
Heterochrony as Diachronically Modified Cell-Cell Interactions

Abstract
:1. Introduction

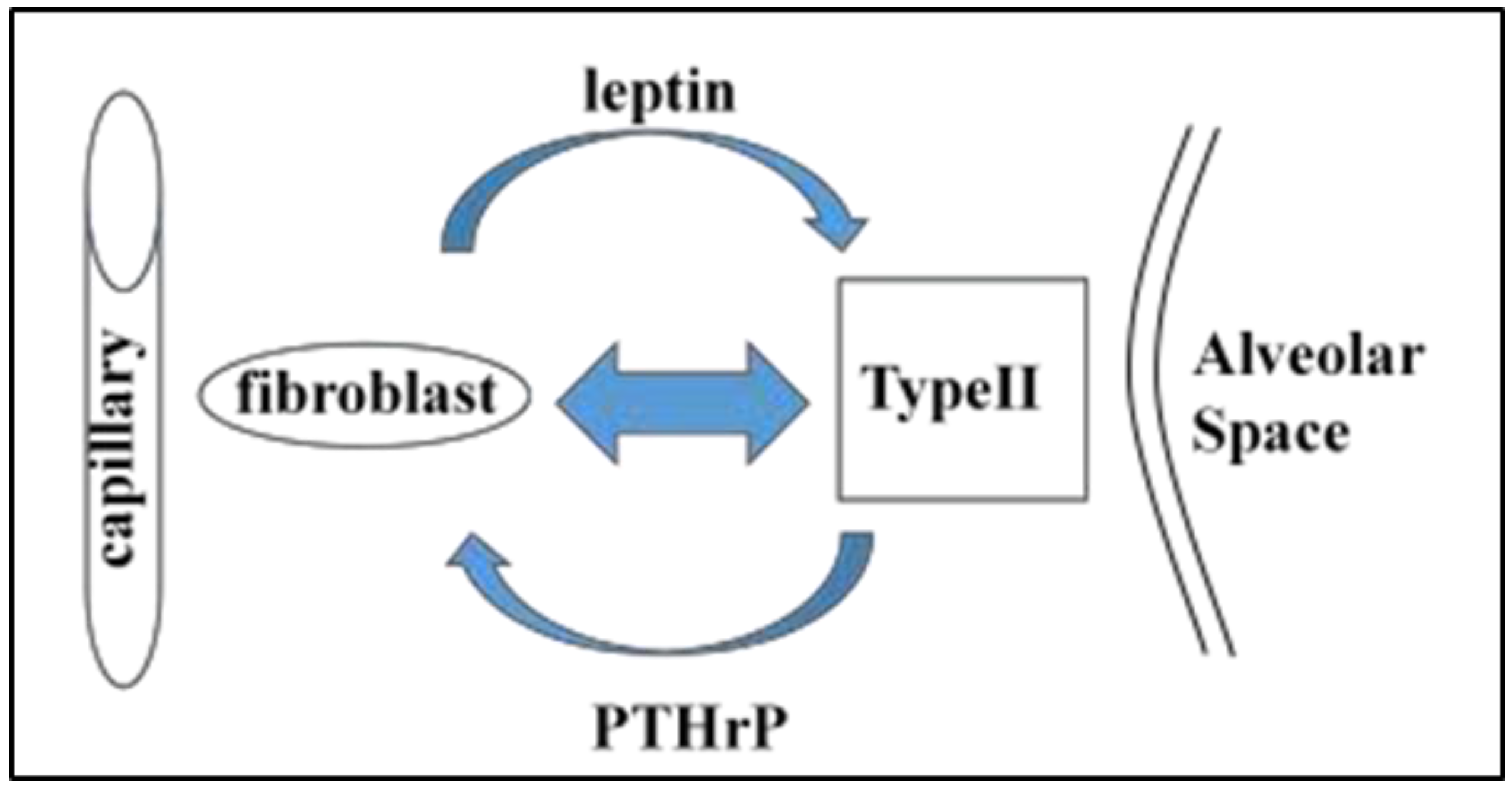{kind=link}
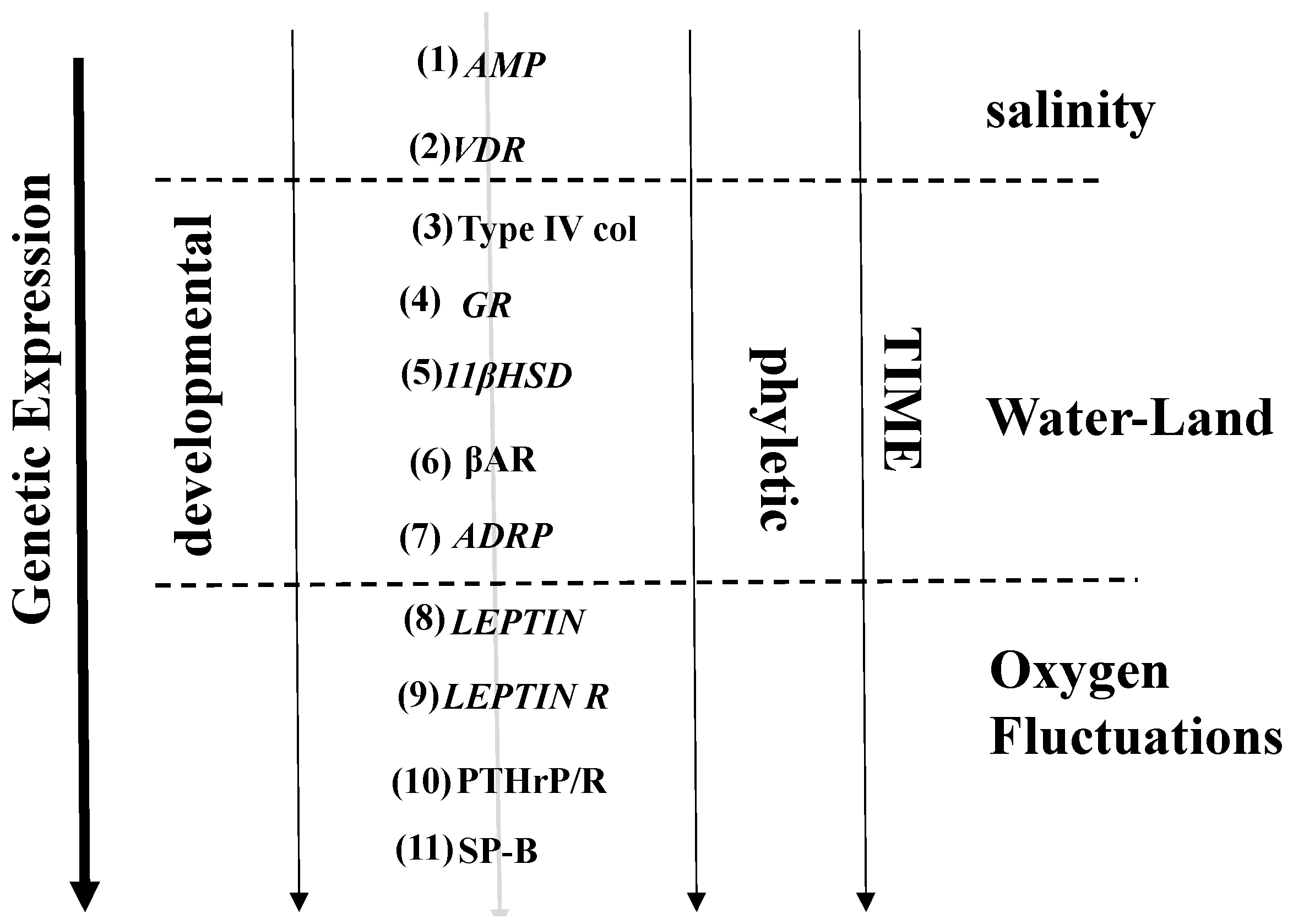{kind=link}
|
|
|
|
|
|
|
|
|
|
2. Normal Embryologic Development, or “Monochrony”, in Contrast to Heterochrony
3. Lung Evolution as Ontogeny and Phylogeny
4. The Lipofibroblast as a “Rosetta Stone” for Lung Evolution


5. Physical Stress and Heterochrony—The Role of Gravity
6. Physiologic Stress—The Role of Hypoxia
7. Chronic Lung Disease as “Reverse” Heterochrony
8. Goodpasture’s Syndrome as Waterproofing
9. Conclusions
Acknowledgments
Conflicts of Interest
References
- Gilbert, S.F. Ernst haeckel and the biogenetic law. In Developmental Biology; Sinauer Associates: Sunderland, MA, USA, 2006. [Google Scholar]
- Gould, S.J. Ontogeny and Phylogeny; Harvard University Press: Cambridge, MA, USA, 1977. [Google Scholar]
- Smith, K.K. Time’s arrow: Heterochrony and the evolution of development. Int. J. Dev. Biol. 2003, 47, 613–621. [Google Scholar] [PubMed]
- Gilbert, S.F.; Opitz, J.M.; Raff, R.A. Resynthesizing evolutionary and developmental biology. Dev. Biol. 1996, 173, 357–372. [Google Scholar] [CrossRef] [PubMed]
- De Robertis, E.M.; Larraín, J.; Oelgeschläger, M.; Wessely, O. The establishment of Spemann’s organizer and patterning of the vertebrate embryo. Nat. Rev. Genet. 2000, 1, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Danesh, S.M.; Villasenor, A.; Chong, D.; Soukup, C.; Cleaver, O. BMP and BMP receptor expression during murine organogenesis. Gene Expr. Patterns 2009, 9, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Croce, J.C.; McClay, D.R. Evolution of the Wnt pathways. Methods Mol. Biol. 2008, 469, 3–18. [Google Scholar] [PubMed]
- Torday, J.S.; Rehan, V.K. Evolutionary Biology, Cell-Cell Communication and Complex Disease; Wiley: Hoboken, NJ, USA, 2012. [Google Scholar]
- Torday, J.S. Evolutionary biology redux. Perspect. Biol. Med. 2013, 56, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. A central theory of biology. Med. Hypotheses 2015, 85, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Smocovitis, V.B. Unifying Biology; Princeton University Press: Princeton, NJ, USA, 1996. [Google Scholar]
- Gould, S.J. The Structure of Evolutionary Theory; Harvard University Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Thisse, B.; Thisse, C. Formation of the vertebrate embryo: Moving beyond the Spemann organizer. Semin. Cell Dev. Biol. 2015, 42, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, T.S. The Structure of Scientific Revolutions; University of Chicago Press: Chicago, IL, USA, 1962. [Google Scholar]
- Saiz, N.; Plusa, B. Early cell fate decisions in the mouse embryo. Reproduction 2013, 145, R65–R80. [Google Scholar] [CrossRef] [PubMed]
- Hubaud, A.; Pourquié, O. Signalling dynamics in vertebrate segmentation. Nat. Rev. Mol. Cell Biol. 2014, 15, 709–721. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, C.; Krivega, M.; Cauffman, G.; Geens, M.; van de Velde, H. Totipotency and lineage segregation in the human embryo. Mol. Hum. Reprod. 2014, 20, 599–618. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Forhead, A.J. Glucocorticoids as regulatory signals during intrauterine development. Exp. Physiol. 2015, 100, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.A.; Larson, E.L.; Harrison, R.G. Hybrid zones: Windows on climate change. Trends Ecol. Evol. 2015, 30, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Wollstein, A.; Stephan, W. Inferring positive selection in humans from genomic data. Investig. Genet. 2015. [Google Scholar] [CrossRef] [PubMed]
- Shubin, N.H.; Daeschler, E.B.; Jenkins, F.A., Jr. Pelvic girdle and fin of Tiktaalik roseae. Proc. Natl. Acad. Sci. USA 2014, 111, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.J. Losing history: How extinctions prune features from the tree of life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.; Labandeira, C.; Laurin, M.; Berner, R.A. Confirmation of Romer’s Gap as a low oxygen interval constraining the timing of initial arthropod and vertebrate terrestrialization. Proc. Natl. Acad. Sci. USA 2006, 103, 16818–16822. [Google Scholar] [CrossRef] [PubMed]
- Romer, A.S. The Vertebrate Story; University of Chicago Press: Chicago, IL, USA, 1949. [Google Scholar]
- Pinheiro, P.L.; Cardoso, J.C.; Power, D.M.; Canário, A.V. Functional characterization and evolution of PTH/PTHrP receptors: Insights from the chicken. BMC Evol. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Verheijen, M.H.; Defize, L.H. Signals governing extraembryonic endoderm formation in the mouse: Involvement of the type 1 parathyroid hormone-related peptide (PTHrP) receptor, p21Ras and cell adhesion molecules. Int. J. Dev. Biol. 1999, 43, 711–721. [Google Scholar] [PubMed]
- Rubin, L.P.; Kifor, O.; Hua, J.; Brown, E.M.; Torday, J.S. Parathyroid hormone (PTH) and PTH-related protein stimulate surfactant phospholipid synthesis in rat fetal lung, apparently by a mesenchymal-epithelial mechanism. Biochim. Biophys. Acta 1994, 1223, 91–100. [Google Scholar] [CrossRef]
- Rubin, L.P.; Kovacs, C.S.; de Paepe, M.E.; Tsai, S.W.; Torday, J.S.; Kronenberg, H.M. Arrested pulmonary alveolar cytodifferentiation and defective surfactant synthesis in mice missing the gene for parathyroid hormone-related protein. Dev. Dyn. 2004, 230, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Hochane, M.; Raison, D.; Coquard, C.; Imhoff, O.; Massfelder, T.; Moulin, B.; Helwig, J.J.; Barthelmebs, M. Parathyroid hormone-related protein is a mitogenic and a survival factor of mesangial cells from male mice: Role of intracrine and paracrine pathways. Endocrinology 2013, 154, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. Parathyroid hormone-related protein is a gravisensor in lung and bone cell biology. Adv. Space Res. 2003, 32, 1569–1576. [Google Scholar] [CrossRef]
- Foley, J.; Longely, B.J.; Wysolmerski, J.J.; Dreyer, B.E.; Broadus, A.E.; Philbrick, W.M. PTHrP regulates epidermal differentiation in adult mice. J. Investig. Dermatol. 1998, 111, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Holland, N.D. Early central nervous system evolution: An era of skin brains? Nat. Rev. Neurosci. 2003, 4, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, Z.; Collins, J.E.; Andrews, R.M.; Stemple, D.; Gong, Z. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung. PLoS ONE 2011, 6, e24019. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.B.; Orgeig, S.; Sullivan, L.C.; Ling, N.; Bennett, M.B.; Schürch, S.; Val, A.L.; Brauner, C.J. The origin and evolution of the surfactant system in fish: Insights into the evolution of lungs and swim bladders. Physiol. Biochem. Zool. 2004, 77, 732–749. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.C.; Daniels, C.B.; Phillips, I.D.; Orgeig, S.; Whitsett, J.A. Conservation of surfactant protein A: Evidence for a single origin for vertebrate pulmonary surfactant. J. Mol. Evol. 1998, 46, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. The evolutionary continuum from lung development to homeostasis and repair. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L608–L611. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. The evolution of cell communication: The road not taken. Cell Commun. Insights 2009, 2, 17–25. [Google Scholar] [PubMed]
- Torday, J.S.; Rehan, V.K. On the evolution of the Lipofibroblast. Exp. Cell Res. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
- De Duve, C. Evolution of the peroxisome. Ann. N. Y. Acad. Sci. 1969, 168, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Developmental cell/molecular biologic approach to the etiology and treatment of bronchopulmonary dysplasia. Pediatr. Res. 2007, 62, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Torday, D.P.; Gutnick, J.; Qin, J.; Rehan, V. Biologic role of fetal lung fibroblast triglycerides as antioxidants. Pediatr. Res. 2001, 49, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.J.; Torres, E.; Londos, C.; Torday, J.S. Role of adipocyte differentiation-related protein in surfactant phospholipid synthesis by type II cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 283, L288–L296. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Sun, H.; Qin, J. Prostaglandin E2 integrates the effects of fluid distension and glucocorticoid on lung maturation. Am. J. Physiol. 1998, 274, L106–L111. [Google Scholar] [PubMed]
- Torday, J.S.; Sun, H.; Wang, L.; Torres, E.; Sunday, M.E.; Rubin, L.P. Leptin mediates the parathyroid hormone-related protein paracrine stimulation of fetal lung maturation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 282, L405–L410. [Google Scholar] [PubMed]
- O’Hare, K.H.; Sheridan, M.N. Electron microscopic observations on the morphogenesis of the albino rat lung, with special reference to pulmonary epithelial cells. Am. J. Anat. 1970, 127, 181–206. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, C.; Brody, J.S. Ultrastructure of developing alveoli. I. The role of the interstitial fibroblast. Anat. Rec. 1978, 192, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Maksvytis, H.J.; Niles, R.M.; Simanovsky, L.; Minassian, I.A.; Richardson, L.L.; Hamosh, M.; Hamosh, P.; Brody, J.S. In vitro characteristics of the lipid-filled interstitial cell associated with postnatal lung growth: Evidence for fibroblast heterogeneity. J. Cell Physiol. 1984, 118, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, N.B.; Grant, M.M.; Brody, J.S. The lipid interstitial cell of the pulmonary alveolus. Age and species differences. Am. Rev. Respir. Dis. 1985, 132, 1307–1312. [Google Scholar] [PubMed]
- Rehan, V.K.; Sugano, S.; Wang, Y.; Santos, J.; Romero, S.; Dasgupta, C.; Keane, M.P.; Stahlman, M.T.; Torday, J.S. Evidence for the presence of lipofibroblasts in human lung. Exp. Lung Res. 2006, 32, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Frank, L.; Bucher, J.R.; Roberts, R.J. Oxygen toxicity in neonatal and adult animals of various species. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1978, 45, 699–704. [Google Scholar] [PubMed]
- Mostello, D.J.; Hamosh, M.; Hamosh, P. Effect of dexamethasone on lipoprotein lipase activity of fetal rat lung. Biol. Neonate 1981, 40, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.; Hua, J.; Slavin, R. Metabolism and fate of neutral lipids of fetal lung fibroblast origin. Biochim. Biophys. Acta 1995, 1254, 198–206. [Google Scholar] [CrossRef]
- Londos, C.; Sztalryd, C.; Tansey, J.T.; Kimmel, A.R. Role of PAT proteins in lipid metabolism. Biochimie 2005, 87, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.; Rehan, V. Neutral lipid trafficking regulates alveolar type II cell surfactant phospholipid and surfactant protein expression. Exp. Lung Res. 2011, 37, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Sanchez-Esteban, J.; Rubin, L.P. Paracrine mediators of mechanotransduction in lung development. Am. J. Med. Sci. 1998, 316, 205–208. [Google Scholar] [PubMed]
- Sanchez-Esteban, J.; Cicchiello, L.A.; Wang, Y.; Tsai, S.W.; Williams, L.K.; Torday, J.S.; Rubin, L.P. Mechanical stretch promotes alveolar epithelial type II cell differentiation. J. Appl. Physiol. 2001, 91, 589–595. [Google Scholar] [PubMed]
- Torday, J.S.; Rehan, V.K. Up-regulation of fetal rat lung parathyroid hormone-related protein gene regulatory network down-regulates the Sonic Hedgehog/Wnt/betacatenin gene regulatory network. Pediatr. Res. 2006, 60, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Purevdorj-Gage, B.; Sheehan, K.B.; Hyman, L.E. Effects of low-shear modeled microgravity on cell function, gene expression, and phenotype in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2006, 72, 4569–4575. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Wandinger-Ness, A.; Roitbak, T. Altered trafficking and epithelial cell polarity in disease. Trends Cell Biol. 2002, 12, 374–381. [Google Scholar] [CrossRef]
- Knop, M. Yeast cell morphology and sexual reproduction—A short overview and some considerations. C. R. Biol. 2011, 334, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.F.; Carrier, D.R. The coupled evolution of breathing and locomotion as a game of leapfrog. Physiol. Biochem. Zool. 2006, 79, 997–999. [Google Scholar] [CrossRef] [PubMed]
- Payne, J.L.; Boyer, A.G.; Brown, J.H.; Finnegan, S.; Kowalewski, M.; Krause, R.A., Jr.; Lyons, S.K.; McClain, C.R.; McShea, D.W.; Novack-Gottshall, P.M.; et al. Two-phase increase in the maximum size of life over 3.5 billion years reflects biological innovation and environmental opportunity. Proc. Nat. Acad. Sci. USA 2009, 106, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Berner, R.A.; Vandenbrooks, J.M.; Ward, P.D. Evolution. Oxyg. Evol. Sci. 2007, 316, 557–558. [Google Scholar]
- Graham, J.B.; Dudley, R.; Aguilar, N.M.; Gans, C. Implications of the later Palaeozoic oxygen pulse for physiology and evolution. Nature 1995, 375, 117–120. [Google Scholar] [CrossRef]
- Mamillapalli, R.; Wysolmerski, J. The calcium-sensing receptor couples to Galpha(s) and regulates PTHrP and ACTH secretion in pituitary cells. J. Endocrinol. 2010, 204, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Suri, L.N.; McCaig, L.; Picardi, M.V.; Ospina, O.L.; Veldhuizen, R.A.; Staples, J.F.; Possmayer, F.; Yao, L.J.; Perez-Gil, J.; Orgeig, S. Adaptation to low body temperature influences pulmonary surfactant composition thereby increasing fluidity while maintaining appropriately ordered membrane structure and surface activity. Biochim. Biophys. Acta 2012, 1818, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Perlman, R.L.; Chalfie, M. Catecholamine release from the adrenal medulla. Clin. Endocrinol. Metab. 1977, 6, 551–576. [Google Scholar] [CrossRef]
- Maina, J.N. Structure, function and evolution of the gas exchangers: Comparative perspectives. J Anat. 2002, 201, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.B.; Lopatko, O.V.; Orgeig, S. Evolution of surface activity related functions of vertebrate pulmonary surfactant. Clin. Exp. Pharmacol. Physiol. 1998, 25, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Lau, M.J.; Keough, K.M. Lipid composition of lung and lung lavage fluid from map turtles (Malaclemys geographica) maintained at different environmental temperatures. Can. J. Biochem. 1981, 59, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Lopatko, O.V.; Orgeig, S.; Palmer, D.; Schürch, S.; Daniels, C.B. Alterations in pulmonary surfactant after rapid arousal from torpor in the marsupial Sminthopsis crassicaudata. J. Appl. Physiol. 1999, 86, 1959–1970. [Google Scholar] [PubMed]
- Voelkel, N.F.; MacNee, W. Chronic Obstructive Lung Diseases 2; BC Decker Inc.: Hamilton, ON, Canada, 2008. [Google Scholar]
- McGowan, S.E.; Torday, J.S. The pulmonary lipofibroblast (lipid interstitial cell) and its contributions to alveolar development. Annu. Rev. Physiol. 1997, 59, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Cerny, L.; Torday, J.S.; Rehan, V.K. Prevention and treatment of bronchopulmonary dysplasia: Contemporary status and future outlook. Lung 2008, 186, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Csete, M.; Walikonis, J.; Slawny, N.; Wei, Y.; Korsnes, S.; Doyle, J.C.; Wold, B. Oxygen-mediated regulation of skeletal muscle satellite cell proliferation and adipogenesis in culture. J. Cell Physiol. 2001, 189, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Regulation of adipocyte gene expression and differentiation by peroxisome proliferator activated receptor gamma. Curr. Opin. Genet. Dev. 1995, 5, 571–576. [Google Scholar] [CrossRef]
- Case, R.M.; Eisner, D.; Gurney, A.; Jones, O.; Muallem, S.; Verkhratsky, A. Evolution of calcium homeostasis: From birth of the first cell to an omnipresent signaling system. Cell Calcium 2007, 42, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F. Evolution and tinkering. Science 1977, 196, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.W. Polycyclic aromatic hydrocarbons: Primitive pigment systems in the prebiotic environment. Adv. Space Res. 1992, 12, 183–189. [Google Scholar] [CrossRef]
- Roux, E. The concept of function in modern physiology. J. Physiol. 2014, 592, 2245–2249. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torday, J.S. Heterochrony as Diachronically Modified Cell-Cell Interactions. Biology 2016, 5, 4. https://doi.org/10.3390/biology5010004
Torday JS. Heterochrony as Diachronically Modified Cell-Cell Interactions. Biology. 2016; 5(1):4. https://doi.org/10.3390/biology5010004
Chicago/Turabian StyleTorday, John S. 2016. "Heterochrony as Diachronically Modified Cell-Cell Interactions" Biology 5, no. 1: 4. https://doi.org/10.3390/biology5010004
APA StyleTorday, J. S. (2016). Heterochrony as Diachronically Modified Cell-Cell Interactions. Biology, 5(1), 4. https://doi.org/10.3390/biology5010004