1. Introduction
Thick filaments play a fundamental role in defining the sarcomeric structure and contractile properties of muscle. The major constituent of thick filaments in most muscle types is myosin II, a highly conserved protein characterized by a globular motor domain responsible for the chemomechanical basis of contraction, and a coiled-coil tail domain responsible for polymerization into highly ordered filaments [
1]. Additional species and muscle-type specific proteins confer thick filaments with distinct structural and functional characteristics tailored to the specific operational demands of the muscle as evidenced, for example, in the various forms of animal locomotion such as running, hopping, flying, swimming, and crawling [
2]. The predominant role of muscle in these various forms of locomotion often obscures its other important functions (e.g., heat production, sound production, metabolic health) that rely on contractile mechanisms that are operationally distinct from those used for locomotion. How a common thin and thick filament-based sarcomere design has evolved to fulfill different functional roles remains largely unknown.
Insect flight muscle engenders complex aerial behaviors involved in predation, mating rituals, and territorial protection, among others [
3,
4]. Insect flight muscle also facilitates ground-based behaviors involved in thermogenesis, ventilation, courtship, and sound generation, among others [
5,
6]. Studies in
Drosophila and other insects are beginning to shed light on the adaptive mechanisms by which muscle fulfills multiple roles [
7,
8]. For example, mutations in the
Drosophila myosin regulatory light chain manifest differently in flight and in the generation of the mating song, consistent with the hypothesis that the contractile based mechanisms underlying these two behaviors are different [
9].
Flightin is a 20 kDa (182 amino acids) myosin coiled-coil binding protein required for the normal assembly and function of thick filaments in
Drosophila flight muscles [
10,
11]. Thick filaments devoid of flightin assemble to abnormally long length, are structurally compromised, and are more compliant (
i.e., lower flexural rigidity) than their normal counterparts [
12,
13]. The absence of flightin renders the flight muscle inoperative due to a reduction in the viscoelastic response of fibers resulting in a significant loss of oscillatory work and power [
14]. Additionally, sarcomere structure is compromised to the point it cannot withstand contractile forces [
15]. Despite its critical role in flight muscle structure and function, little is known about the molecular properties of flightin and the mechanisms by which this important protein fulfills its functional roles.
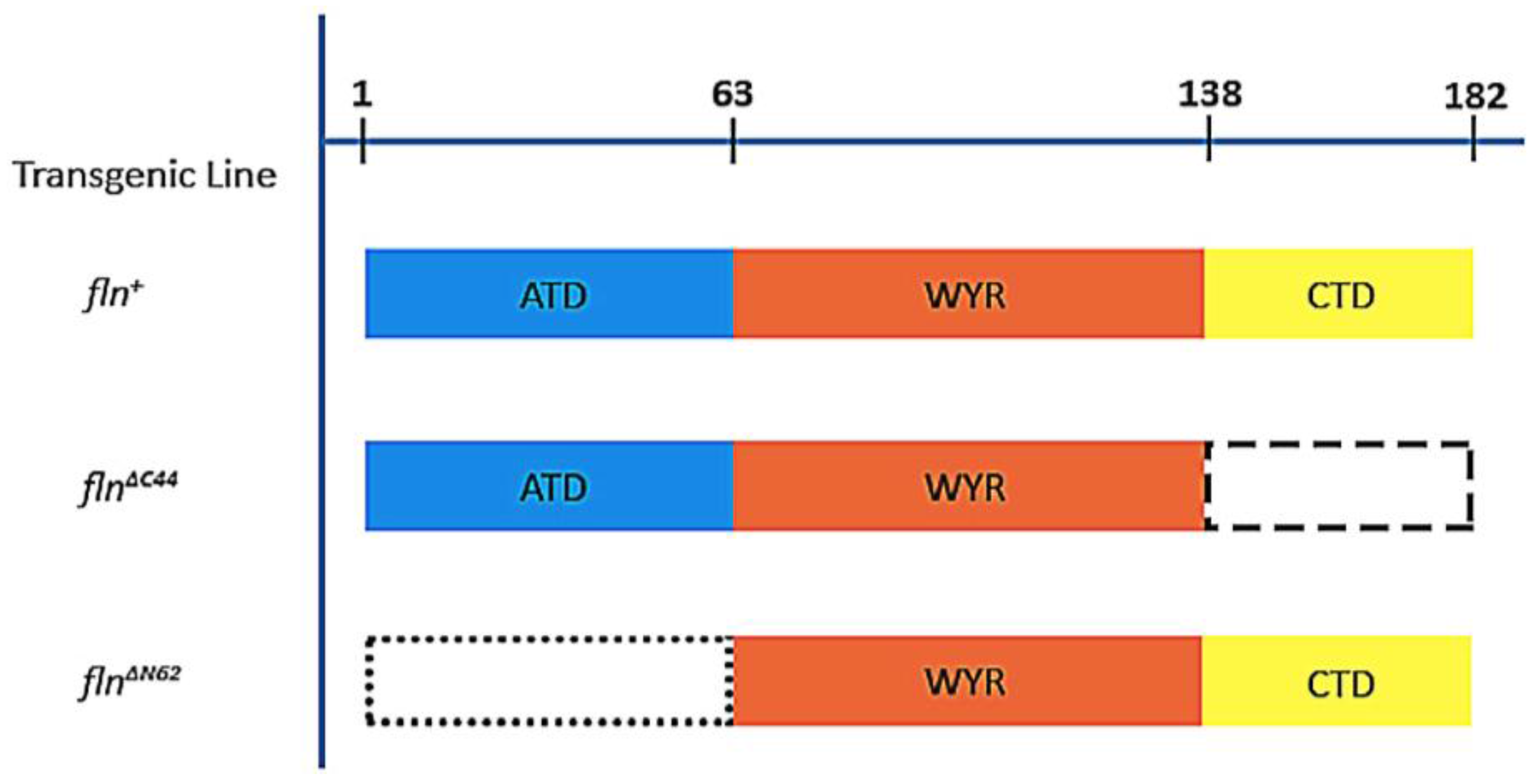
While flightin’s native structure is currently unknown, a comparison of its amino acid sequence amongst twelve
Drosophila species revealed a tripartite organization indicative of three separate domains [
16]. The region spanning the 84th to the 135th position (
D. melanogaster numbering), referred to as the WYR domain for its conserved tryptophan (W), tyrosines (Y) and arginine (R), shows a high degree (>90%) of amino acid sequence identity among
Drosophila and represents the only conserved flightin domain throughout the clade Pancrustacea [
17]. The amino terminal residues 1 through 65 are highly variable and present a low degree (<20%) of sequence conservation. Compared to these two regions, the carboxy terminal region (amino acid residues spanning positions 137 through 182) shows intermediate conservation (60% identity) [
16]. To establish if these regions that differ in amino acid sequence conservation represent independent protein domains, we created transgenic flies that express flightin devoid of the amino terminal region (
flnΔN62) and the carboxy terminal region (
flnΔC44) (
Figure 1) [
16,
18]. Both forms of truncated flightin are incorporated into thick filaments and partially restore flight muscle functionality in
fln0, a mutant strain that does not express flightin [
12]. The extent to which flight muscle function is restored in
flnΔC44 differs markedly from that in
flnΔN62, evidence in support of the hypothesis that the N-terminal region and the C-terminal region are protein domains with distinct functions.
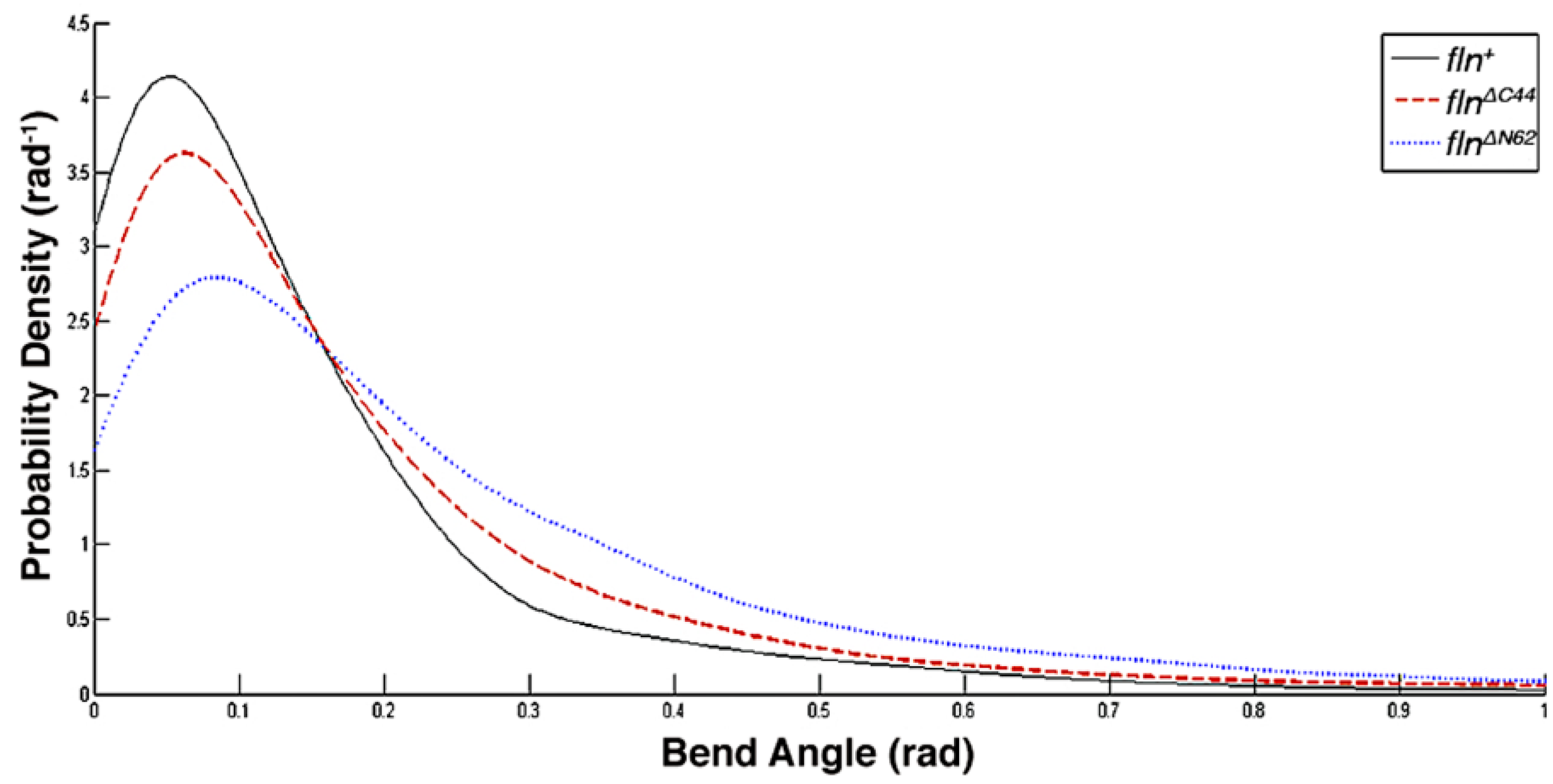
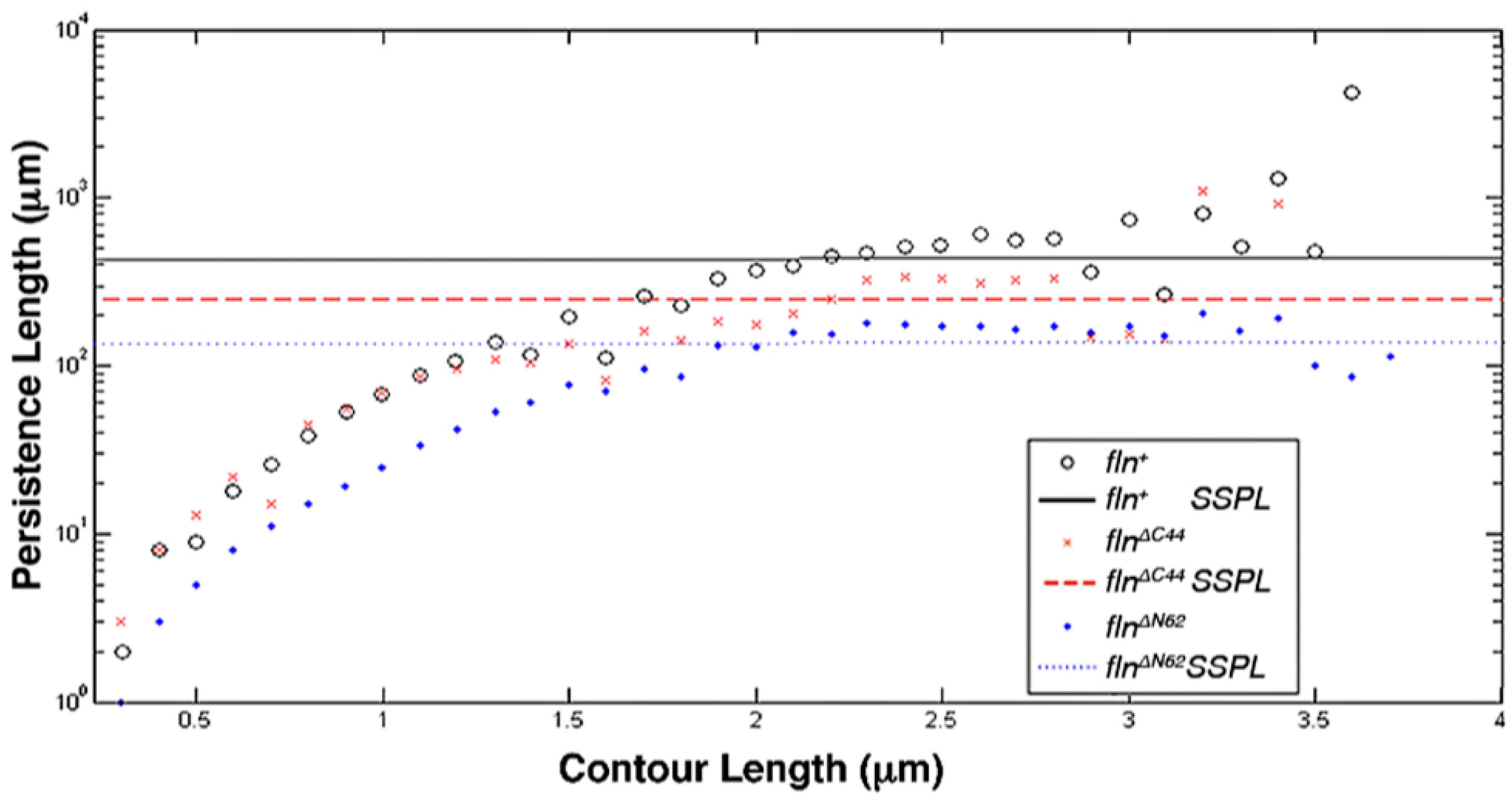
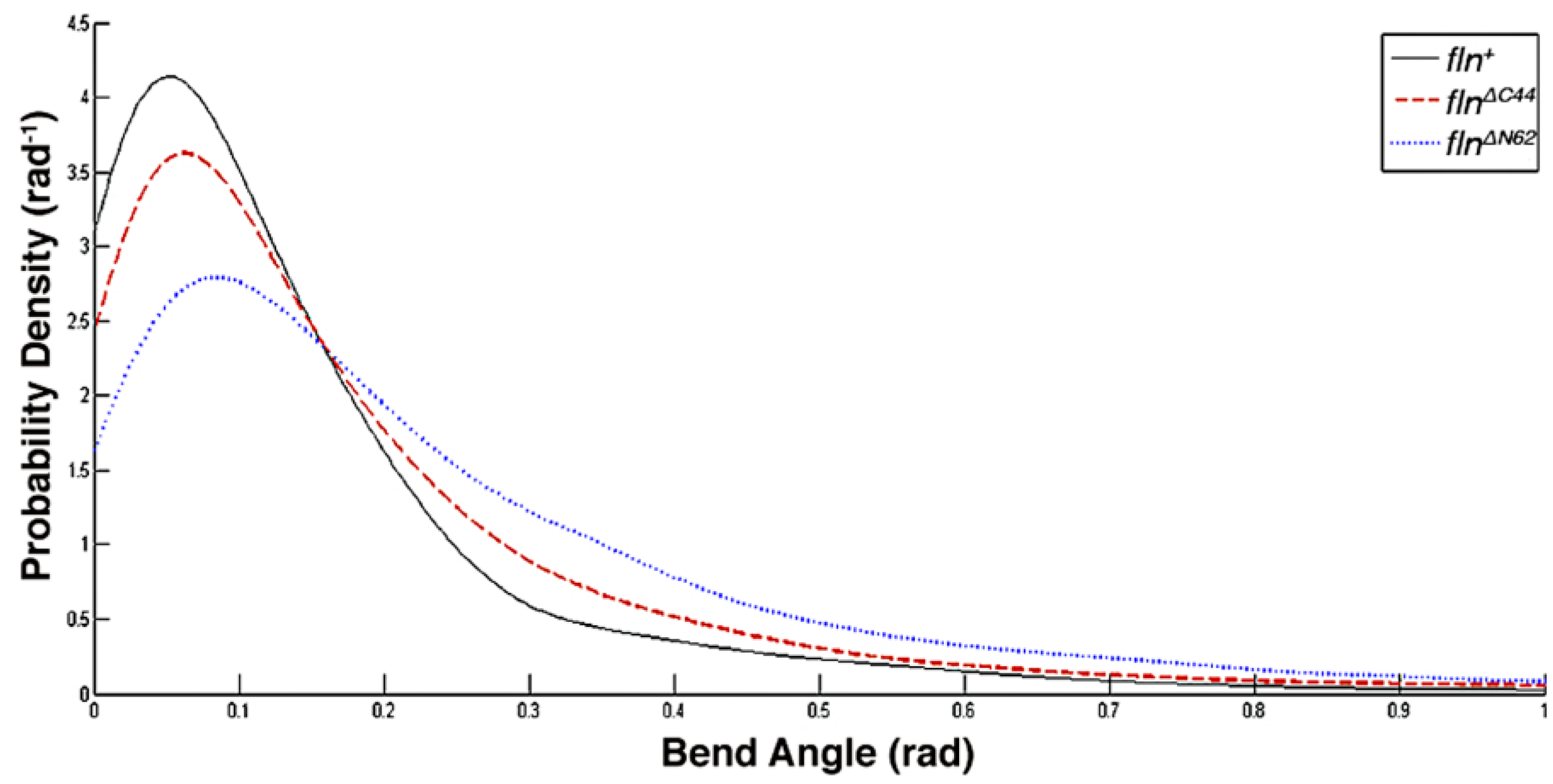
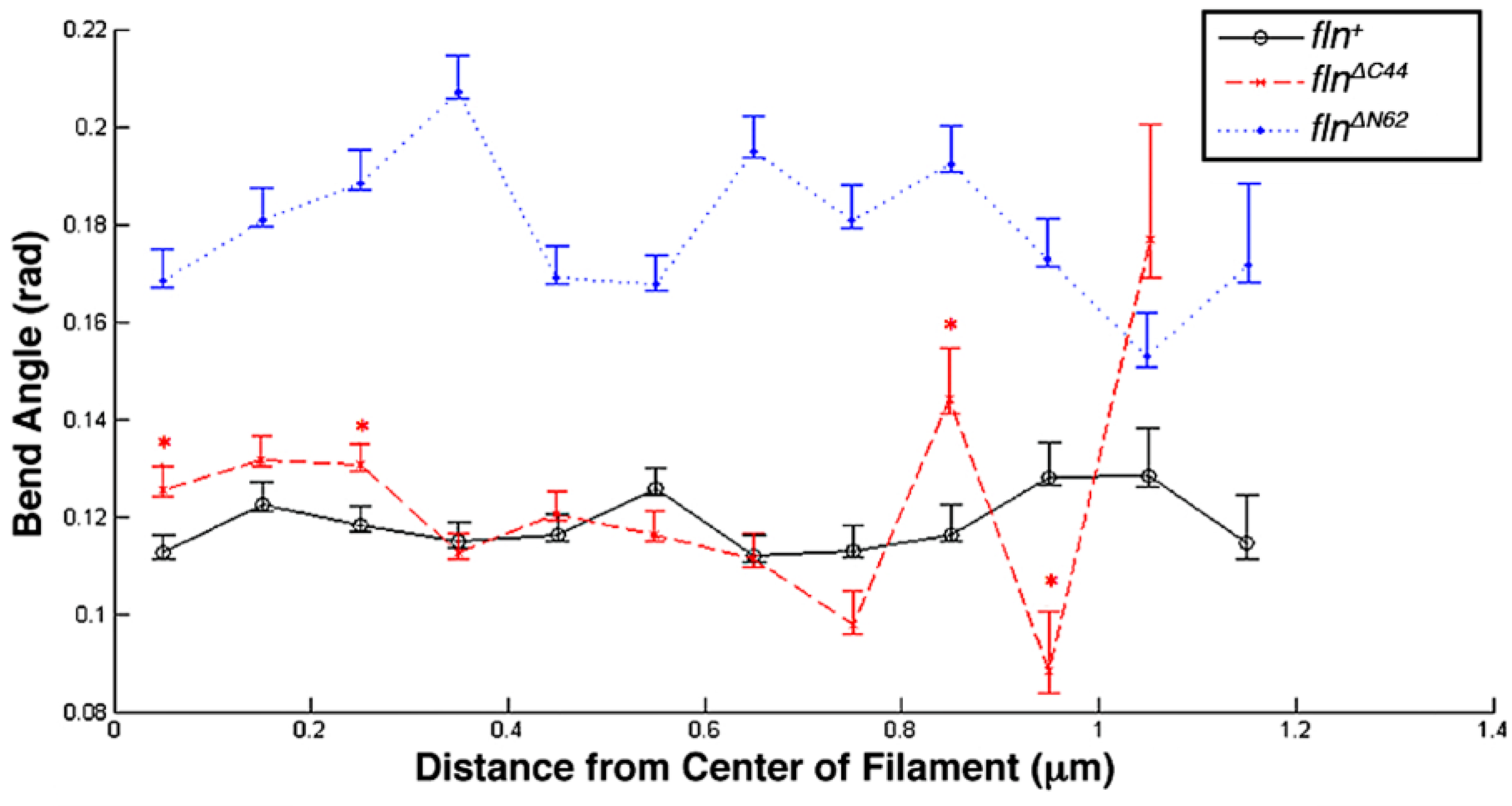
This study was conducted to examine how the flightin N-terminal domain and the C-terminal domain contribute to the biomechanical properties of thick filaments. Using atomic force microscopy (AFM), the length, bending propensity, and persistence length of isolated, native thick filaments from flnΔN62 and flnΔC44 flies were compared to fln+ control flies. Statistical polymer chain analysis revealed that each domain makes distinct contributions to thick filament biomechanics.
2. Materials and Methods
2.1. Drosophila Stocks
The generation of transgenic
fln+,
flnΔN62, and
flnΔC44 Drosophila melanogaster has been previously described [
16,
18,
19]. All fly stocks were raised in vials of yeast agar food at 22 °C, constant humidity, and on a 12:12 light:dark cycle. Vials were cleared of adults and newly eclosed females collected daily for dissection, ensuring that all flies were between one and three days old.
2.2. Solutions
Rigor solution (pCa 4.5) consisted of 200 mM ionic strength, pH 7.0: 149.2 mM sodium methanesulfonate, 5 mM ethylene glycol bis(β-aminoethyl ether)N,N′-tetraacetic acid (EGTA), 4.97 mM CaCl2, 1.18 mM MgCl2, 8 mM Pi, 20 mM Bes-7.0 (N,N-bis(2-hydroxyethyl)-2-aminoethanesulfonic acid), 7.4 mM KOH, 2 mM DTT, and 1% (v/v) Protease Inhibitor Cocktail Set III (Merck, Darmstadt, Germany). Relaxing solution (pCa 8.0) consisted of 200 mM ionic strength, pH 7.1: 97.6 mM sodium methanesulfonate, 15 mM EGTA, 55 μM CaCl2, 6.88 mM MgCl2, 5.39 mM ATP, 8 mM Pi, 20 mM Bes-7.0, 2.4 mM KOH, 20 mM BDM (2,3-butanedione monoxime), and 1% (v/v) Protease Inhibitor Cocktail Set III (Merck). Dissecting solution consisted of rigor solution with 2% (v/v) Triton X-100 and 50% (v/v) glycerol. Imaging Solution (pCa 4.5) consisted of 150 mM ionic strength, pH 7.0: 114 mM sodium methanesulfonate, 5 mM EGTA, 5 mM CaCl2, 1 mM MgCl2, 20 mM Bes-7.0, 1 mM KOH, 2 mM DTT, and 1% (v/v) Protease Inhibitor Cocktail Set III (Merck). Calpain Solution contained 2 mg/mL calpain-1 from porcine erythrocytes (Merck), 20 mM imidazole HCl, 5 mM β-mercaptoethanol, 1 mM EDTA, 1 mM EGTA, and 30% glycerol, pH 6.8.
With the exception of the calpain solution and the protease inhibitors (Merck KGaA: Darmstadt, Germany), all reagents were obtained from Sigma (St. Louis, MO, USA).
2.3. IFM Thick Filament Isolation
For each transgenic line of flies, 21–30 female flies aged one to three days old were anaesthetized with CO2 and transferred to a dissection plate. The head, wings, abdomen, and legs were removed, leaving thoraces that were then transferred into a dissection disk containing iced dissecting solution with fresh protease inhibitors. Thoraces were then split in half and transferred to a dish of fresh dissecting solution. This dish was covered and incubated overnight at −20 °C. IFM fibers were then dissected from the half thoraces and transferred to a chilled dish with fresh dissecting solution. Next, the isolated IFM fibers were transferred to a 1.5 mL centrifuge tube with rigor solution and sheared once with a 20 G needle, followed by centrifugation at 200 rpm and 4 °C for 4 min. The supernatant was decanted, and fresh rigor solution was added. This wash sequence was repeated two additional times with centrifugations at 400 rpm and 600 rpm. After the final wash, all but 15 μL of the rigor supernatant was removed and 20 μL of 2 mg/mL calpain solution was added. Calpain activation was achieved by adding 40 μL of 0.1 M CaCl2. This solution was placed on a rocking tray at 22 °C for 35 min followed by an additional 5 min at room temperature. Sixty μl of the calpain and CaCl2 solution added was removed from the fibers and digestion was then completely stopped by the addition of 400 μL of relaxing solution containing fresh protease inhibitors. The fibers were then sheared 7 times with a 1 mL syringe fitted with a 20 G needle, producing a transparent filament suspension. This suspension was then centrifuged for 4 min at 2000× g and the supernatant was separated from the pellet for AFM imaging.
2.4. Atomic Force Microscopy Imaging
An 80-μL thick filament sample was allowed to incubate on a freshly cleaved mica substrate for 10 min after which 70 μL of excess solution was removed. Approximately 70 μL of imaging solution lacking protease inhibitors was then added to the prepared mica disk. Images were produced using an MFP3D Bioscope AFM (Oxford Instruments/Asylum Research, Santa Barbara, CA, USA) in tapping mode. Images were recorded at scan sizes of 5 or 6 μm and 512 × 512 pixels, providing images with pixel resolutions of 9.8 and 11.7 nm, respectively. The AFM probes used were the Budget Sensors (Innovatie Solutions Bulgaria Ltd, Sofia, Bulgaria) SiNi-30 short, silicone nitride tips.
2.5. Thick Filament Analysis
Images generated by the AFM were analyzed as described in a prior study [
20]. Briefly, images were first processed in the proprietary format of the AFM via IgorPro (WaveMetrics Inc., Portland, OR, USA) where the images were flattened, planefit, and smoothed twice with a median filter. Images were then exported to the AFM’s accessory program ArgyleLite (Oxford Instruments/Asylum Research), where they were converted into ASCII matrix files that preserved pixel intensity from the AFM topography to be later analyzed in MatLab (MathWorks, Natick, MA, USA). Using a custom program written in Matlab and LabView, filaments were digitally rotated, aligned horizontally and isolated via image masking. These digitally isolated filaments were then fit with points placed perpendicular to the filament contour according to a Gaussian distribution to generate a digital filament consisting of a series of (x, y) coordinates. This provided a means of accurately measuring thick filament contour lengths (s). Along the individual contours, end-to-end length (R), and the angle between tangent vectors at the end of each contour (θ) were recorded. Using these measurements, thick filament persistence length was calculated on both a pooled and per-filament basis in order to retain information about the intrinsic variability in the stiffness of filaments from each sample. The individual filament persistence length, referred to as the Specific Persistence Length (SPL), was estimated according to Equation (1):
where (R
2(s)) is the mean squared end-to-end length and λ is the SPL. Group persistence length was calculated by averaging the individual persistence lengths at each contour and estimating the steady state value that was approached. Pixel limitation was determined by using completely straight digital “test filaments”, as described previously [
13].
2.6. Statistical Analysis
All values are reported as mean ± SE. Filament lengths were compared to the control, flightin rescued line (fln+) using a t-test. The Mann-Whitney U test, which operates on nonnormal distributions, was used to determine significance in the differences between SPL values and control. An ANCOVA multiple comparisons test allowed variations in contour length to be accounted for in the assessment of differences between steady state persistence lengths.
4. Discussion
The results presented here demonstrate that the amino terminal domain and the carboxy terminal domain of flightin make distinct contributions to the biomechanical properties of thick filaments, providing further insight into how flightin influences insect flight muscle functionality. Previously, we had shown that reintroduction of the full length flightin gene into a flightin null (
fln0) strain (
i.e., the
fln+ control strain used here) renews the structural integrity and function of the flight muscle to nearly wild-type levels [
19]. The extent of this phenotype rescue is further manifested here: the flightin rescue
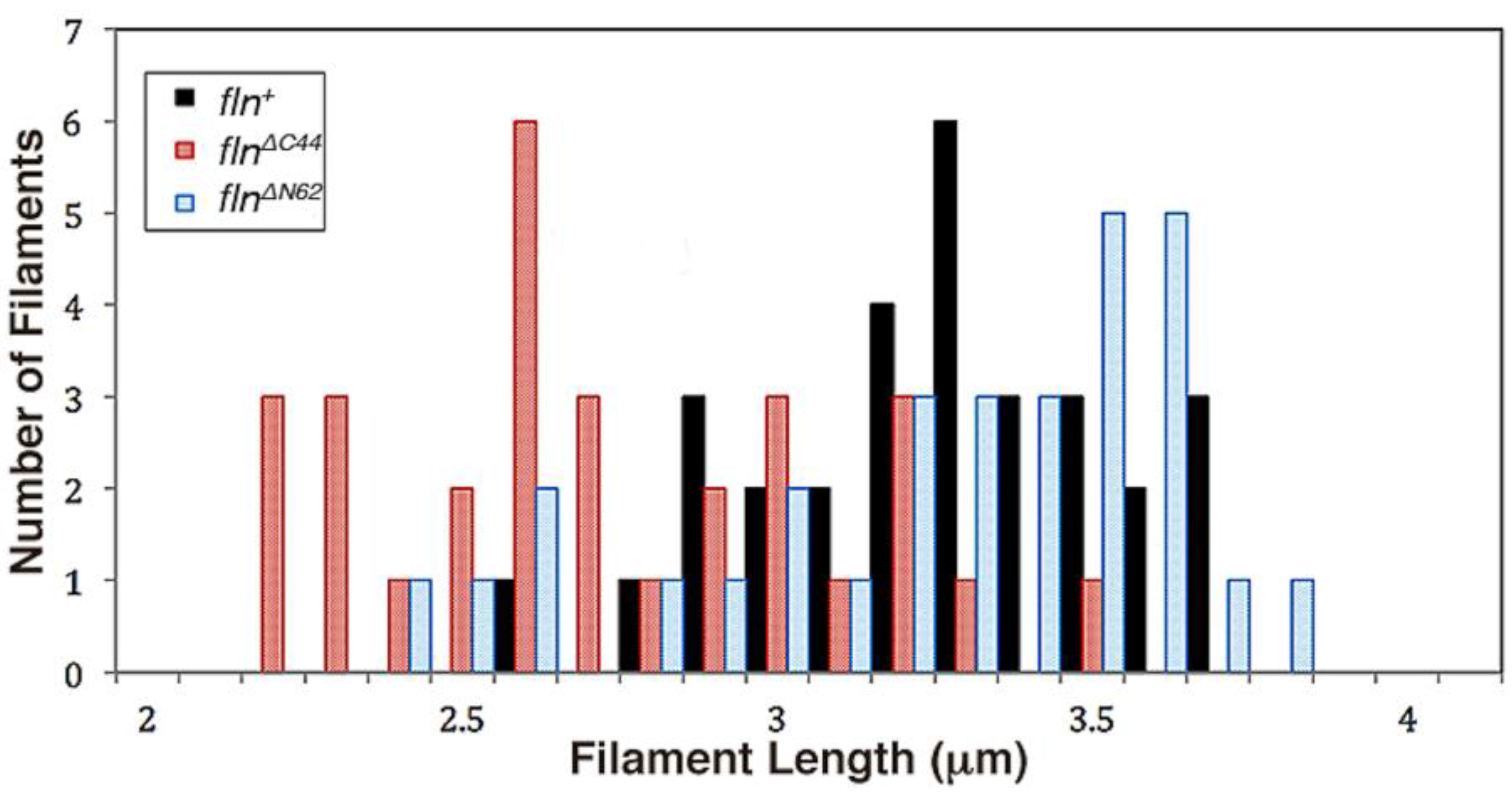
fln+ flies and the previously characterized wild type OR flies contain thick filaments of comparable lengths (3.21 ± 0.05 μm and 3.20 ± 0.04 μm) and specific persistence lengths (1386 ± 196 μm and 1742 ± 266 μm) [
13]. In contrast, the SSPL and mean bend angle values of thick filaments from
fln+ flies differed from those of OR flies reported previously [
13]. While this, in part, can be attributed to genetic differences between the wild-type OR strain and the transgenic
fln+ stock, the different imaging substrate used in this study (mica) and the prior study (HOPG) is also a contributing factor. For this study a transparent mica substrate was used to take advantage of the AFM’s inverse optical scope during imaging. The difference in transparency, however, is also accompanied by a change in surface charge distribution. In contrast to the nonpolar and hydrophobic surface of HOPG, mica has a negative charge. As the thick filament is also highly negative in charge, repulsive forces influence the typical Brownian motion of thick filaments and reduce equilibration onto the substrate, as was shown with intermediate filaments [
21]. In light of this, it is more appropriate to refer to the extrapolated filament stiffness indices as “apparent” persistence lengths, due to deviations in filament equilibration [
20].
Our results indicate the amino terminal domain exerts greater influence on filament stiffness than the carboxy terminal domain, while the latter has a greater impact on filament length. This division of labor is reflected in the amino acid sequence. The carboxy terminal domain sequence is more conserved than the amino terminal domain sequence, consistent with its role in defining a fundamental characteristic of thick filaments. While information on how the length of thick filaments determines muscle functionality is incomplete, it is evident from the remarkable consistency in filament length within a sarcomere that mechanisms involved in length determination are strongly selected for. A role in defining thick filament length provides an explanation for our prior observation that the carboxy terminal domain sequence is strongly conserved within a taxon, but decays rapidly between taxa [
17]. Among invertebrate muscles, thick filaments vary considerably in length (~1.5 to 50 μm), compared to their relatively narrow length range (~1.6 μm) in vertebrate skeletal muscle [
1,
22]. The exclusive presence of flightin in Pancrustacea, by far the most speciose clade, may afford these organisms greater latitude in tinkering with the common sarcomere design, and hence muscle functionality, through differences in thick filament length and architecture. A comparison of thick filament lengths between
flnΔC44 and
fln+ flies (2.68 ± 0.06 μm and 3.21 ± 0.05 μm, respectively) is imitative of the reduction in sarcomere length found between these transgenic strains (3.12 ± 0.02 μm for
flnΔC44 vs. 3.42 ± 0.04 μm for
fln+) [
16]. This suggests that factors influencing thick filament length may also impose constraints on higher-level sarcomere structure.
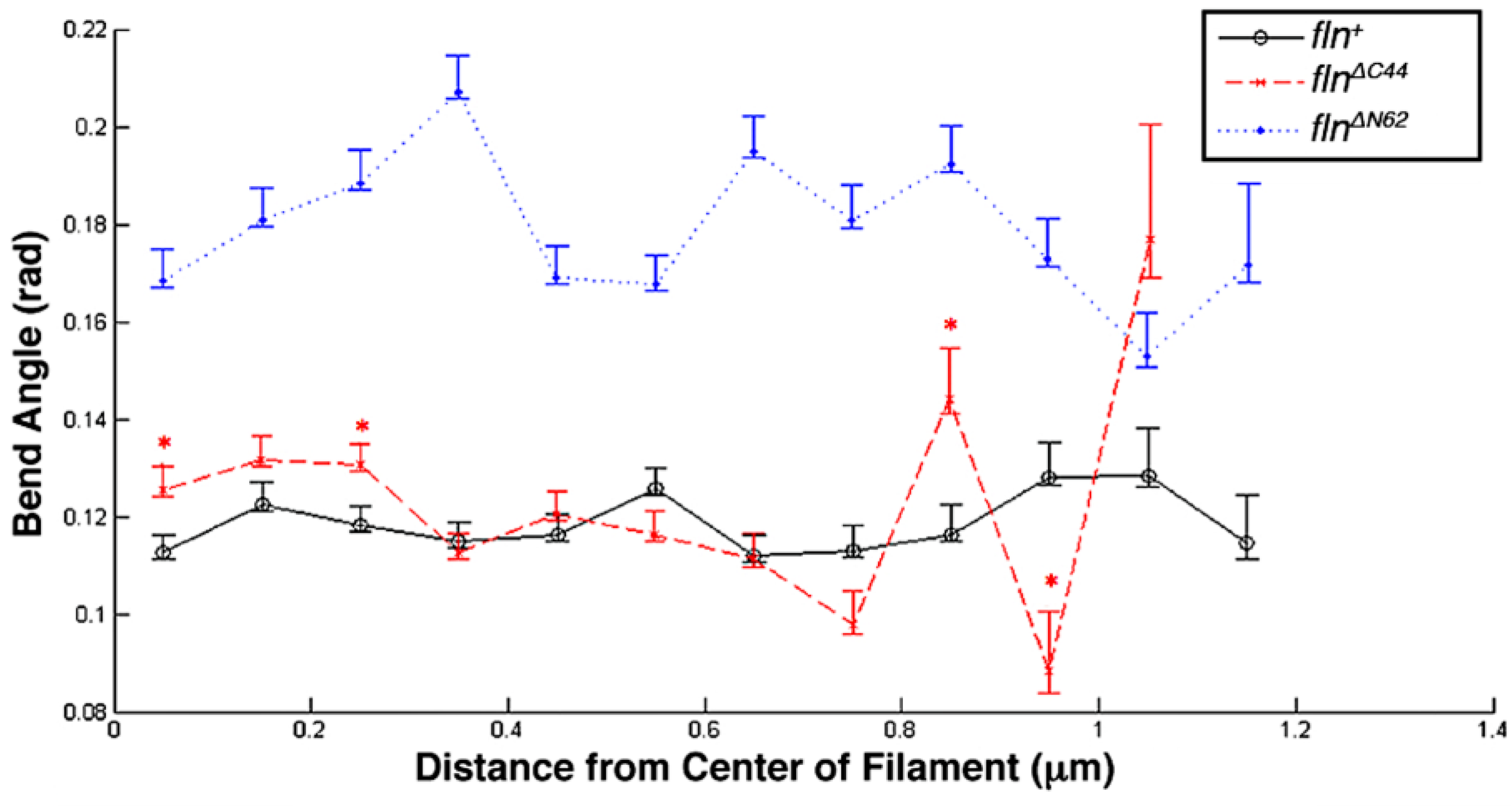
Of the three transgenic fly lines studied here, only
flnΔC44 flies are incapable of beating their wings [
16]. Transitively, the results presented here showing a significant decrease in filament length and, to an extent, flexural rigidity provides a potential mechanism for the reduced power output observed in
flnΔC44, as shorter filaments have fewer molecular motors. This is consistent with the reduction observed in modeled parameters B and C, indicative of a decrease in the number of strongly bound cross-bridges during active contraction. Additionally, deletion of the carboxy terminal domain affects myofilament lattice organization and decreases cross-bridge cycling kinetics by reducing the rate of cross-bridge recruitment [
16]. Furthermore, sarcomeres in
flnΔC44 flies have characteristic defects in M-line structure, suggesting that the carboxy terminal domain may facilitate a stabilizing interaction with an M-line protein such as miniparamyosin [
23]. As miniparamyosin is also distributed at the thick filament tips, an interaction with the flightin carboxy terminal domain may also explain the greater variation in bend angle profile at the terminal ends of
flnΔC44 thick filaments [
23]. In terms of SPL, there was no significant difference between the thick filaments found in the
fln+ and
flnΔC44 flies. This is reflected in the mechanical performance of single skinned fibers as there is a consistency between the elastic modulus of
fln+ and
flnΔC44 fibers under relaxing conditions [
16]. In summary, the direct effect of the carboxy terminal domain on thick filament length may underlie the myofilament lattice disorder and compromised contractile kinetics with consequent loss of flight in
flnΔC44 flies.
The amino terminal domain is the least conserved region in flightin. The marked influence of this region on filament stiffness provides a possible explanation for its lack of sequence conservation. Thick filament stiffness influences fiber stiffness, which in turn defines muscle functional outcomes [
22]. Altering the amino acid sequence of the N-terminal domain may be a mechanism for fine-tuning filament stiffness without compromising basic contractile function. This is evident inasmuch as
flnΔN62 flies are flight competent and have normal wing beat frequency [
18]. The flight muscle is responsible for generating the wing beats required for flight and for production of the mating song, two key behaviors with distinct muscle power requirements and subject to different evolutionary constraints. Among Drosophilids, mating songs are species-specific and have evolved in response to sexual selection and contributed to speciation [
24,
25]. As genes under sexual selection tend to evolve at a faster rate, we speculate that the amino terminal domain of flightin contributes to species-specific song attributes via its role in thick filament stiffness.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}