

Environmental Drivers of Phytoplankton Structure in a Semi-Arid Reservoir

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
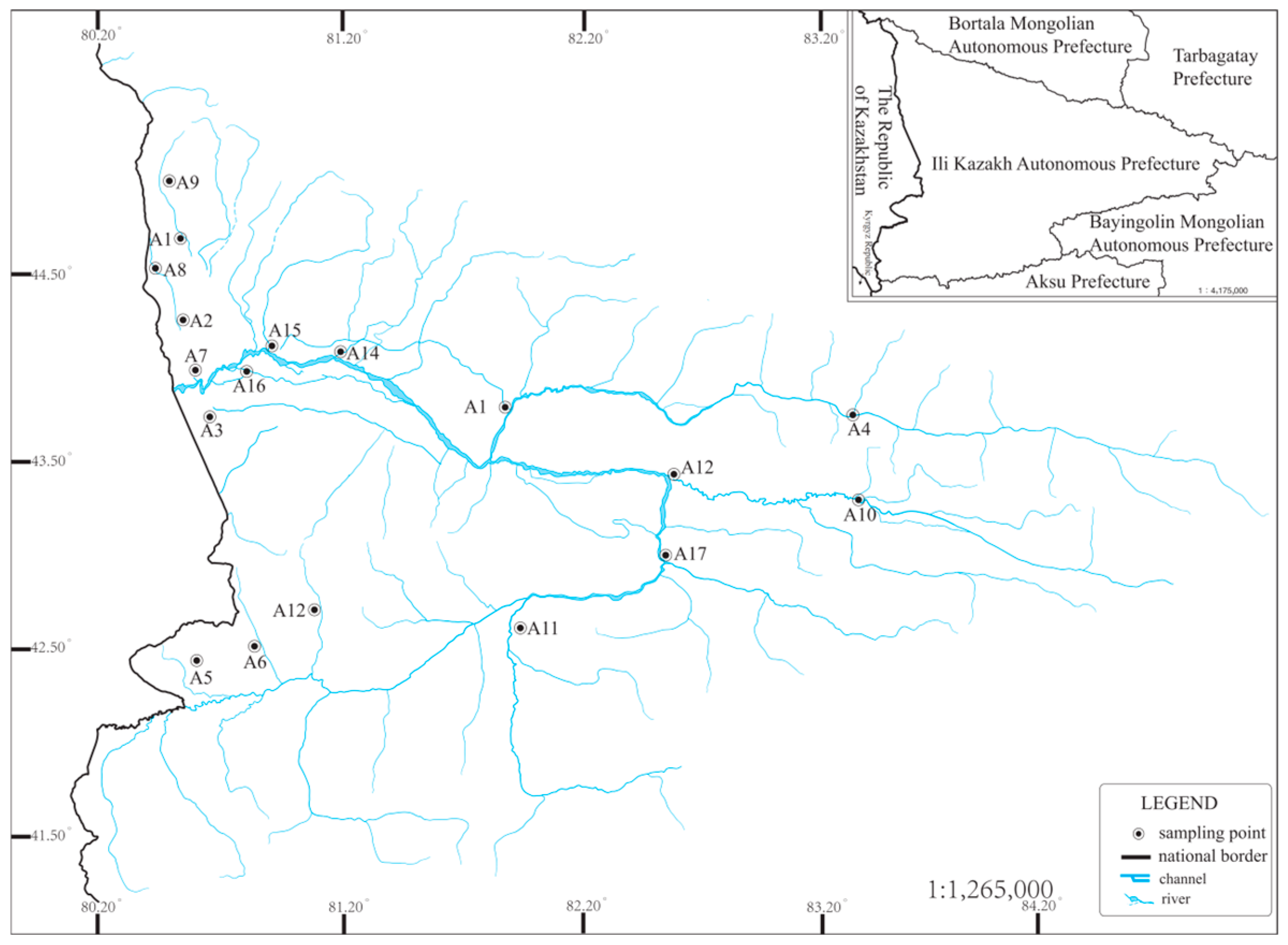
2.1. Study Area
2.2. Field Sampling and Data Acquisition
2.3. Data Analysis
3. Results
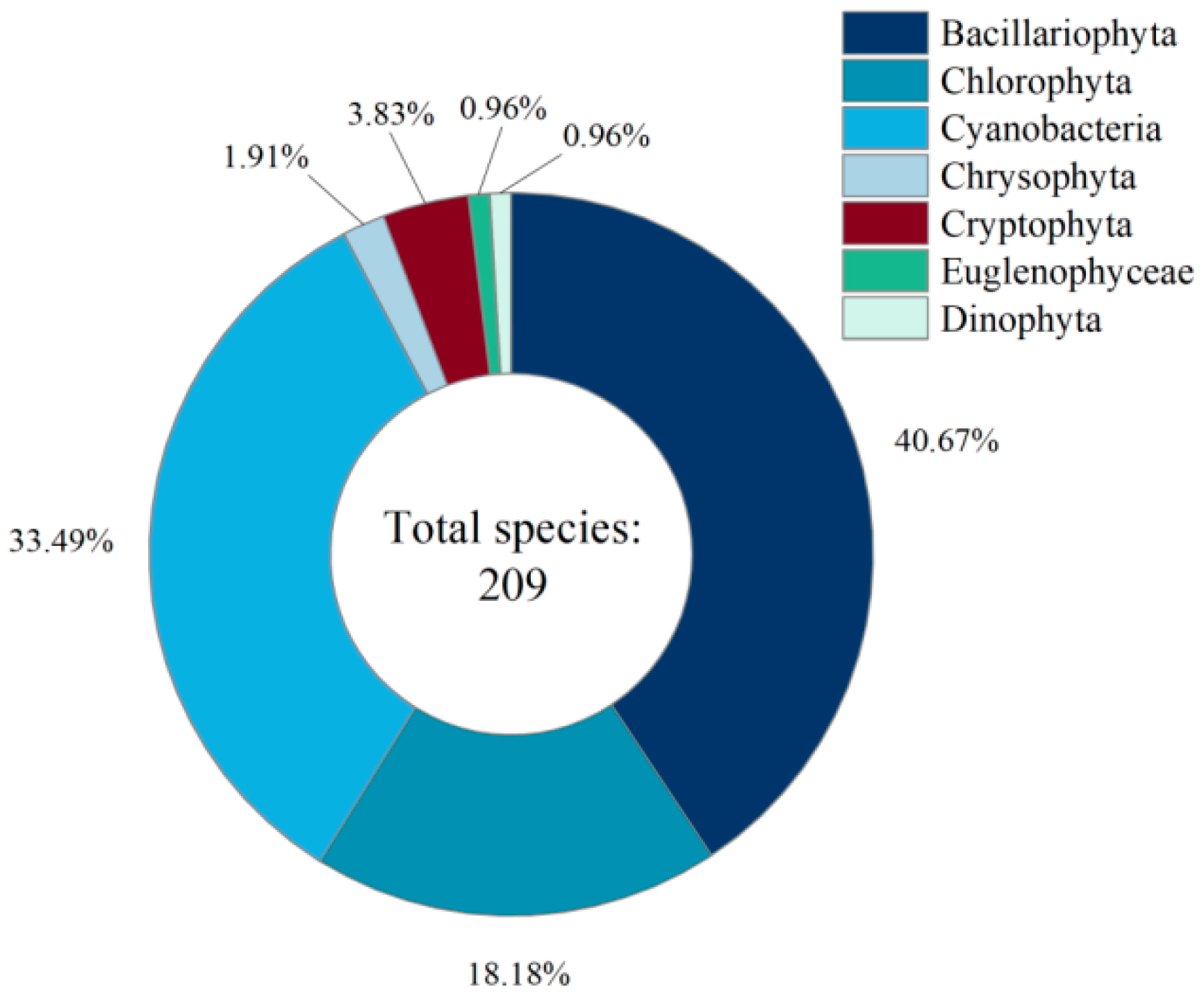
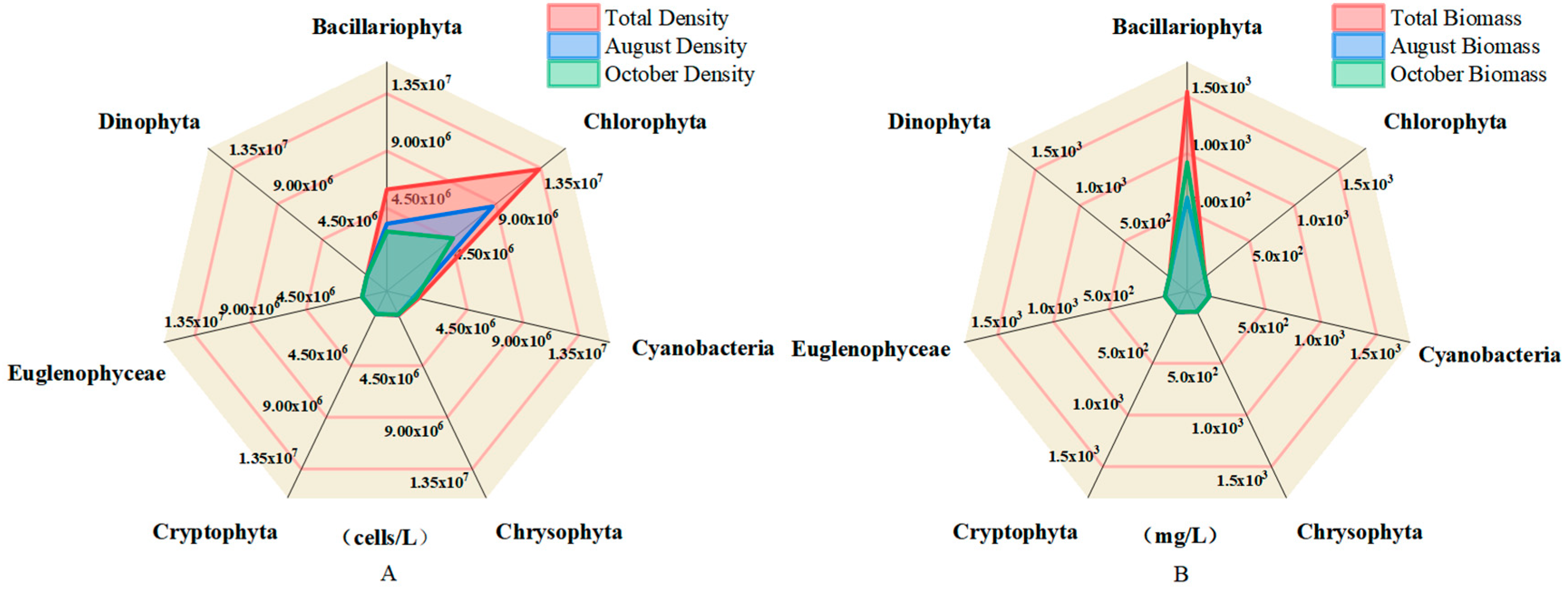
3.1. Taxonomic Composition and Structural Features of Phytoplankton Communities
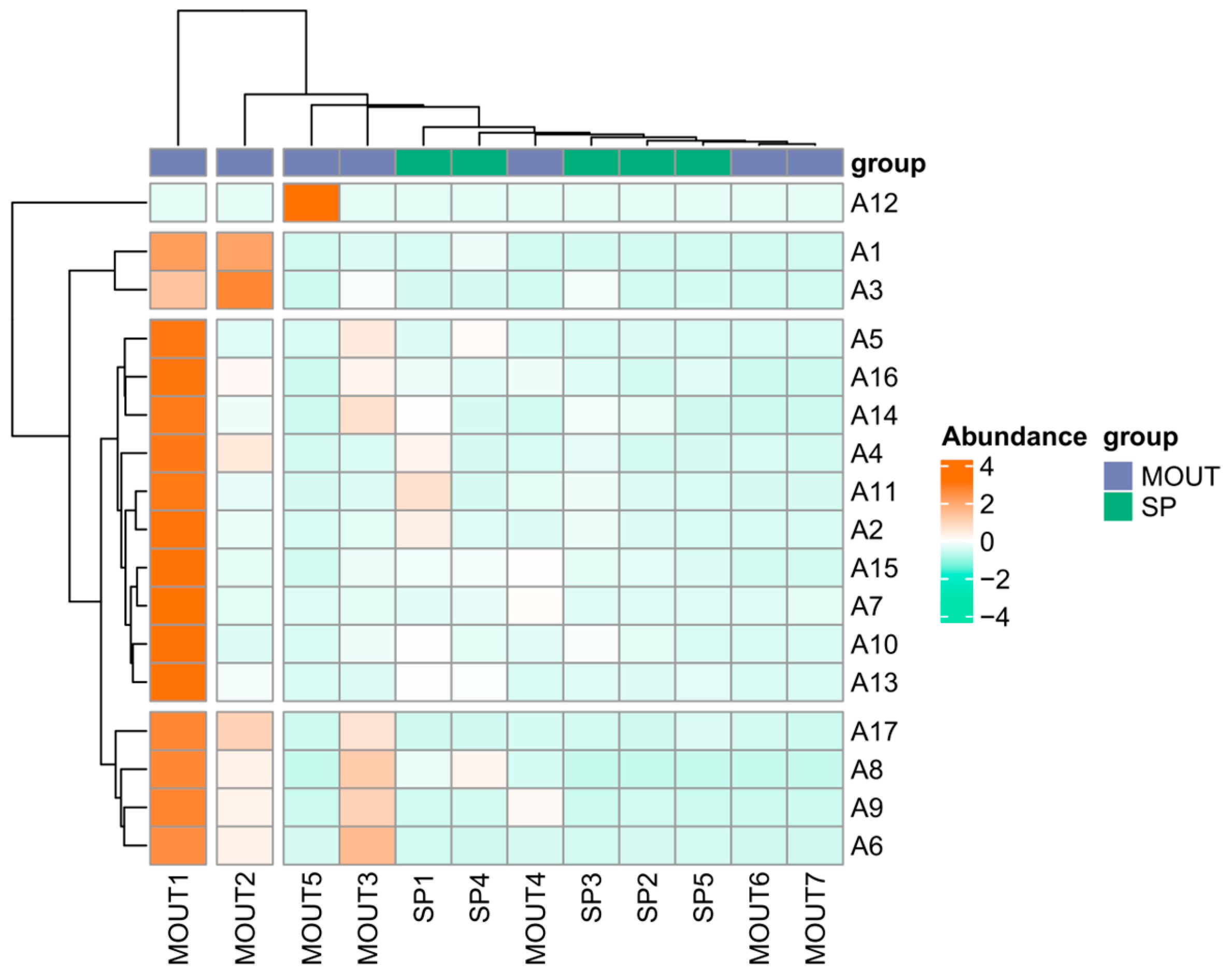
3.2. Characterization of the Environmental Response and Spatial Structure of Phytoplankton Communities
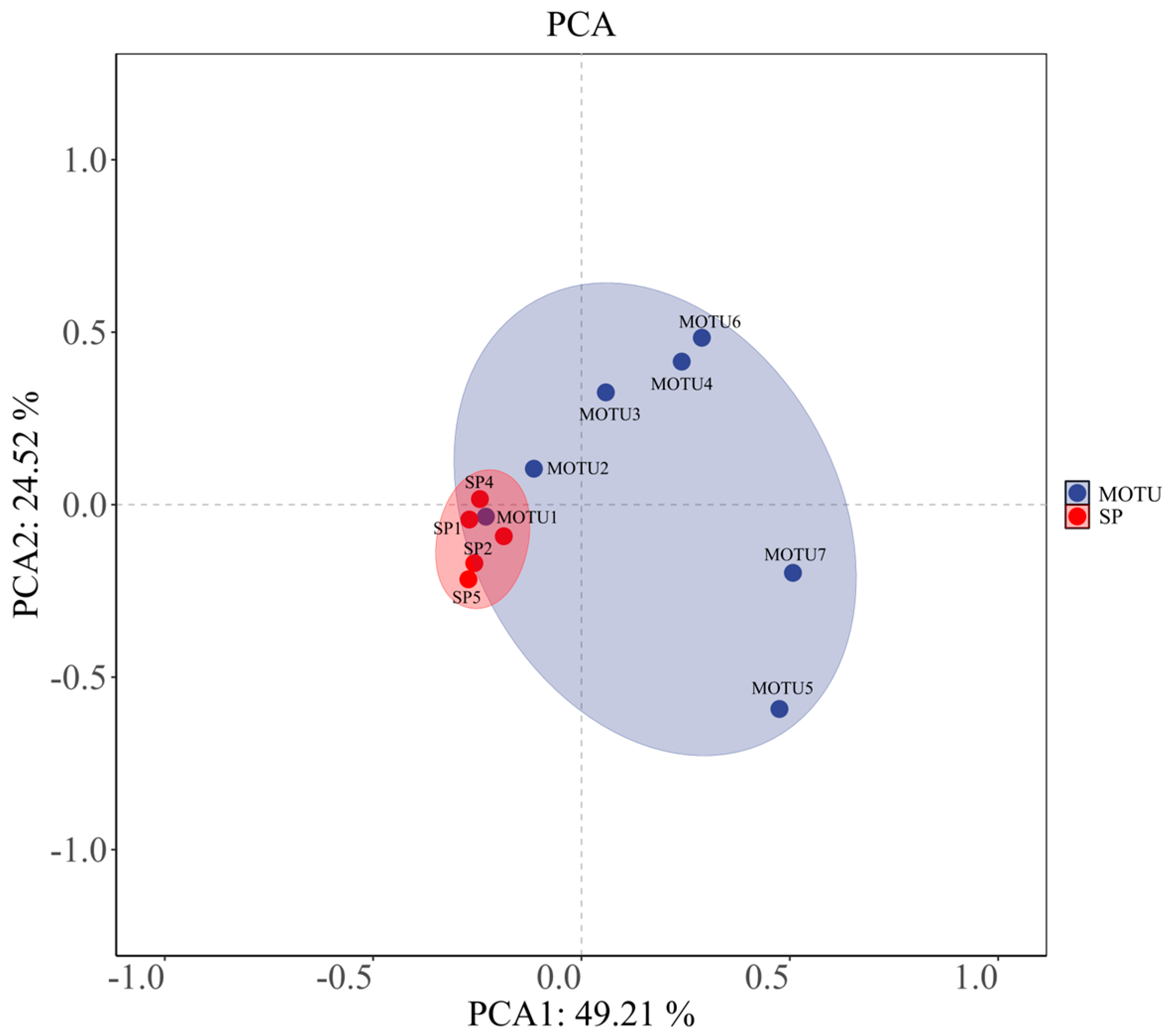
3.3. Multivariate Analysis of Phytoplankton Environmental Response
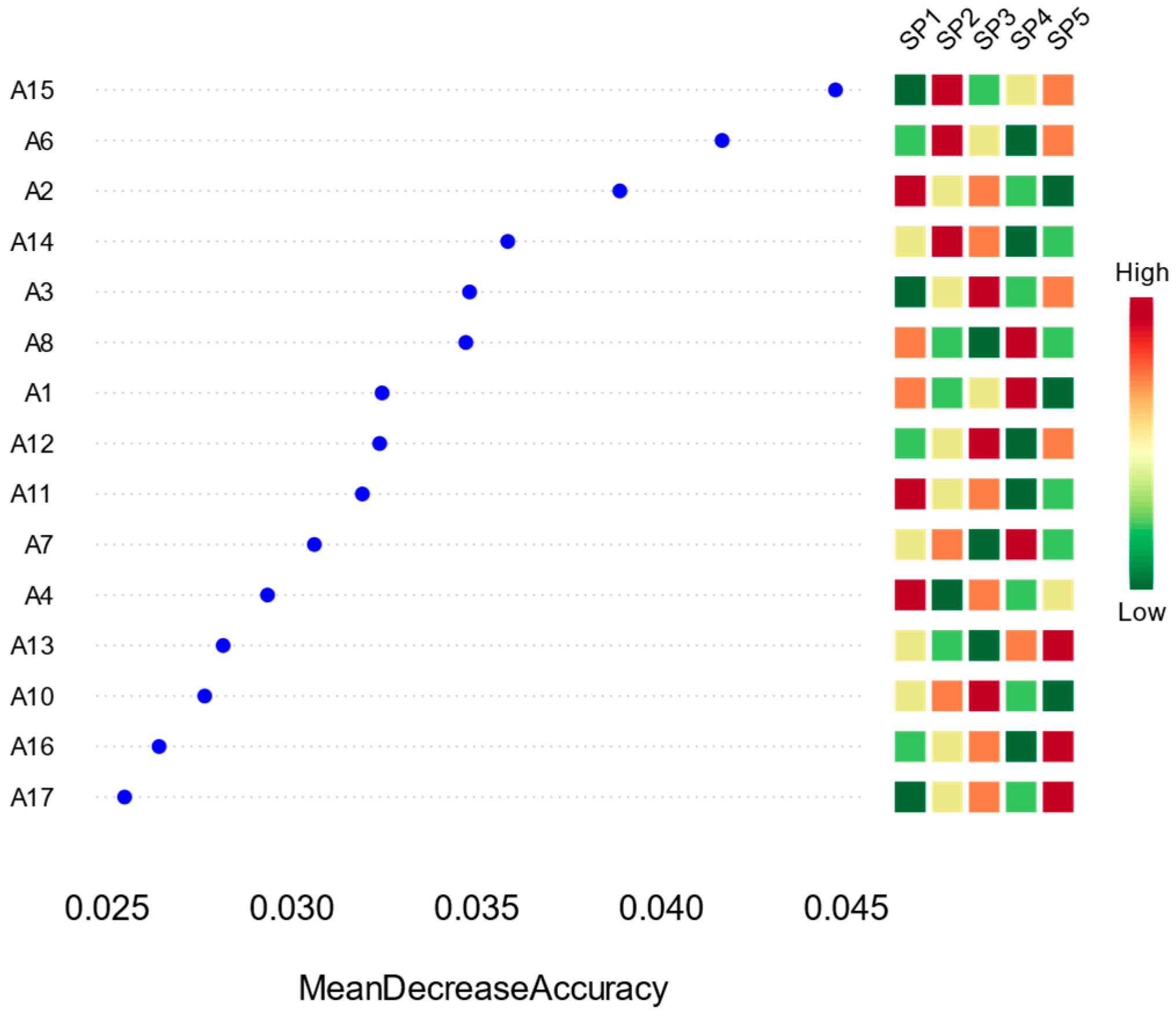
3.4. Identification of Key Phytoplankton Taxa and Ecological Response Analysis Based on a Random Forest Model
4. Discussion
4.1. Characterisation of Phytoplankton Community Structure and Analysis of Dominant Mechanisms
4.2. Community Response Mechanisms and Ecological Niche Differentiation Driven by Environmental Factors
4.3. Phytoplankton Response Patterns at Different Ecological Levels
4.4. Machine-Learning-Based Detection of Core Taxa and Their Environmental Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dai, J.J.; Mei, Y.; Chang, C.C. Stream, Lake, and Reservoir Management. Water Environ. Res. 2017, 89, 1517–1541. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Wan, W.; Zhang, G.; Yao, Z.; Xu, Y.; Liu, B.; Guo, Z.; Luo, Z.; Xiong, W.; Ji, R.; et al. Exploring the topographical pattern beneath the water surface: Global bathymetric volume-area-height curves (BVAH) of inland surface water bodies. Geod. Geodyn. 2024, 15, 602–615. [Google Scholar] [CrossRef]
- Mu, X.J.; Jiang, D.Q.; Hayat, T. Analysis on dynamical behavior of a stochastic phytoplankton-zooplankton model with nonlinear perturbation. Math. Methods Appl. Sci. 2023, 46, 5505–5520. [Google Scholar] [CrossRef]
- Liu, Q.; Tian, Y.L.; Liu, Y.; Yu, M.; Hou, Z.J.; He, K.J.; Xu, H.; Cui, B.S.; Jiang, Y. Relationship between dissolved organic matter and phytoplankton community dynamics in a human-impacted subtropical river. J. Clean. Prod. 2021, 289, 125144. [Google Scholar] [CrossRef]
- Leonard, J.A.; Paerl, H.W. Zooplankton community structure, micro-zooplankton grazing impact, and seston energy content in the St. Johns river system, Florida as influenced by the toxic cyanobacterium Cylindrospermopsis raciborskii. Hydrobiologia 2005, 537, 89–97. [Google Scholar] [CrossRef]
- Hou, L.G.; Deng, Y.J.; Wang, X.L.; Liu, T.; Xu, Y.H.; Wang, J. An Analysis of Policy Transmission Flow in the Chengdu Plain Urban Agglomeration in Southwest China: Towards Building an Ecological Protection Network. Sustainability 2024, 16, 5398. [Google Scholar] [CrossRef]
- Wang, T.W.; Huang, Y.C.; Cheng, J.H.; Xiong, H.; Ying, Y.; Feng, Y.; Wang, J.M. Construction and optimization of watershed-scale ecological network based on complex network method: A case study of Erhai Lake Basin in China. Ecol. Indic. 2024, 160, 111794. [Google Scholar] [CrossRef]
- Kraemer, B.M.; Boudet, S.; Burlakova, L.E.; Haltiner, L.; Ibelings, B.W.; Karatayev, A.Y.; Karatayev, V.A.; Rossbacher, S.; Stoeckli, R.; Straile, D.; et al. An abundant future for quagga mussels in deep European lakes. Environ. Res. Lett. 2023, 18, 124008. [Google Scholar] [CrossRef]
- Kaiser, P.; Hagen, W.; Schukat, A.; Metfies, K.; Biederbick, J.; Dorschner, S.; Auel, H. Phytoplankton diversity and zooplankton diet across Fram Strait: Spatial patterns with implications for the future Arctic Ocean. Prog. Oceanogr. 2025, 234, 103423. [Google Scholar] [CrossRef]
- Çetin, T. A Phytoplankton Composition Index Response to Eutrophication in Turkish Lakes and Reservoirs. Water Air Soil Pollut. 2023, 234, 343. [Google Scholar] [CrossRef]
- Lazareva, V.I.; Zhdanova, S.M.; Sabitova, R.Z.; Sokolova, E.A. Zooplankton of Volga River Reservoirs: Structure, Abundance and Dynamics. Inland Water Biol. 2024, 17, 148–161. [Google Scholar] [CrossRef]
- Li, Z.L.; Mu, Z.X.; Qiu, X.Y.; Liu, J. Changes in future drought characteristics in the Ili River Basin, China, using the new comprehensive standardized drought index. Ecol. Indic. 2025, 173, 113412. [Google Scholar] [CrossRef]
- Guo, N.; Chen, F.L.; He, C.F.; Wang, T.X.; Long, A.H.; Xu, X.W. Multi-year average water vapor characteristics and potential sources and transport pathways of intense water vapor during extreme precipitation events in the Ili River Valley, China. J. Hydrol.-Reg. Stud. 2025, 58, 102278. [Google Scholar] [CrossRef]
- Liu, S.; Long, A.H.; Yan, D.H.; Luo, G.P.; Wang, H. Predicting Ili River streamflow change and identifying the major drivers with a novel hybrid model. J. Hydrol.-Reg. Stud. 2024, 53, 101807. [Google Scholar] [CrossRef]
- Verlecar, X.; Desai, S. Phytoplankton Identification Manual; CSIR-National Institute of Oceanography (NIO): Goa, India, 2004. [Google Scholar]
- John, D.M.; Whitton, B.A.; Brook, A.J. The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Suthers, I.; Rissik, D.; Richardson, A. Plankton: A Guide to Their Ecology and Monitoring for Water Quality; CSIRO publishing: Clayton, Australia, 2019. [Google Scholar]
- Zhang, C.F.; Wu, Y.X.; Zhang, M.C.; Li, Z.Y.; Tian, X.; Li, G.R.; Huang, J.; Li, C. Harnessing diatoms for sustainable economy: Integrating metabolic mechanism with wastewater treatment, biomass production and applications. Algal Res.-Biomass Biofuels Bioprod. 2025, 88, 104031. [Google Scholar] [CrossRef]
- Ullah, N.; Hussain, F.; Nawaz, A.; Khan, M.S.; Saddiq, G.; Zafar, M.; Majeed, S.; Ramadan, M.F.; Makhkamov, T.; Naraliyeva, N.; et al. Integrated Ecohydrology and Systems Biology: Algae Morphotypes and Their Role in Freshwater Ecosystem Functioning. Ecohydrology 2025, 18, e70027. [Google Scholar] [CrossRef]
- Kislioglu, M.S.; Obek, E.; Konakci, N.; Sasmaz, A. Heavy Metal Accumulation in Dominant Green Algae Living in a Habitat Under the Influence of Cu Mine Discharge Water. Plants 2025, 14, 993. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Hong, P.B.; Schmid, B.; De Laender, F.; Eisenhauer, N.; Zhang, X.W.; Chen, H.Z.; Craven, D.; De Boeck, H.J.; Hautier, Y.; Petchey, O.L.; et al. Biodiversity promotes ecosystem functioning despite environmental change. Ecol. Lett. 2022, 25, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, E.; Smith, E.M.; Richardson, T.L. Spectral Irradiance, Phytoplankton Community Composition and Primary Productivity in a Salt Marsh Estuary, North Inlet, South Carolina, USA. Estuaries Coasts 2013, 36, 347–364. [Google Scholar] [CrossRef]
- Van Vliet, M.T.H.; Thorslund, J.; Strokal, M.; Hofstra, N.; Flörke, M.; Macedo, H.E.; Nkwasa, A.; Tang, T.; Kaushal, S.S.; Kumar, R.; et al. Global river water quality under climate change and hydroclimatic extremes. Nat. Rev. Earth Environ. 2023, 4, 687–702. [Google Scholar] [CrossRef]
- Huo, D.; Gan, N.Q.; Geng, R.Z.; Cao, Q.; Song, L.R.; Yu, G.L.; Li, R.H. Cyanobacterial blooms in China: Diversity, distribution, and cyanotoxins. Harmful Algae 2021, 109, 102106. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bai, M.G.; Yao, L.L.; Ma, J.; He, F.; Bian, G.D.; Li, W.X. Phytoplankton and Zooplankton Community Dynamics in an Alpine Reservoir: Environmental Drivers and Ecological Implications in Daqing Reservoir, China. Water 2025, 17, 1202. [Google Scholar] [CrossRef]
- Amorim, C.A.; Moura, A.D. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhu, Y.; Hu, X.Y.; Zhu, B.R.; Lyu, S.; Yinglan, A.; Wang, G.Q. Assembly mechanism and stability of zooplankton communities affected by China’s south-to-north water diversion project. J. Environ. Manag. 2024, 365, 121497. [Google Scholar] [CrossRef] [PubMed]
- Vezhnavets, V.V.; Kouraev, A.V.; Gukasyan, E.K.; Gabrielyan, B.K. Zooplankton Study of Lake Sevan as an Indicator of Ecosystem Stability in the Context of Global Climate Change. Inland Water Biol. 2024, 17, 48–58. [Google Scholar] [CrossRef]
- Wu, M.; Yan, H.; Fu, S.; Han, X.; Jia, M.; Dou, M.; Liu, H.; Fohrer, N.; Messyasz, B.; Li, Y. Seasonal Dynamics of Planktonic Algae in the Danjiangkou Reservoir: Nutrient Fluctuations and Ecological Implications. Sustainability 2025, 17, 406. [Google Scholar] [CrossRef]
- Wang, C.; Jia, H.; Wei, J.; Yang, W.; Gao, Y.; Liu, Q.; Ge, D.; Wu, N. Phytoplankton functional groups as ecological indicators in a subtropical estuarine river delta system. Ecol. Indic. 2021, 126, 107651. [Google Scholar] [CrossRef]
- Krylov, A.V.; Hayrapetyan, A.O.; Kosolapov, D.B.; Sakharova, E.G.; Kosolapova, N.G.; Sabitova, R.Z.; Malin, M.I.; Malina, I.P.; Gerasimov, Y.V.; Hovsepyan, A.A.; et al. Features of Structural Changes in the Plankton Community of An Alpine Lake with Increasing Fish Density in Summer and Autumn. Zool. Zhurnal 2021, 100, 147–158. [Google Scholar] [CrossRef]
- Manier, J.T.; Haro, R.J.; Houser, J.N.; Strauss, E.A. Spatial and temporal dynamics of phytoplankton assemblages in the upper Mississippi River. River Res. Appl. 2021, 37, 1451–1462. [Google Scholar] [CrossRef]
- Yang, Y.; Colom, W.; Pierson, D.; Pettersson, K. Water column stability and summer phytoplankton dynamics in a temperate lake (Lake Erken, Sweden). Inland Waters 2016, 6, 499–508. [Google Scholar] [CrossRef]
- Burdis, R.M.; Ward, N.K.; Manier, J.T. Phytoplankton assemblage dynamics in relation to environmental conditions in a riverine lake. Aquat. Ecol. 2025, 59, 467–485. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Wu, Y.F.; Xu, Y.J.; Yu, Y.X.; Zhang, G.X.; Mao, D.H.; Liu, X.M.; Dai, C.L. Phytoplankton growth and succession driven by topography and hydrodynamics in seasonal ice-covered lakes. Ecol. Inform. 2025, 86, 103053. [Google Scholar] [CrossRef]
- Rao, K.; Cao, X.; Wang, Y.F.; Zhang, Y.Q.; Huang, H.S.; Ma, Y.L.; Xu, J. Spatial-temporal distributions of phytoplankton shifting, chlorophyll-a, and their influencing factors in shallow lakes using remote sensing. Ecol. Inform. 2024, 82, 102765. [Google Scholar] [CrossRef]
- Chen, Y.C.; Yue, Y.H.; Wang, J.; Li, H.R.; Wang, Z.K.; Zheng, Z. Microbial community dynamics and assembly mechanisms across different stages of cyanobacterial bloom in a large freshwater lake. Sci. Total Environ. 2024, 907, 168207. [Google Scholar] [CrossRef] [PubMed]
- Gadelha, E.S.; Dunck, B.; Colares, L.F.; Akama, A. Temporal patterns of alpha and beta diversities of microzooplankton in a eutrophic tidal river in the eastern Amazon. Limnology 2023, 24, 193–204. [Google Scholar] [CrossRef]
- Picapedra, P.H.S.; Fernandes, C.; Taborda, J.; Baurrigartneri, G.; Sanches, P. A long-term study on zooplankton in two contrasting cascade reservoirs (Iguacu River, Brazil): Effects of inter-annual, seasonal, and environmental factors. PeerJ 2020, 8, e8979. [Google Scholar] [CrossRef] [PubMed]
- Lamouille-Hébert, M.; Arthaud, F.; Datry, T. Climate change and the biodiversity of alpine ponds: Challenges and perspectives. Ecol. Evol. 2024, 14, e10883. [Google Scholar] [CrossRef] [PubMed]
- Houliez, E.; Lefebvre, S.; Dessier, A.; Huret, M.; Marquis, E.; Bréret, M.; Dupuy, C. Spatio-temporal drivers of microphytoplankton community in the Bay of Biscay: Do species ecological niches matter? Prog. Oceanogr. 2021, 194, 102558. [Google Scholar] [CrossRef]
- Zhong, Y.P.; Liu, X.; Xiao, W.P.; Laws, E.A.; Chen, J.X.; Wang, L.; Liu, S.G.; Zhang, F.; Huang, B.Q. Phytoplankton community patterns in the Taiwan Strait match the characteristics of their realized niches. Prog. Oceanogr. 2020, 186, 102366. [Google Scholar] [CrossRef]
- Piccioni, F.; Casenave, C.; Baragatti, M.; Cloez, B.; Vinçon-Leite, B. Calibration of a complex hydro-ecological model through Approximate Bayesian Computation and Random Forest combined with sensitivity analysis. Ecol. Inform. 2022, 71, 101764. [Google Scholar] [CrossRef]
- Hipsey, M.R.; Gal, G.; Arhonditsis, G.B.; Carey, C.C.; Elliott, J.A.; Frassl, M.A.; Janse, J.H.; de Mora, L.; Robson, B.J. A system of metrics for the assessment and improvement of aquatic ecosystem models. Environ. Model. Softw. 2020, 128, 104697. [Google Scholar] [CrossRef]
- Zhou, Z.X.; Yu, R.C.; Zhou, M.J. Seasonal succession of microalgal blooms from diatoms to dinoflagellates in the East China Sea: A numerical simulation study. Ecol. Model. 2017, 360, 150–162. [Google Scholar] [CrossRef]
- Dunker, S.; Nadrowski, K.; Jakob, T.; Kasprzak, P.; Becker, A.; Langner, U.; Kunath, C.; Harpole, S.; Wilhelm, C. Assessing in situ dominance pattern of phytoplankton classes by dominance analysis as a proxy for realized niches. Harmful Algae 2016, 58, 74–84. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phyla | Species | Dominance | Codes |
|---|---|---|---|
| Bacillariophyta | Nitzschia palea (Kützing) W. Smith | 0.07 | SP1 |
| Bacillariophyta | Achnanthes exigua Grunow | 0.02 | SP2 |
| Bacillariophyta | Synedra acus Kützing | 0.02 | SP3 |
| Bacillariophyta | Cymbella cistula (Ehrenberg) Kirchner | 0.02 | SP4 |
| Chlorophyta | Limnothrix redekei (Van Goor) Meffert | 0.24 | SP5 |
| Bacillariophyta | - | - | MOTU1 |
| Chlorophyta | - | - | MOTU2 |
| Cyanobacteria | - | - | MOTU3 |
| Chrysophyta | - | - | MOTU4 |
| Cryptophyta | - | - | MOTU5 |
| Euglenophyceae | - | - | MOTU6 |
| Dinophyta | - | - | MOTU7 |
| Parameter | Unit | August (Mean ± SD) | Range (Aug) | October (Mean ± SD) | Range (Oct) | ANOVA F-Value | p-Value |
|---|---|---|---|---|---|---|---|
| WT | °C | 21.82 ± 2.40 | 18.11–26.20 | 14.62 ± 1.90 | 11.20–17.80 | 115.72 | <0.001 |
| pH | – | 7.93 ± 0.25 | 7.52–8.45 | 7.61 ± 0.18 | 7.29–7.95 | 14.83 | <0.01 |
| EC | μS/cm | 650 ± 81 | 505–812 | 683 ± 75 | 545–810 | 2.57 | 0.118 |
| Sal | ppt | 0.34 ± 0.04 | 0.27–0.42 | 0.36 ± 0.03 | 0.30–0.42 | 2.02 | 0.163 |
| TDS | mg/L | 437 ± 61 | 331–538 | 452 ± 59 | 355–547 | 1.78 | 0.191 |
| ORP | mV | 124 ± 27 | 85–179 | 141 ± 31 | 96–188 | 5.94 | 0.024 |
| DO | mg/L | 6.42 ± 1.14 | 4.41–8.26 | 7.86 ± 0.92 | 6.35–9.33 | 28.73 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zi, F.; Song, T.; Cai, W.; Liu, J.; Ma, Y.; Lin, X.; Zhao, X.; Hu, B.; Ren, D.; Song, Y.; et al. Environmental Drivers of Phytoplankton Structure in a Semi-Arid Reservoir. Biology 2025, 14, 914. https://doi.org/10.3390/biology14080914
Zi F, Song T, Cai W, Liu J, Ma Y, Lin X, Zhao X, Hu B, Ren D, Song Y, et al. Environmental Drivers of Phytoplankton Structure in a Semi-Arid Reservoir. Biology. 2025; 14(8):914. https://doi.org/10.3390/biology14080914
Chicago/Turabian StyleZi, Fangze, Tianjian Song, Wenxia Cai, Jiaxuan Liu, Yanwu Ma, Xuyuan Lin, Xinhong Zhao, Bolin Hu, Daoquan Ren, Yong Song, and et al. 2025. "Environmental Drivers of Phytoplankton Structure in a Semi-Arid Reservoir" Biology 14, no. 8: 914. https://doi.org/10.3390/biology14080914
APA StyleZi, F., Song, T., Cai, W., Liu, J., Ma, Y., Lin, X., Zhao, X., Hu, B., Ren, D., Song, Y., & Chen, S. (2025). Environmental Drivers of Phytoplankton Structure in a Semi-Arid Reservoir. Biology, 14(8), 914. https://doi.org/10.3390/biology14080914