
Aerobiological Dynamics and Climatic Sensitivity of Airborne Pollen in Southeastern Türkiye: A Two-Year Assessment from Siirt


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. The Study Area, Flora, and Climate Characteristics
2.2. Aerobiological Sampling and Statistical Analysis
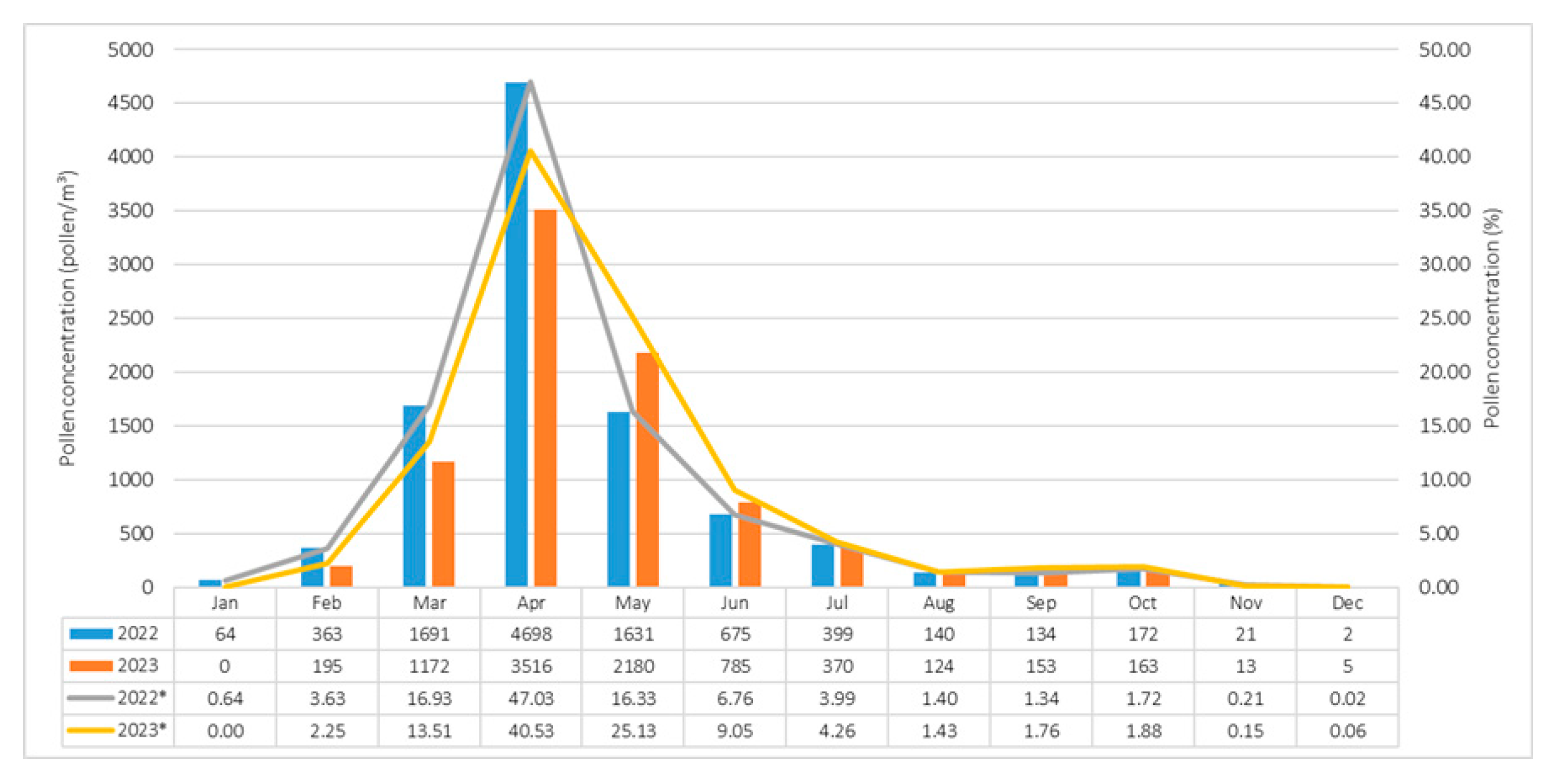
3. Results
3.1. Pollen Concentrations and Groups
3.2. Variations in Pollen Concentrations
3.3. Pollen Calendar
3.4. Dominant Pollen Taxa and MPS Periods
3.5. Meteorological Drivers of Daily Pollen Variability in Siirt (2022–2023)
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APIn | Annual Pollen Integral |
| CORINE | Coordination of Information on the Environment |
| MPS | Main Pollen Season |
References
- Scott, R.J.; Spielman, M.; Dickinson, H.G. Stamen structure and function. Plant Cell 2004, 16 (Suppl. S1), 46–60. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.; Mazer, S. Pollen-tiny and ephemeral but not forgotten: New ideas on their ecology and evolution. Am. J. Bot. 2016, 103, 365–374. [Google Scholar] [CrossRef]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell: Oxford, UK, 1991. [Google Scholar]
- Buters, J.T.M.; Antunes, C.; Galveias, A.; Bergmann, K.C.; Thibaudon, M.; Galán, C.; Schmidt-Weber, C.; Oteros, J. Pollen and spore monitoring in the world. Clin. Transl. Allergy 2018, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Liccardi, G.; Popov, T.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy 2007, 62, 976–990. [Google Scholar] [CrossRef]
- D’Amato, G.; Chong-Neto, H.J.; Ortega, O.P.M.; Vitale, C.; Ansotegui, I.; Rosario, N.; Haahtela, T.; Galan, C.; Pawankar, R.; Murrieta-Aguttes, M.; et al. The effects of climate change on respiratory allergy and asthma induced by pollen and mold allergens. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Traidl-Hoffmann, C. Pollen on their way astray—First contact via cross-kingdom signaling leading to far-reaching consequences for the atopic march. Allergy 2022, 77, 3496–3497. [Google Scholar] [CrossRef]
- Rodríguez-Rajo, F.; Méndez, J.; Jato, V. Factors affecting pollination ecology of Quercus anemophilous species in North-West Spain. Bot. J. Linn. Soc. 2005, 149, 283–297. [Google Scholar] [CrossRef]
- Negrini, A.; Negrini, S.; Giunta, V.; Quaglini, S.; Ciprandi, G. Thirty-year survey on airborne pollen concentrations in Genoa, Italy: Relationship with sensitizations, meteorological data, and air pollution. Am. J. Rhinol. Allergy 2011, 25, e232–e241. [Google Scholar] [CrossRef]
- Gregory, P.H. Distribution of airborne pollen and spores and their long distance transport. Pure Appl. Geophys. 1978, 116, 309–315. [Google Scholar] [CrossRef]
- Hedhly, A.; Hormaza, J.; Herrero, M. Influence of genotype-temperature interaction on pollen performance. J. Evol. Biol. 2005, 18, 1494–1502. [Google Scholar] [CrossRef]
- Jetschni, J.; Fritsch, M.; Jochner-Oette, S. How does pollen production of allergenic species differ between urban and rural environments? Int. J. Biometeorol. 2023, 67, 1839–1852. [Google Scholar] [CrossRef]
- Çeter, T.; Özler, H.; Pınar, N.M. First aeropalynological survey on the atmosphere of Sinop, Turkey. Kastamonu Univ. J. For. Fac. 2020, 20, 272–284. [Google Scholar] [CrossRef]
- Kızılpınar, İ.; Doğan, C.; Artaç, H.; Reisli, İ.; Pekcan, S. Pollen grains in the atmosphere of Konya (Turkey) and their relationship with meteorological factors, in 2008. Turk. J. Bot. 2012, 36, 344–357. [Google Scholar]
- Karadağ, G.E.A.; Altunoğlu, M.K. Airborne pollen seasonality of Kars province, a high-altitude region in NE Anatolia-Turkey. Palynology 2024, 49, 2382959. [Google Scholar] [CrossRef]
- Tosunoglu, A.; Saatcioglu, G.; Bekil, S.; Malyer, H.; Bicakci, A. Atmospheric pollen spectrum in Stone City, Mardin; the Northern Border of Mesopotamia/SE-Turkey. Environ. Monit. Assess. 2018, 190, 635. [Google Scholar] [CrossRef] [PubMed]
- Güvensen, A.; Öztürk, M. Airborne pollen calendar of Buca-İzmir, Turkey. Aerobiologia 2002, 18, 229–237. [Google Scholar] [CrossRef]
- Tosunoğlu, A.; İlçim, A.; Malyer, H.; Bıçakçı, A. Aeropalynological spectrum of Hatay, Turkey, the eastern coast of the Mediterranean Sea. Aerobiologia 2018, 34, 557–572. [Google Scholar] [CrossRef]
- Celenk, S.; Bicakci, A.; Tamay, Z.; Guler, N.; Altunoglu, M.K.; Canitez, Y.; Malyer, H.; Sapan, N.; Ones, U. Airborne pollen in European and Asian parts of Istanbul. Environ. Monit. Assess. 2010, 164, 391–402. [Google Scholar] [CrossRef]
- Bıçakçı, A.; Tosunoğlu, A. Allergenic pollen in Turkey. Asthma Allergy Immunol. 2019, 17, 140–151. [Google Scholar] [CrossRef]
- Pınar, S.M.; Fidan, M.; Eroğlu, H. Siirt ili florasına genel bir bakış. Commagene J. Biol. 2021, 5, 99–125. [Google Scholar] [CrossRef]
- Özyazıcı, M.A.; Dengiz, O.; İmamoğlu, A. Siirt ili bazı arazi ve toprak özelliklerinin coğrafi bilgi sistem analizleriyle değerlendirilmesi. Türkiye Tarımsal Araştırmalar Derg. 2014, 1, 128–137. [Google Scholar] [CrossRef]
- Mut, S. Comparative Land Use in the Central District of Siirt and Tillo District. Master’s Thesis, Karabük University, Karabük, Türkiye, 2020. [Google Scholar]
- Galán, C.; Cariñanos, P.; Alcázar, P.; Dominguez-Vilches, E. Spanish aerobiology network (REA) management andquality manual. Serv. Publicaciones Univ. Córdoba 2007, 184, 1–300. [Google Scholar]
- Andersen, T.B. A model to predict the beginning of the pollen season. Grana 1991, 30, 269–275. [Google Scholar] [CrossRef]
- de Weger, L.A.; Bergmann, K.C.; Rantio-Lehtimäki, A.; Dahl, Å.; Buters, J.; Déchamp, C.; Belmonte, J.; Thibaudon, M.; Cecchi, L.; Besancenot, J.P.; et al. Impact of pollen. In Allergenic Pollen: A Review of the Production, Release, Distribution and Health Impacts; Sofiev, M., Bergmann, K.-C., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 161–215. [Google Scholar]
- American Academy of Allergy, Asthma & Immunology (n.d.). AAAAI: The Global Leader in Allergy, Asthma, and Immunology. Available online: http://www.aaaai.org/ (accessed on 11 December 2024).
- Tasioulis, T.; Karatzas, K.; Charalampopoulos, A.; Damialis, A.; Vokou, D. Five ways to define a pollen season: Exploring congruence and disparity in its attributes and their long-term trends. Aerobiologia 2022, 38, 71–83. [Google Scholar] [CrossRef]
- Spieksma, F.T.M. Regional European pollen calendars. In Allergenic Pollen and Pollinosis in Europe; D’Amato, G., Spieksma, F.T.M., Bonini, S., Eds.; Blackwell Scientific Publications: Oxford, UK, 1991; pp. 49–65. [Google Scholar]
- Karabağ, M. Determination of Atmospheric Pollens in Ardahan Province, Posof District. Master’s Thesis, Kafkas University, Kars, Türkiye, 2023. [Google Scholar]
- Akpınar, S.; Altunoğlu, M.K. Determination of atmospheric pollen grains by volumetric method in Sarıkamış District (Kars-Türkiye). Biology 2024, 13, 475. [Google Scholar] [CrossRef] [PubMed]
- Gucel, S.; Guvensen, A.; Ozturk, M.; Çelik, A. Analysis of airborne pollen fall in Nicosia (Cyprus). Environ. Monit. Assess. 2013, 185, 157–169. [Google Scholar] [CrossRef]
- Rizzi-Longo, L.; Pizzulin-Sauli, M.; Stravisi, F.; Ganis, P. Airborne pollen calendar for Trieste (Italy), 1990–2004. Grana 2007, 46, 98–109. [Google Scholar] [CrossRef]
- Calderón-Ezquerro, M.C.; Guerrero-Guerra, C.; Martínez-López, B.; Fuentes-Rojas, F.; Téllez-Unzueta, F.; López-Espinoza, E.D.; Calderón-Segura, M.E.; Martínez-Arroyo, A.; Trigo-Pérez, M.M. First airborne pollen calendar for Mexico City and its relationship with bioclimatic factors. Aerobiologia 2016, 32, 225–244. [Google Scholar] [CrossRef]
- Camacho, I.C. Airborne pollen in Funchal City, (Madeira Island, Portugal)-First pollinic calendar and allergic risk assessment. Ann. Agric. Environ. Med. 2015, 22, 608–613. [Google Scholar] [CrossRef]
- Jiang, K.; Du, C.; Huang, L.; Luo, J.; Liu, T.; Huang, S. Phylotranscriptomics and evolution of key genes for terpene biosynthesis in Pinaceae. Front. Plant Sci. 2023, 14, 1114579. [Google Scholar] [CrossRef]
- He, T.; Belcher, C.; Lamont, B.; Lim, S. A 350-million-year legacy of fire adaptation among conifers. J. Ecol. 2015, 104, 352–363. [Google Scholar] [CrossRef]
- Bowe, L.M.; Coat, G.; de Pamphilis, C.W. Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales’ closest relatives are conifers. Proc. Natl. Acad. Sci. USA 2000, 97, 4092–4097. [Google Scholar] [CrossRef] [PubMed]
- Krutovsky, K.V.; Troggio, M.; Brown, G.R.; Jermstad, K.D.; Neale, D.B. Comparative mapping in the Pinaceae. Genetics 2004, 168, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Ortega, J.; López-Matas, M.; Alonso, M.; Felíu, A.; Ruiz-Hornillos, J.; González, E.; Moya, R.; Carnés, J. Prevalence of allergic sensitization to conifer pollen in a high Cypress exposure area. Allergy Rhinol. 2016, 7, ar-2016. [Google Scholar] [CrossRef]
- Erkara, İ.; Cingi, C.; Ayrancı, Ü.; Gürbüz, K.; Pehlivan, S.; Tokur, S. Skin prick test reactivity in allergic rhinitis patients to airborne pollens. Environ. Monit. Assess. 2008, 151, 401–412. [Google Scholar] [CrossRef]
- García-Gallardo, M.V.; Algorta, J.; Longo, N.; Espinel, S.; Aragones, A.; Lombardero, M.; Bernaola, G.; Jauregui, I.; Aranzabal, A.; Albizu, M.V.; et al. Evaluation of the effect of pollution and fungal disease on Pinus radiata pollen allergenicity. Int. Arch. Allergy Immunol. 2013, 160, 241–250. [Google Scholar] [CrossRef]
- Bicakci, A.; Tosunoglu, A.; Altunoglu, M.K.; Saatcioglu, G.; Keser, A.M.; Ozgokce, F. An aeropalynological survey in the city of Van, a high altitudinal region, East Anatolia-Turkey. Aerobiologia 2017, 33, 93–108. [Google Scholar] [CrossRef]
- Kilic, M.; Altunoglu, M.K.; Akpınar, S.; Akdogan, G.E.; Taskin, E. Relationship between airborne pollen and skin prick test results in Elazığ, Turkey. Aerobiologia 2019, 35, 593–604. [Google Scholar] [CrossRef]
- Kafashan, H.A.; Khosravi, A.R.; Alyasin, S.; Sepahi, N.; Kanannejad, Z.; Shirazi, F.M.A.Z.; Karami, S. Airborne pollens and their association with meteorological parameters in the atmosphere of Shiraz, Southwest Iran. Iran. J. Allergy Asthma Immunol. 2021, 20, 294–302. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Z.; Ferguson, D.K.; Shong, J.Y. An integrative view on the systematic position of the cupressophyte Cephalotaxus. Ecol. Evol. 2023, 13, e10273. [Google Scholar] [CrossRef]
- Ghimire, B.; Jeong, M.; Lee, C.; Heo, K. Inclusion of Cephalotaxus in Taxaceae: Evidence from morphology and anatomy. Korean J. Pl. Taxon. 2018, 48, 109–114. [Google Scholar] [CrossRef]
- Quinn, C.; Price, R.; Gadek, P. Familial concepts and relationships in the conifer based on rbcL and matK sequence comparisons. Kew Bull. 2002, 57, 513–531. [Google Scholar] [CrossRef]
- Majeed, A.; Singh, A.; Choudhary, S.; Bhardwaj, P. RNAseq-based phylogenetic reconstruction of Taxaceae and Cephalotaxaceae. Cladistics 2019, 35, 461–468. [Google Scholar] [CrossRef]
- Sundararaj, R.; Mathimaran, A.; Prabhu, D.; Ramachandran, B.; Jeyaraman, J.; Muthupandian, S.; Asmelash, T. In silico approaches for the identification of potential allergens among hypothetical proteins from Alternaria alternata and its functional annotation. Sci. Rep. 2024, 14, 6696. [Google Scholar] [CrossRef]
- Galveias, A.; Costa, A.R.; Bortoli, D.; Alpizar-Jara, R.; Salgado, R.; Costa, M.J.; Antunes, C.M. Cupressaceae pollen in the city of Évora, South of Portugal: Disruption of the pollen during air transport facilitates allergen exposure. Forests 2021, 12, 64. [Google Scholar] [CrossRef]
- Charpin, D.; Pichot, C.; Belmonte, J.; Sutra, J.P.; Zidkova, J.; Chavez, P.; Shahali, Y.; Senéchal, H.; Poncet, P. Cypress pollinosis: From tree to clinic. Chin. Rev. Allergy Immunol. 2017, 54, 174–195. [Google Scholar] [CrossRef]
- Yoo, K.H.; Kwon, T.R.; Kim, Y.U.; Kim, E.H.; Kim, B.J. The effects of fabric containing Chamaecyparis obtusa essential oil on atopic dermatitis-like lesions: A functional clothing possibility. Ski. Pharmacol. Physiol. 2020, 33, 142–152. [Google Scholar] [CrossRef]
- Majrashi, A.A. Preliminary assessment of weed population in vegetable and fruit farms of Taif, Saudi Arabia. Braz. J. Biol. 2022, 82, e255816. [Google Scholar] [CrossRef]
- Grass Phylogeny Working Group III; Arthan, W.; Baker, W.J.; Barrett, M.D.; Barrett, R.L.; Bennetzen, J.; Besnard, G.; Bianconi, M.E.; Birch, J.L.; Catalán, P.; et al. Nuclear phylogenomics of grasses (Poaceae) supports current classification and reveals repeated reticulation. bioRxiv 2024, 5, 1–33. [Google Scholar] [CrossRef]
- Adachukwu, O.; Kenneth, E.; Chukwu, O.; Adaugo, N.; Chisom, I. Ecological survey on species of Poaceae family present in Nnamdi Azikiwe University campus Awka, Anambra State. Asian J. Environ. Ecol. 2023, 21, 34–42. [Google Scholar] [CrossRef]
- Céccoli, G.; Ramos, J.; Pilatti, V.; Dellaferrera, I.; Tivano, J.C.; Taleisnik, E.; Vegetti, A.C. Salt glands in the Poaceae family and their relationship to salinity tolerance. Bot. Rev. 2015, 81, 162–178. [Google Scholar] [CrossRef]
- Xu, X.; Dimitrov, D.; Shrestha, N.; Rahbek, C.; Wang, Z. Sensitization profiles of timothy grass pollen in Northern China. J. Asthma Allergy 2021, 14, 1431–1439. [Google Scholar] [CrossRef]
- Li, J.D.; Gu, J.Q.; Xu, Y.Y.; Cui, L.; Li, L.S.; Wang, Z.X.; Yin, J.; Guan, K. Serum IgE profiles in Chinese pollinosis patients with grass pollen sensitization. World Allergy Organ. J. 2022, 15, 100624. [Google Scholar] [CrossRef]
- Mohamed, M.; Refaat, M.; Melek, N.; Ahmed, E.; Aldin, N.; Latif, O. Pollen sensitization among Egyptian patients with respiratory allergic diseases. Egypt. J. Immunol. 2022, 29, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Puc, M. Threat of allergenic airborne grass pollen in Szczecin, Nw Poland: The dynamics of pollen seasons, effect of meteorological variables and air pollution. Aerobiologia 2010, 27, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Kmenta, M.; Bastl, K.; Berger, U.; Kramer, M.F.; Heath, M.D.; Pätsi, S.; Pessi, A.-M.; Saarto, A.; Werchan, B.; Werchan, M.; et al. The grass pollen season 2015: A proof of concept multi-approach study in three different European cities. World Allergy Organ. J. 2017, 10, 31. [Google Scholar] [CrossRef]
- Celenk, S.; Bicakci, A. Aerobiological investigation in Bitlis, Turkey. Ann. Agric. Environ. Med. 2005, 12, 87–93. [Google Scholar]
- Rodríguez-de la Cruz, D.; Sánchez-Reyes, E.; Dávila-González, I.; Lorente-Toledano, F.; Sánchez-Sánchez, J. Airborne pollen calendar of Salamanca, Spain, 2000–2007. Allergol. Immunopathol. 2010, 38, 307–312. [Google Scholar] [CrossRef]
- Clement, W.L.; Weiblen, G.D. Morphological evolution in the Mulberry family (Moraceae). Syst. Bot. 2009, 34, 530–552. [Google Scholar] [CrossRef]
- Aneklaphakij, C.; Bunsupa, S.; Sirichamorn, Y.; Bongcheewin, B.; Satitpatipan, V. Taxonomic notes on the ‘Mahat’ (Artocarpus lacucha and A. thailandicus, Moraceae) species complex in Thailand. Plants 2020, 9, 391. [Google Scholar]
- Papia, F.; Incorvaia, C.; Genovese, L.; Gangemi, S.; Minciullo, P.L. Allergic reactions to genus Morus plants: A review. Clin. Mol. Allergy 2020, 18, 1. [Google Scholar] [CrossRef]
- Ahmed, F. Ficus benghalensis bark extract shows blood pressure lowering effect in normotensive and angiotensin II-induced hypertensive rats. Pharmacophore 2021, 12, 7–10. [Google Scholar] [CrossRef]
- Li, J.D.; Du, Z.R.; Liu, J.; Xu, Y.Y.; Wang, R.Q.; Yin, J. Characteristics of pollen-related food allergy based on individual pollen allergy profiles in the Chinese population. World Allergy Organ. J. 2020, 13, 100120. [Google Scholar] [CrossRef] [PubMed]
- Nitiu, D.S. Aeropalynologic analysis of La Plata City (Argentina) during a 3-year period. Aerobiologia 2006, 22, 79–87. [Google Scholar] [CrossRef]
- Tosunoğlu, A.; Bıçakçı, A.; Malyer, H.; Sapan, N. Analysis of airborne pollen fall in Köyceğiz specialty protected area (SW Turkey). Fresenius Environ. Bull. 2009, 18, 1860–1865. [Google Scholar]
- Gharbi, D.; Al-Nesf, M.; Trigo, M. Do we need aerobiological air monitoring in desert climates? The Qatar experience. Qatar Med. J. 2022, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska-Weryszko, K.; Weryszko-Chmielewska, E.; Sulborska, A.; Puc, M.; Malkiewicz, M.; Siergiejko, Z.; Dąbrowska-Zapart, K.; Ziemianin, M.; Rapiejko, A.; Wieczorkiewicz, A.; et al. Concentration of pollen of Chenopodiaceae/Amaranthaceae plants in the air of selected Polish cities in 2020. Alergoprofil 2020, 16, 34–40. [Google Scholar] [CrossRef]
- Gonçalves, F.; Sousa, A.; Oliveira, R. Use of mass spectrometry as a tool for the search or identification of flavonoids in Urticaceae. Rodriguésia 2023, 74, e01152022. [Google Scholar] [CrossRef]
- Arilla, M.C.; González-Rioja, R.; Ibarrola, I.; Mir, A.; Monteseirín, J.; Conde, J.; Martínez, A.; Asturias, J.A. A sensitive monoclonal antibody-based enzyme-linked ımmunosorbent assay to quantify Parietaria judaica major allergens, Par j 1 and Par j 2. Clin. Exp. Allergy 2006, 36, 87–93. [Google Scholar] [CrossRef]
- Cavender-Bares, J. Diversification, adaptation, and community assembly of the American oaks (Quercus), a model clade for integrating ecology and evolution. New Phytol. 2018, 221, 669–692. [Google Scholar] [CrossRef]
- Hipp, A.L.; Manos, P.S.; Hahn, M.; Avishai, M.; Bodénès, C.; Cavender-Bares, J.; Crowl, A.A.; Deng, M.; Denk, T.; Fitz-Gibbon, S.; et al. Genomic landscape of the global oak phylogeny. New Phytol. 2020, 226, 1198–1212. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-J.; Li, Y.-X.; Li, N.; Zhu, H.-T.; Wang, D.; Zhang, Y.-J. The genus Rumex (Polygonaceae): An ethnobotanical, phytochemical and pharmacological review. Nat. Prod. Bioprospect. 2022, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Tommaso, M.; Luciani, A.; Crisi, P.; Beschi, M.; Rosi, P.; Rocconi, F.; Miglio, A. Detection of serum allergen-specific ige in atopic dogs tested in Northern Italy: Preliminary study. Animals 2021, 11, 358. [Google Scholar] [CrossRef]
- Tosunoglu, A.; Altunoglu, M.K.; Bicakci, A.; Kilic, O.; Gonca, T.; Yilmazer, I.; Saatcioglu, G.; Akkaya, A.; Celenk, S.; Canitez, Y.; et al. Atmospheric pollen concentrations in Antalya, South Turkey. Aerobiologia 2015, 31, 99–109. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Abatzoglou, J.T.; Anderegg, L.D.L.; Bielory, L.; Kinney, P.L.; Ziska, L. Anthropogenic climate change is worsening North American pollen seasons. Proc. Natl. Acad. Sci. USA 2021, 118, e2013284118. [Google Scholar] [CrossRef]
- Ziska, L.; Knowlton, K.; Rogers, C.; Dalan, D.; Tierney, N.; Elder, M.A.; Filley, W.; Shropshire, J.; Ford, L.B.; Hedberg, C.; et al. Recent warming by latitude associated with increased length of ragweed pollen season in central North America. Proc. Natl. Acad. Sci. USA 2011, 108, 4248–4251. [Google Scholar] [CrossRef] [PubMed]
- Harbele, S.G.; Bowman, D.M.J.S.; Newham, R.M.; Johnston, F.H.; Beggs, P.J.; Buters, J.; Campbell, B.; Erbas, B.; Godwin, I.; Green, B.J.; et al. The macroecology of airborne pollen in Australia and New Zealand urban areas. PLoS ONE 2014, 9, e97925. [Google Scholar]
- Veriankaitė, L.; Šaulienė, I.; Bukantis, A. Evaluation of meteorological parameters influence upon pollen spread in the atmosphere. J. Environ. Eng. Landsc. 2011, 19, 5–11. [Google Scholar] [CrossRef][Green Version]
- Izquierdo, R.; Belmonte, J.; Avila, A.; Alarcón, M.; Cuevas, E.; Alonso-Pérez, S. Source areas and long-range transport of pollen from continental land to Tenerife (Canary Islands). Int. J. Biometeorol. 2011, 55, 67–85. [Google Scholar] [CrossRef]
- Frei, T.; Gassner, E. Climate change and its impact on birch pollen quantities and the start of the pollen season: An example from Switzerland for the period 1969–2006. Int. J. Biometeorol. 2008, 52, 667–674. [Google Scholar] [CrossRef]
- Bortenschlager, S.; Bortenschlager, I. Altering airborne pollen concentrations due to global warming: A comparative analysis of airborne pollen records from Innsbruck and Obergurgl (Austria) for the period 1980–2001. Grana 2005, 44, 172–180. [Google Scholar] [CrossRef]
- Yli-Panula, E.; Fekedulegn, D.B.; Green, B.J.; Ranta, H. Analysis of airborne Betula pollen in Finland: A 31-year perspective. Int. J. Environ. Res. Public Health 2009, 6, 1706–1723. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | 2022 | 2023 | Mean | |||
|---|---|---|---|---|---|---|
| Total (p/m3) | % | Total (p/m3) | % | Total (p/m3) | % | |
| Pinaceae | 3394 | 33.97 | 2392 | 27.57 | 2893 | 31.00 |
| Cupressaceae/Taxaceae | 2932 | 29.35 | 2255 | 25.99 | 2594 | 27.79 |
| Moraceae | 434 | 4.34 | 356 | 4.10 | 395 | 4.23 |
| Quercus | 169 | 1.69 | 120 | 1.38 | 145 | 1.55 |
| Platanus | 84 | 0.84 | 89 | 1.03 | 87 | 0.93 |
| Acer | 70 | 0.70 | 85 | 0.98 | 78 | 0.83 |
| Fraxinus | 80 | 0.80 | 72 | 0.83 | 76 | 0.81 |
| Rosaceae | 66 | 0.66 | 67 | 0.77 | 67 | 0.71 |
| Pistacia | 50 | 0.50 | 62 | 0.71 | 56 | 0.60 |
| Salix | 27 | 0.27 | 74 | 0.85 | 51 | 0.54 |
| Olea | 44 | 0.44 | 49 | 0.56 | 47 | 0.50 |
| Juglans | 41 | 0.41 | 51 | 0.59 | 46 | 0.49 |
| Ligustrum | 21 | 0.21 | 26 | 0.30 | 24 | 0.25 |
| Populus | 23 | 0.23 | 18 | 0.21 | 21 | 0.22 |
| Betula | 8 | 0.08 | 8 | 0.09 | 8 | 0.09 |
| Alnus | 7 | 0.07 | 7 | 0.08 | 7 | 0.08 |
| Ericaceae | 1 | 0.01 | 3 | 0.03 | 2 | 0.02 |
| Tilia | 1 | 0.01 | 3 | 0.03 | 2 | 0.02 |
| Fagus | 1 | 0.01 | - | - | 1 | 0.01 |
| Ulmus | 1 | 0.01 | - | - | 1 | 0.01 |
| Woody taxa | 7454 | 74.61 | 5737 | 66.12 | 6596 | 70.67 |
| Poaceae | 1519 | 15.21 | 1920 | 22.13 | 1720 | 18.42 |
| Amaranthaceae | 207 | 2.07 | 245 | 2.82 | 226 | 2.42 |
| Urticaceae | 206 | 2.06 | 191 | 2.20 | 199 | 2.13 |
| Fabaceae | 130 | 1.30 | 110 | 1.27 | 120 | 1.29 |
| Rumex | 93 | 0.93 | 97 | 1.12 | 95 | 1.02 |
| Taraxacum | 65 | 0.65 | 90 | 1.04 | 78 | 0.83 |
| Boraginaceae | 51 | 0.51 | 49 | 0.56 | 50 | 0.54 |
| Asteraceae | 44 | 0.44 | 45 | 0.52 | 45 | 0.48 |
| Artemisia | 29 | 0.29 | 56 | 0.65 | 43 | 0.46 |
| Mercurialis | 54 | 0.54 | 24 | 0.28 | 39 | 0.42 |
| Apiaceae | 36 | 0.36 | 38 | 0.44 | 37 | 0.40 |
| Plantago | 25 | 0.25 | 27 | 0.31 | 26 | 0.28 |
| Brassicaceae | 30 | 0.30 | 13 | 0.15 | 22 | 0.23 |
| Lamiaceae | 22 | 0.22 | 16 | 0.18 | 19 | 0.20 |
| Cyperaceae | 16 | 0.16 | 9 | 0.10 | 13 | 0.13 |
| Xanthium | 6 | 0.06 | 6 | 0.07 | 6 | 0.06 |
| Caryophyllaceae | 3 | 0.03 | 3 | 0.03 | 3 | 0.03 |
| Herbaceous taxa | 2536 | 25.39 | 2939 | 33.88 | 2738 | 29.33 |
| Total | 9990 | 100.00 | 8676 | 100.00 | 9333 | 100.00 |
| Taxa | 2022 | 2023 | |
|---|---|---|---|
| Amaranthaceae | MPS | 27 May–26 October | 24 May–27 October |
| Season length (days) | 152 | 156 | |
| Maximum pollen/day | 5 pollen/m3—23 September | 5 pollen/m3—18 September | |
| Cupressaceae/Taxaceae | MPS | 20 February–26 May | 24 February–31 May |
| Season length (days) | 95 | 96 | |
| Maximum pollen/day | 177 pollen/m3—20 February | 134 pollen/m3—13 March | |
| Fabaceae | MPS | 19 May–2 October | 30 April–18 September |
| Season length (days) | 136 | 141 | |
| Maximum pollen/day | 5 pollen/m3—25 June | 7 pollen/m3—18 June | |
| Moraceae | MPS | 5 April–21 May | 8 April–29 April |
| Season length (days) | 46 | 21 | |
| Maximum pollen/day | 74 pollen/m3—17 April | 56 pollen/m3—21 April | |
| Pinaceae | MPS | 1 April–30 Jun | 5 April–7 Aug |
| Season length (days) | 90 | 124 | |
| Maximum pollen/day | 484 pollen/m3—3 April | 395 pollen/m3—7 April | |
| Poaceae | MPS | 12 April–16 October | 18 April–9 October |
| Season length (days) | 187 | 174 | |
| Maximum pollen/day | 69 pollen/m3—17 May | 87 pollen/m3—17 May | |
| Quercus | MPS | 31 March–24 May | 4 April–29 May |
| Season length (days) | 54 | 55 | |
| Maximum pollen/day | 12 pollen/m3—16 May | 10 pollen/m3—22 April | |
| Rumex | MPS | 25 March–20 July | 29 March–8 June |
| Season length (days) | 117 | 71 | |
| Maximum pollen/day | 5 pollen/m3—28 May | 7 pollen/m3—23 May | |
| Urticaceae | MPS | 26 March–30 July | 30 March–3 June |
| Season length (days) | 126 | 65 | |
| Maximum pollen/day | 8 pollen/m3—27 May | 10 pollen/m3—20 May |
| Mean Daily Temperature °C | Mean Daily Relative Humidity (%) | Mean Daily Wind Speed (m/s) | Daily Rainfall (mm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2022 | 2023 | MPS Periods | 2022 | 2023 | MPS Periods | 2022 | 2023 | MPS Periods | 2022 | 2023 | MPS Periods | |
| Amarantaceae | −0.161 | −0.008 | −0.160 | 0.171 | −0.028 | 0.189 * | −0.001 | −0.177 * | −0.152 | 0.363 | 0.334 | 0.253 |
| Cupressaceae/Tax. | −0.206 * | −0.274 ** | −0.211 * | 0.197 * | 0.339 ** | −0.005 | −0.142 | −0.054 | −0.096 | −0.033 | −0.349 * | −0.022 |
| Fabaceae | −0.176 | 0.016 | −0.130 | 0.185 | −0.041 | 0.143 | 0.100 | 0.113 | 0.144 | −0.211 | 0.083 | −0.293 |
| Moraceae | 0.186 | 0.305 | 0.131 | −0.370 ** | −0.176 | −0.363 * | −0.294 * | 0.031 | −0.282 | −0.480 | −0.165 | −0.160 |
| Pinaceae | −0.101 | −0.157 * | −0.799 ** | 0.123 | 0.163 * | 0.766 ** | 0.024 | 0.296 ** | −0.095 | −0.181 | −0.363 * | −0.048 |
| Poaceae | −0.014 | −0.096 | −0.336 ** | 0.077 | 0.133 | 0.405 ** | 0.299 ** | 0.336 ** | 0.322 ** | −0.141 | −0.222 | −0.116 |
| Quercus | 0.163 | −0.059 | 0.232 | 0.085 | −0.094 | −0.128 | 0.012 | −0.028 | 0.029 | −0.106 | −0.466 | −0.021 |
| Rumex | 0.011 | 0.330 * | 0.114 | 0.010 | −0.225 | −0.052 | −0.146 | 0.247 | −0.016 | −0.423 | −0.197 | −0.086 |
| Urticaceae | −0.198 | 0.179 | −0.399 ** | 0.092 | −0.058 | 0.381 ** | −0.085 | −0.086 | −0.185 | −0.161 | −0.076 | 0.064 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akpınar, S. Aerobiological Dynamics and Climatic Sensitivity of Airborne Pollen in Southeastern Türkiye: A Two-Year Assessment from Siirt. Biology 2025, 14, 841. https://doi.org/10.3390/biology14070841
Akpınar S. Aerobiological Dynamics and Climatic Sensitivity of Airborne Pollen in Southeastern Türkiye: A Two-Year Assessment from Siirt. Biology. 2025; 14(7):841. https://doi.org/10.3390/biology14070841
Chicago/Turabian StyleAkpınar, Salih. 2025. "Aerobiological Dynamics and Climatic Sensitivity of Airborne Pollen in Southeastern Türkiye: A Two-Year Assessment from Siirt" Biology 14, no. 7: 841. https://doi.org/10.3390/biology14070841
APA StyleAkpınar, S. (2025). Aerobiological Dynamics and Climatic Sensitivity of Airborne Pollen in Southeastern Türkiye: A Two-Year Assessment from Siirt. Biology, 14(7), 841. https://doi.org/10.3390/biology14070841