatial influences incur reconfiguration of arctic heathland soil bacterial community structure. Environ. Microbiol. 2016, 18, 1942–1953. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.-K.; Kim, J.; Ha, S.-M.; Oh, H.-S.; Chun, J.; Sohn, J.; Yi, H. Metagenomic insights into the bioaerosols in the indoor and outdoor environments of childcare facilities. PLoS ONE 2015, 10, e0126960. [Google Scholar] [CrossRef] [PubMed]
- Galperin, M.Y. Genome diversity of spore-forming Firmicutes. Microbiol. Spectr. 2013, 1. TBS-0015-2012. [Google Scholar] [CrossRef] [PubMed]
- Lightfield, J.; Fram, N.R.; Ely, B. Across bacterial phyla, distantly-related genomes with similar genomic GC content have similar patterns of amino acid usage. PLoS ONE 2011, 6, e17677. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. The phylogeny of Proteobacteria: Relationships to other eubacterial phyla and eukaryotes. FEMS Microbiol. Rev. 2000, 24, 367–402. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.M.; Clements, N.; Emerson, J.B.; Wiedinmyer, C.; Hannigan, M.P.; Fierer, N. Seasonal variability in bacterial and fungal diversity of the near-surface atmosphere. Environ. Sci. Technol. 2013, 47, 12097–12106. [Google Scholar] [CrossRef] [PubMed]
- Stibal, M.; Gözdereliler, E.; Cameron, K.A.; Box, J.E.; Stevens, I.T.; Gokul, J.K.; Schostag, M.; Zarsky, J.D.; Edwards, A.; Irvine-Fynn, T.D.L.; et al. Microbial abundance in surface ice on the Greenland Ice Sheet. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.X.; Yan, M.; Yu, Y.; Li, H.R.; He, J.F.; Sun, K.; Zhang, F. Diversity of bacteria in surface ice of Austre Lovenbreen Glacier, Svalbard. Arch. Microbiol. 2013, 195, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Lysnes, K.; Thorseth, I.H.; Steinsbu, B.O.; Øvreås, L.; Torsvik, T.; Pedersen, R.B. Microbial community diversity in seafloor basalt from the Arctic spreading ridges. FEMS Microbiol. Ecol. 2004, 50, 213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Cao, T.; Ying, J.; Yang, Y.; Ma, L. Diversity and novelty of Actinobacteria in Arctic marine sediments. Antonie van Leeuwenhoek 2014, 105, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Choidash, B.; Begum, Z.; Shivaji, S. Bacterial diversity of Ny-Ålesund, Arctic Archipelago Svalbard. Mong. J. Biol. Sci. 2012, 10, 67–72. [Google Scholar]
- Abraham, W.P.; Thomas, S. Draft genome sequence of pseudomonas psychrophila MTCC 12324, isolated from the arctic at 79° N. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Sanschagrin, S.; Beaumier, D.; Greer, C.W. Metagenomic analysis of the bioremediation of diesel-contaminated canadian High Arctic soils. PLoS ONE 2012, 7, e30058. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.L.; Hou, S.G.; Le Baoge, R.; Li, Z.G.; Xu, H.; Liu, Y.P.; Du, W.T.; Liu, Y.Q. Differences in bacterial diversity and communities between glacial snow and glacial soil on the chongce ice cap, west Kunlun Mountains. Sci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Schneiker, S.; Martins dos Santos, V.A.; Bartels, D.; Bekel, T.; Brecht, M.; Buhrmester, J.; Chernikova, T.N.; Denaro, R.; Ferrer, M.; Gertler, C.; et al. Genome sequence of the ubiquitous hydrocarbon-degrading marine bacterium alcanivorax borkumensis. Nat. Biotechnol. 2006, 24, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.M.; Lee, K.; Jang, Y.; Kang, I.; Kim, H.J.; Kang, T.W.; Kim, S.Y.; Cho, J.C. Genome sequence of strain IMCC9480, a xanthorhodopsin-bearing betaproteobacterium isolated from the Arctic Ocean. J. Bacteriol. 2011, 193, 3421. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.H.; Yergeau, E.; Maynard, C.; Juck, D.; Whyte, L.G.; Greer, C.W. Predictable bacterial composition and hydrocarbon degradation in arctic soils following diesel and nutrient disturbance. ISME J. 2013, 7, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.V.; Whyte, L.G. Polar Microbiology: Life in a Deep Freeze; ASM Press: Washington, DC, USA, 2012. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Location | Environment | Sampling Mechanism | Date | Flow Rate (L m−1) | Duration (min) |
|---|---|---|---|---|---|
| Bjørndalen | Terrestrial | Drop plates | 13 July 2015 | - | 15 |
| Deltaneset | Terrestrial | Impaction onto media | 13 July 2015 | 50 | 20 |
| Drop plates | 13 July 2015 | - | 15 | ||
| Gipsdalen | Terrestrial | Impaction onto media | 13 July 2015 | 50 | 20 |
| Drop plates | 13 July 2015 | - | 15 | ||
| Longyearbyen | Terrestrial | Impaction onto media | 16 July 2015 | 30, 50 | 20, 40, 60, 80 |
| Membrane filtration | 06, 19, 21–23 July 2015 | ~20 | 30, 60, 120, 300, 3 days | ||
| Mine (Gruve) 7 | Terrestrial | Impaction onto media | 13 July 2015 | 50 | 20 |
| Drop plates | 13 July 2015 | - | 15 | ||
| Adventfjorden | Marine | Impaction onto media | 17 July 2015 | 50 | 20 |
| Billefjorden | Marine | Impaction onto media | 17 July 2015 | 50 | 20 |
| Isfjorden | Marine | Membrane filtration | 11 July 2015 | ~20 | 480 |
| Sassenfjorden | Marine | Impaction onto media | 17 July 2015 | 50 | 20 |
| 06 July 2015 | 11 July 2015 | 13 July 2015 | 16 July 2015 | 17 July 2015 | 19 July 2015 | 21 July 2015 | 22 July 2015 | 23 July 2015 | 21–23 July 2015 (Average) | Average across All Sampling Days | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Average temperature (°C) | 8 | 10 | 8 | 6 | 8 | 8 | 10 | 8 | 6 | 8 | 8 |
| Total precipitation (mm) | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Average wind speed (km h−1) | 13 | 20 | 10 | 20 | 14 | 22 | 18 | 12 | 12 | 14 | 16 |
| Average humidity (%) | 63 | 75 | 90 | 57 | 68 | 59 | 65 | 68 | 61 | 65 | 67 |
| Pressure (hPa) | 1025 | 1022 | 1019 | 1009 | 1013 | 1015 | 1015 | 1013 | 1006 | 1012 | 1016 |
| UNIS Day 1, 30 min | UNIS Day 1, 60 min | UNIS Day 1, 120 min | UNIS Day 1, 300 min | ||||
| OTU assignment | Relative abundance | OTU assignment | Relative abundance | OTU assignment | Relative abundance | OTU assignment | Relative abundance |
| Pseudomonas | 38% | Pseudomonadaceae | 39% | Pseudomonadaceae | 31% | Corynebacterium | 17% |
| Acinetobacter | 13% | Corynebacterium | 22% | Staphylococcus | 22% | Unassigned | 16% |
| Bacillales | 10% | Pseudomonas | 13% | Unassigned | 12% | Acinetobacter | 14% |
| Corynebacterium | 9% | Unassigned | 8% | Streptophyta | 9% | Gaiellaceae | 14% |
| Staphylococcus | 8% | Micrococcus | 7% | Bacillales | 9% | Pseudomonadaceae | 9% |
| Pseudomonadaceae | 8% | Bacillales | 3% | Pseudomonas | 6% | Pseudomonadaceae | 9% |
| Delftia | 7% | Gammaproteobacteria | 2% | Pseudomonadaceae | 3% | Staphylococcus | 7% |
| Propionibacterium | 5% | Gaiellaceae | 2% | Burkholderiales | 3% | Bacillales | 5% |
| Unassigned | 0% | Nocardioidaceae | 1% | Acetobacteraceae | 2% | Acetobacteraceae | 3% |
| Cloacibacterium | 0% | Acinetobacter | 0% | Sphingomonas | 1% | Alcaligenaceae | 2% |
| UNIS Day 2, 30 min | UNIS Day 2, 60 min | UNIS Day 2, 120 min | UNIS Day 2, 300 min | ||||
| OTU assignment | Relative abundance | OTU assignment | Relative abundance | OTU assignment | Relative abundance | OTU assignment | Relative abundance |
| Unassigned | 49% | Oxalobacteraceae | 27% | Oxalobacteraceae | 26% | Comamonadaceae | 36% |
| Corynebacterium | 18% | Acinetobacter | 15% | Staphylococcus | 20% | Candidate division TM7 | 24% |
| Alcaligenaceae | 7% | Arthrobacter | 14% | Arthrobacter | 11% | Alcanivorax | 6% |
| Staphylococcus | 6% | Corynebacterium | 11% | Brevibacterium | 10% | Bacillaceae | 5% |
| Arthrobacter | 2% | Alcaligenaceae | 11% | Candidate division TM7 | 9% | Enterococcus | 3% |
| Comamonadaceae | 2% | Nocardioidaceae | 4% | iii1-15 | 8% | Gaiellaceae | 3% |
| Acinetobacter | 2% | Unassigned | 3% | Corynebacterium | 4% | Brevibacterium | 3% |
| Candidate division TM7 | 1% | Streptomyces | 3% | Comamonadaceae | 3% | Comamonas | 3% |
| Gaiellaceae | 1% | Staphylococcus | 2% | Weeksellaceae | 3% | Staphylococcus | 2% |
| Pseudomonas | 1% | Cloacibacterium | 2% | Comamonas | 1% | Acidovorax | 2% |
| Isfjorden, 8 h | UNIS, 3 Day | ||||||
| OTU assignment | Relative abundance | OTU assignment | Relative abundance | ||||
| Oxalobacteraceae | 39% | Oxalobacteraceae | 22% | ||||
| Bacteroides | 10% | Alcanivorax | 14% | ||||
| iii1-15 | 6% | Burkholderiales | 7% | ||||
| Delftia | 5% | Corynebacterium | 6% | ||||
| Burkholderia | 4% | oc28 | 6% | ||||
| Achromobacter | 4% | Candidatus Aquiluna | 5% | ||||
| Caulobacteraceae | 3% | Bacillaceae | 5% | ||||
| Comamonadaceae | 3% | Microbacteriaceae | 5% | ||||
| Staphylococcaceae | 2% | Streptomyces | 4% | ||||
| Burkholderiales | 2% | iii1-15 | 4% | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuthbertson, L.; Amores-Arrocha, H.; Malard, L.A.; Els, N.; Sattler, B.; Pearce, D.A. Characterisation of Arctic Bacterial Communities in the Air above Svalbard. Biology 2017, 6, 29. https://doi.org/10.3390/biology6020029
Cuthbertson L, Amores-Arrocha H, Malard LA, Els N, Sattler B, Pearce DA. Characterisation of Arctic Bacterial Communities in the Air above Svalbard. Biology. 2017; 6(2):29. https://doi.org/10.3390/biology6020029
Chicago/Turabian StyleCuthbertson, Lewis, Herminia Amores-Arrocha, Lucie A. Malard, Nora Els, Birgit Sattler, and David A. Pearce. 2017. "Characterisation of Arctic Bacterial Communities in the Air above Svalbard" Biology 6, no. 2: 29. https://doi.org/10.3390/biology6020029
APA StyleCuthbertson, L., Amores-Arrocha, H., Malard, L. A., Els, N., Sattler, B., & Pearce, D. A. (2017). Characterisation of Arctic Bacterial Communities in the Air above Svalbard. Biology, 6(2), 29. https://doi.org/10.3390/biology6020029