1. Introduction
Plant diversity, a key component of biodiversity, underpins ecosystem stability and human well-being [
1,
2,
3]. However, global climate change, accelerating urbanization, and rapid land-use changes have led to extensive habitat loss and increased landscape fragmentation, posing serious threats to plant diversity [
4,
5]. In recent years, understanding the spatial patterns of plant species diversity and their driving mechanisms have become a central focus in biodiversity research [
6]. Such knowledge is essential for elucidating community assembly and succession processes and provides a theoretical foundation for assessing ecosystem responses to environmental disturbances and their potential for recovery [
7].
In recent decades, the impacts of global climate change have intensified, with arid and semi-arid ecosystems being particularly sensitive due to their chronic water limitations and limited buffering capacity [
8]. Climate change acts as a primary driver influencing primary productivity, water availability, and community structure, and can reconfigure ecohydrological regimes and vegetation trajectories [
9,
10]. Broad-scale climatic variables, such as temperature and precipitation, shape species distribution and ecological strategies by altering habitat suitability and resource accessibility [
11,
12]. The topographic heterogeneity contributes to microclimatic diversity, with factors such as elevation and slope regulating solar radiation, temperature gradients, and soil moisture. These conditions create a variety of ecological niches, promoting species coexistence and increasing community structural complexity [
13,
14]. In parallel, human activities restructure landscape composition and configuration, intensifying habitat fragmentation, severing ecological corridors, and increasing dispersal resistance, thereby constraining species’ ranges and ultimately eroding regional plant biodiversity [
15].
Landscape patterns, as spatial representations of ecosystem structure and function, are increasingly recognized in biodiversity research [
16]. The metrics describing the patch number, shape, area, and spatial configuration capture the degree of heterogeneity and stability within ecosystems, directly influencing habitat accessibility, dispersal routes, and population persistence [
17,
18]. Under global environmental change, shifts in landscape structures have become the key drivers of plant community composition and ecosystem functioning [
16,
19]. The landscape connectivity, which reflects the spatial continuity between habitat patches, plays a critical role in maintaining ecological processes and stabilizing species dynamics [
20,
21]. It influences species distribution at the landscape scale and is shaped by matrix permeability, patch configuration, and species-specific traits [
22,
23]. Although growing research has linked connectivity to dispersal, migration, and genetic diversity, most studies still examine climate impacts on species distributions in isolation, lacking an integrated view with the landscape configuration and connectivity [
9,
24]. Empirical studies that integrate climate, landscape structure, and connectivity in a unified analytical framework remain limited [
25,
26]. Moreover, conventional approaches, such as generalized linear mixed models and redundancy analyses, struggle to capture the complex, nonlinear, and mediated pathways among the interacting variables—limitations especially evident in structurally complex arid ecosystems [
1,
5,
27].
Partial least squares structural equation modeling (PLS-SEM) has emerged as a powerful tool for disentangling multifactor causal mechanisms [
28,
29]. It effectively handles non-normal data, multicollinearity, and small samples, enabling the simultaneous estimation of direct, indirect, and mediating effects among the latent constructs [
30]. PLS-SEM has been successfully applied to model ecological interactions in highly variable environments, particularly in drylands where the theoretical frameworks are often uncertain. Previous studies have used this method to reveal how climate, land use, and connectivity jointly affect biodiversity across forests, bird communities, and arid vegetation systems, highlighting its theoretical adaptability and empirical strength [
31,
32].
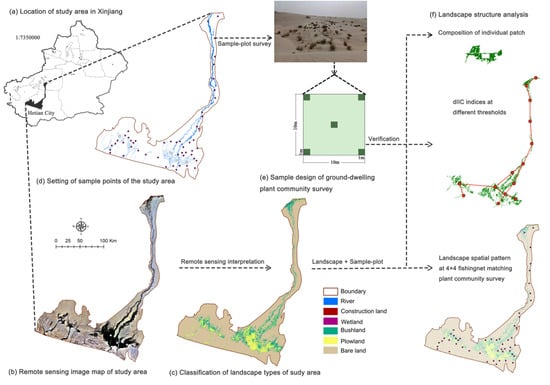
This study focuses on the Hotan River Basin, a typical arid inland river catchment in the southwestern Tarim Basin, China. The region features a distinct desert–oasis transition zone with important ecological functions and biodiversity value [
33]. Over the past two decades, hydrological interventions, such as the Tarim River regulation project and “water transfer from north to south”, have significantly altered the local hydrology and landscape structure, with pronounced effects on the patch configuration, connectivity, and biodiversity. Against this backdrop, based on the theory of environmental sustainability, from the perspective of scale–place–space, our goal was to evaluate how environmental factors influence plant diversity directly and indirectly via the landscape structure and connectivity, and to place these pathways in a multi-scale, place-based sustainability context. Using multi-temporal remote sensing data from 2000, 2012, and 2023, together with field vegetation plots, DEM-derived terrain metrics, and climate data, we constructed a PLS-SEM model to explore the interactions among the environmental factors, landscape patterns, connectivity, and plant diversity. The specific objectives were as follows: (1) characterize the landscape pattern changes across the three time periods and, using the Patch-generating Land Use Simulation (PLUS) model, project the land cover for 2035 and 2050 under SSP1-2.6, SSP2-4.5, and SSP5-8.5; (2) analyze spatial patterns of plant diversity along hydro-topographic gradients; and (3) identify the causal pathways linking the environmental and spatial factors to plant diversity. By embedding dryland biodiversity within an environment → structure → function → diversity causal chain, the study delivers pathway-based, connectivity-informed evidence to guide climate-robust corridor design, water–land coordination, and conservation prioritization in arid river basins, thereby advancing a scalable (scale–place–space) framework for environmental sustainability.
4. Discussion
4.1. Landscape Area Dynamics and Conversion Pathways
At the basin scale, land-use/cover change in arid river systems reflects coupled hydroclimatic and anthropogenic influences on landscape composition [
8]. In the Hotan River Basin (2000–2023), bare land remained the dominant class (~83% of area) but exhibited a consistent net decline, with most conversions directed to shrubland and, secondarily, cultivated land. This shift mirrors the patterns reported for desert–oasis ecotones across the Tarim Basin, where modest improvements in water availability and local water management have facilitated vegetative infilling on formerly barren substrates [
49]. Construction land, although a small fraction of the total, expanded rapidly—largely at the expense of shrubland—consistent with peri-urban growth and settlement infill along oasis margins and transport corridors [
50]. Together, these trajectories indicate a dual-driver regime: the hydrologically enabled “greening” of bare surfaces alongside the steady encroachment of built-up areas into ecologically functional spaces.
The forward simulations reinforce these tendencies while clarifying pathway differences. Under SSP1-2.6, bare land decreases and both shrubland and cultivated land increase by 2035 and 2050, whereas construction land expands in all the scenarios, most prominently under SSP5-8.5. SSP2-4.5 yields mixed responses in the vegetated classes but maintains the construction land’s growth. Read against the historical baseline, these projections imply a continued net conversion from bare land to vegetated types, with the rate and spatial footprint contingent on the socioeconomic trajectory and water availability. In short, the basin’s landscape is likely to remain dominated by bare land and shrubland through mid-century, with the persistent expansion of construction land—a combination that will keep the composition of the desert–oasis mosaic tightly governed by both climate-sensitive hydrology and human development.
4.2. Vegetation Distribution and Hydrothermal Adaptation
Our study reveals that the vegetation distribution in the study area shows strong alignment with the environmental gradients, especially precipitation and slope. We reveal that the areas with moderate slopes and annual precipitation between 25 and 36 mm exhibit the highest diversity indices (H′ and D) and relatively stable community structures. This finding supports the ecological niche theory in arid zones, where species richness peaks under intermediate energy and stress conditions [
26,
27,
51]. The dominant species, such as
Populus euphratica and
Tamarix chinensis, demonstrate high adaptability to steep slopes and low water availability, forming critical transitional communities between riparian zones and deserts [
11,
32].
The composition and diversity of plant communities are influenced by the combined effects of slope and precipitation. The lower and mid-slope zones (L and M) support higher species richness, including regional endemics such as
Calligonum roborowskii and
Seriphidium korovinii, which are sensitive indicators of desertification. In contrast, the high-slope zones (H) exhibit limited species richness due to the constraints imposed by both moisture availability and the topographic conditions. These patterns align with broader desert ecosystem studies from Central Asia, affirming the role of slope–precipitation interactions in shaping biodiversity [
14,
27,
33].
4.3. Pathway Mechanisms Linking Landscape Pattern, Connectivity, and Biodiversity
4.3.1. Direct and Indirect Effects of Landscape Pattern on Biodiversity
The spatiotemporal configuration of landscapes is a major component and driver of global change, with pervasive consequences for ecosystem services and biodiversity. In our PLS-SEM, the landscape pattern (LSP) exerts both a direct negative effect on plant diversity (β_direct = −0.189,
p < 0.05) and an additional negative indirect effect via connectivity (β_indirect = −0.103,
p < 0.05). These pathways are consistent with the 2000–2023 observations of stronger fragmentation and weaker aggregation in the basin (NP/PD and SHDI/SHEI ↑; AI/CONTAG ↓; IJI and DIVISION ↑; see
Section 3.1.3).
Ecologically, the LSP indicators used here capture distinct mechanisms. The ED and LSI tend to intensify the edge effects under hot, high-resistance matrices, which lowers the J and raises the D, and also suppresses the functional connectivity (IIC/PC). The PD and DIVISION quantify fragmentation and isolation; increases in them typically reduce dispersal and rescue effects, thereby depressing the H′ and J [
33,
52]. The LPI reflects the control of the dominant matrix. In the Hotan River Basin—where bare land retains the largest patch—the high LPI implies a resistance-dominated matrix that disrupts path continuity and accessibility, lowering the IIC/PC and diversity metrics; the converse can occur if the largest patch is a continuous shrubland corridor [
20,
41]. Finally, SHDI/SHEI indicate compositional heterogeneity and balance. When fragmentation remains below the threshold, a higher SHDI/SHEI can increase microhabitat variety and support a higher H′; however, if rising SHDI/SHEI co-occurs with CONTAG decline and PD/DIVISION increase, heterogeneity shifts toward a finely “speckled” mosaic that undermines connectivity and community stability [
53].
Together, these processes explain the net negative direct effect of LSP on diversity in an arid, high-resistance context: elevated ED/LSI/PD/DIVISION and a persistently high bare land LPI outweigh any benefits of moderate heterogeneity. Because the LSP also depresses connectivity, and connectivity in turn promotes diversity (
Section 4.3.2), the LSP → CON → DIV route contributes an additional negative (competitive) mediation. These findings align with the circuit-theory arguments that functional connectivity, not mere structural adjacency, governs ecological exchange in fragmented drylands, and with the threshold responses to fragmentation reported for arid systems [
52].
4.3.2. Connectivity as the Bridge from Structure to Biodiversity
The connectivity (CON) showed a significant positive direct effect on diversity (β = 0.242,
p < 0.001), whereas ENV → CON was non-significant, making LSP → CON → DIV the principal indirect pathway. The IIC and PC increased with an assumed dispersal distance and plateaued near ~10 km, indicating a functional threshold: below this scale, the network remains partly broken; above it, most vegetated patches become functionally linked. This threshold accords with the field conditions—wind and seasonal flows are the dominant vectors—and explains why local expansions of built-up land or extensive bare land matrices (“hard edges–high resistance–breaks”) markedly reduce the effective path probabilities and increase travel times [
40]. Empirical and modeling studies similarly show that once the IIC/PC cross a scale-dependent threshold, colonization/gene flow accelerate and communities stabilize; below it, systems drift toward isolation and degradation [
20]. Thus, raising the IIC/PC is not a mere numeric gain; they represent lower dispersal costs, more permeable corridors, and higher renewal probabilities—the proximate mechanisms behind the positive CON → DIV effect.
The observed coupling between the LSP and CON is also clear: a higher PD/DIVISION with a lower AI/CONTAG suppresses the IIC/PC, whereas linear or stepping-stone ribbons of shrubland/wetland along riparian corridors (moderate ED/LSI, higher CONTAG) enhance functional linkages. Given that ENV → CON was not significant, the basin’s functional network is generated through a staged process—first reconfiguring the structure (LSP), then expressing connectivity (CON). Consequently, the rising SHDI/SHEI between 2000 and 2023 does not automatically translate into diversity gains when the bare land LPI remains high and the PD/DIVISION increase, unless the ~10 km functional threshold is met. Connectivity, therefore, is the necessary bridge that converts “pattern optimization” into sustained biodiversity benefits.
4.3.3. Hierarchical Roles of Environmental Factors
The environmental factors (ENV) had significant positive direct effects on both the diversity and landscape pattern (ENV → DIV: β = 0.341; ENV → LSP: β = 0.541; both
p < 0.001), but no direct effect on the connectivity (ENV → CON: β = −0.207, 95% CI includes 0). This implies a staged “environment → structure → function → diversity” process: the environmental constraints first reshape the patch geometry and composition (LSP), which then alter the dispersal costs and accessibility (CON), ultimately influencing community diversity (DIV) [
20,
54,
55].
The ENV latent variable comprised the D_set (negative loading indicating human disturbance), gwd, prcp, slope, and st. The disturbance gradient matches the spatial signal of a rising PD/LSI/IJI and a declining CONTAG (
Section 3.1.3). The gwd directly constrains
Populus/
Tamarix persistence; deeper water tables contract shrubland patches and depress SHDI/SHEI, narrowing riparian corridors and lowering the IIC/PC and the H′/J [
51,
56]. Water availability is the primary limitation in drylands [
49]; in our plots, the H′ and D peaked at ~32–36 mm annual precipitation (
Section 3.2), consistent with enhanced coexistence and rescue effects at moderate moisture, and with concomitant gains in SHDI/SHEI. The slope captures microtopographic control of soil moisture, radiation, and substrate stability; steeper slopes lengthen edges (ED) and increase shape complexity (LSI), raise the PD/ED and reduce the IIC/PC once the thresholds are exceeded, and lower the establishment probabilities (H′/J down, D up) [
57]. The observed advantage of low–mid slopes for cover and diversity (
Section 3.2) corroborates these mechanisms. The soil type governs water holding and nutrients, often sustaining a high bare land LPI, lower CONTAG, and higher DIVISION, which depress the IIC/PC and shifts communities toward a lower H′/J and higher D [
54].
In sum, the positive ENV → DIV pathway reflects the direct benefits of moisture and favorable topo-edaphic conditions; the strong ENV → LSP effect indicates that the environment first reconfigures the spatial “skeleton”; the lack of a direct ENV → CON link suggests that connectivity improvements depend on prior structural modification in this basin.
4.3.4. An Integrated Chain Mechanism and Its Consistency with Prior Work
Collectively, the model supports a four-level chain: ENV → LSP → CON → DIV. The LSP fully mediates ENV → CON, and DIV is co-determined by the direct environmental signal (β = 0.341), structural constraint from the LSP (β = −0.189), and functional promotion by CON (β = 0.253), plus multiple indirect paths. The chained mediation (ENV → LSP → CON → DIV) has the largest total effect (0.389), indicating that in arid mosaics, environmental constraints typically act first through spatial reconfiguration and then through functional linkage to shape biodiversity.
These findings are consistent with large-scale syntheses in which climate/water set broad species distributions, whereas landscape-level heterogeneity and connectivity exert stronger control on local diversity [
49,
58,
59]. They also echo cross-regional comparisons (e.g., Frishkoff [
60]; Beissinger [
61]) while underscoring a dryland specificity: under high-resistance matrices—reflected by the high bare land LPI—and concurrent human disturbance, functional connectivity becomes the key buffer against climatic stress. Critically, only when networks surpass the ~10 km dispersal threshold and structural resistance is reduced can increases in SHDI/SHEI yield net biodiversity gains rather than fragmentation costs.
4.4. Theoretical and Methodological Contributions
Anchored in the environmental sustainability framework of Ribeiro et al. [
62], this study adopts a scale–place–space perspective and embeds dryland biodiversity into a causal chain of environment → structure → function → diversity, yielding a transferable analytical framework for arid social–ecological systems. At the scale dimension, macro-level hydroclimate and human disturbance are downscaled into meso-level landscape metrics (PD, ED, LSI, LPI, SHDI, SHEI, DIVISION) and micro-level functional connectivity (PC, IIC), revealing an ~10 km dispersal threshold that links landscape resistance to realized community diversity. At the place dimension, the groundwater depth, soil type, and proximity to settlements are explicitly incorporated for the oasis–desert context, showing that compositional heterogeneity translates into biodiversity gains only when fragmentation remains sub-threshold and matrix resistance is reduced. At the space dimension, remotely sensed configuration change is coupled with movement-related functional processes and the SSP1-2.6/SSP2-4.5/SSP5-8.5 scenarios, converting “pattern change” into an operational process diagnosis. Overall, the work clarifies how scale interacts with geographic context, extends sustainability theory to arid inland river basins, and provides a reusable indicator set and framework to inform biodiversity conservation and landscape governance.
Methodologically, this study advances landscape–ecological modeling in two complementary ways. First, it integrates environmental (ENV), structural (LSP), and functional (CON) components within a PLS-SEM architecture, thereby identifying the multiple indirect and chain-mediated pathways among the drivers and biodiversity responses. Multi-source data—multi-temporal satellite imagery, DEM-derived terrain, climate records, and 57 field plots—are harmonized on a 4 × 4 km2 grid to ensure cross-dataset comparability. Pairing classic landscape indices (e.g., SHDI, PD, CONTAG) with Conefor-based connectivity metrics (PC, IIC) enables a comprehensive assessment of structure–function–diversity linkages and moves beyond a metric description toward mechanism-oriented inference. Second, we couple PLUS with a causal framework to bridge “what-is” and “what-if.” The PLUS model combines machine learning suitability (LEAS/Random Forest) with patch-level CA dynamics (CARS), allowing us to (i) explain the land-use drivers and simulate patch evolution consistent with recent applications in China’s drylands, and (ii) generate scenario outputs (SSP1-2.6/SSP2-4.5/SSP5-8.5; 2035/2050) that are directly interpretable within the ENV → LSP → CON → DIV chain. In practice, PLS-SEM quantifies how environmental constraints propagate through spatial configurations and functional connectivity to affect diversity, while PLUS projects the spatial trajectories of those constraints under alternative futures. Together, they deliver a transferable, scale-aware template that links diagnosis (causal pathways) with prognosis (scenario-based maps), thereby positioning our findings squarely within contemporary environmental—sustainability research and supplying actionable evidence for dryland river-basin planning.
4.5. Implications and Limitations
4.5.1. Implications for Sustainable Landscape Planning
Our findings translate the “environment → structure → function → diversity” chain into four actionable principles for arid river basins. (i) Connectivity-first design: Because biodiversity increased with the functional connectivity and an ~10 km dispersal threshold emerged from the IIC/PC analysis, planning should prioritize continuous riparian and shrubland corridors and maintain stepping-stone patches at ≤10 km intervals, while minimizing the breaks created by extensive bare land or new construction. (ii) Manage fragmentation, not just heterogeneity: The instances where SHDI/SHEI rose alongside a higher PD/DIVISION and lower AI/CONTAG show that more heterogeneous mosaics can still undermine habitat function; management targets should therefore include reducing the bare land LPI, stabilizing the ED/LSI at moderate levels, and removing corridor pinch points. (iii) Place-based water–land coordination: Plots in zones with shallower groundwater and moderate precipitation supported higher diversity; maintaining environmental flows and safeguarding shallow-groundwater belts can widen riparian corridors and raise the PC/IIC without large land conversions. (iv) Scenario-informed zoning: The SSP projections indicate a persistent expansion of construction land; guiding this growth away from high-centrality links in the connectivity network and toward low-resistance areas could preserve basin-scale permeability under climate variability.
4.5.2. Limitations and Future Directions
First, the sample size (n = 57) may limit the model’s robustness. Future work should expand the spatial coverage using UAV high-resolution imagery and lightweight object-detection models (e.g., YOLOv10) [
63] coupled with Sentinel-2 data [
64] to automatically delineate vegetation patches and high-density species areas, and cross-validate against field plots to reduce sampling bias. Second, several socio-ecological covariates (e.g., population density, distance to roads) were excluded due to low measurement loadings (<0.5), potentially omitting environment–biodiversity pathways. Higher-resolution, process-proximal predictors (e.g., grazing intensity, groundwater extraction), together with variable transformations, hierarchical modeling, and multi-model sensitivity analyses should be used to test the robustness of human-pressure effects on the structure and connectivity. Lastly, our biodiversity metrics emphasize taxonomic diversity; integrating functional traits and phylogenetic diversity, ideally within a long-term monitoring design, will better capture ecosystem stability and adaptive capacity under climate stress.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}