
The Regulatory Role of EvfG Through Coordinated Control of Flagellar Biosynthesis and Energy Metabolism in Porcine Extraintestinal Pathogenic Escherichia coli (ExPEC)

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. Bacteria Motility Assays
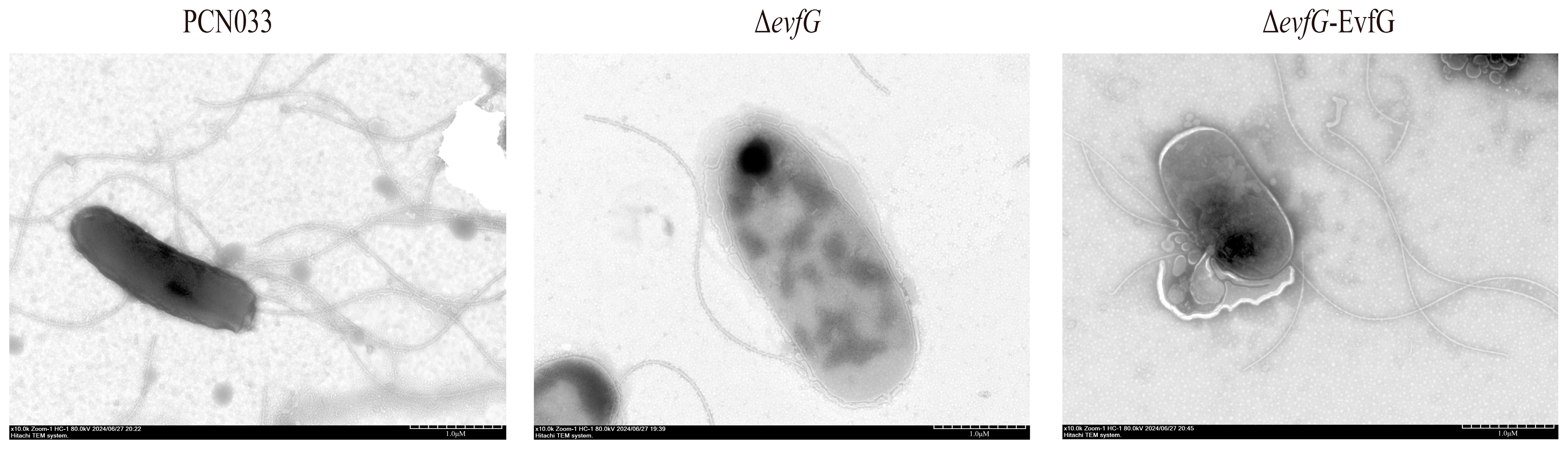
2.3. Bacterial Flagella Visualization with Transmission Electron Microscopy (TEM)
2.4. Total RNA Extraction, Library Building, and Transcriptome Sequencing
2.5. Transcriptome Data Analysis
2.6. Metabolite Extraction
2.7. Metabolite Extract LC-MS and Quality Control (QC) Measurements
2.8. Metabolite Data Analysis
2.9. Bacterial RNA Extraction and qRT-PCR Analysis
2.10. Statistical Analysis
3. Results
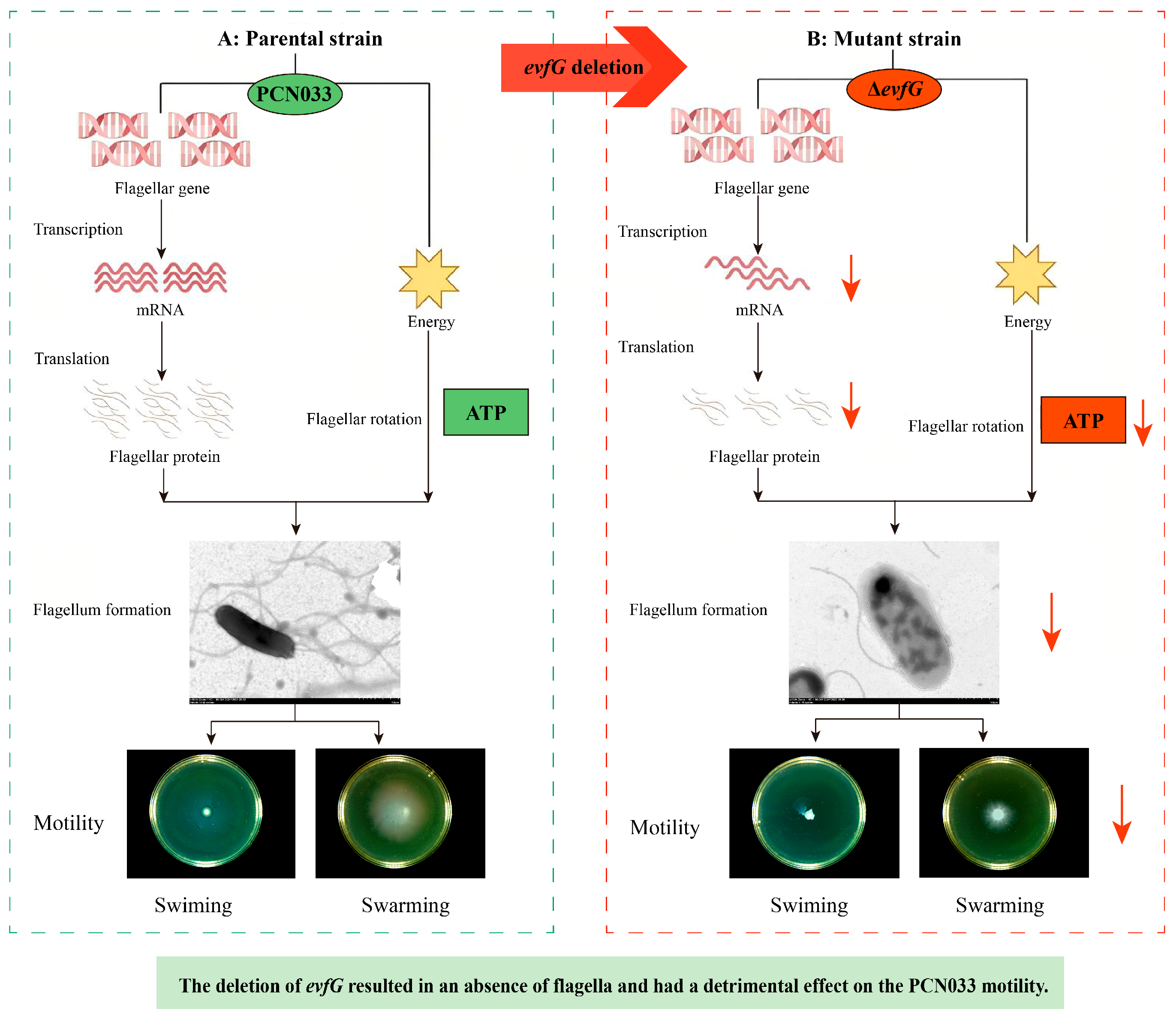
3.1. The Deletion of Gene evfG Affects Bacterial Swimming and Swarming
3.2. Deletion of evfG Affects the Flagellum Formation in PCN033
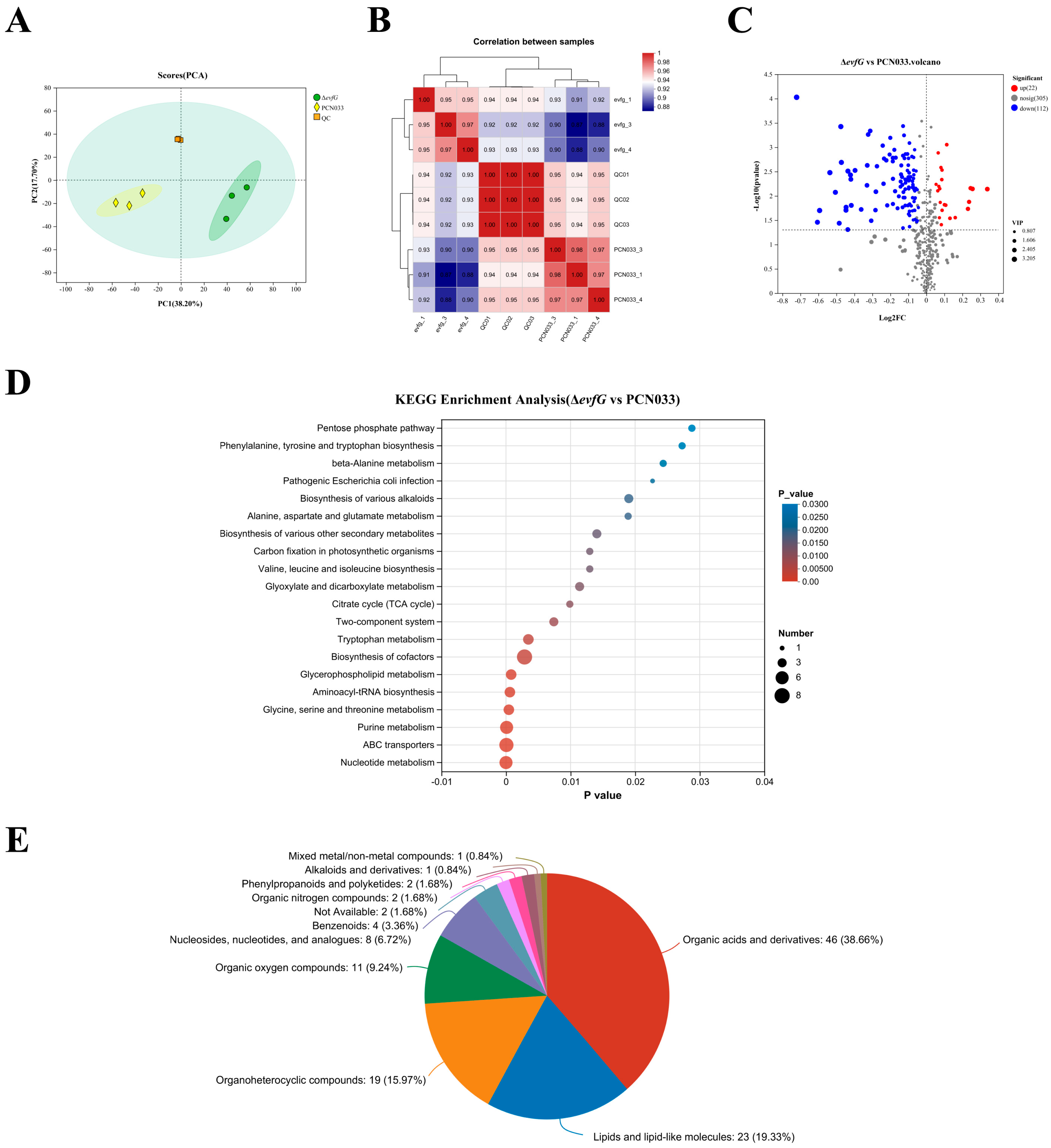
3.3. Metabolome Profiling
3.4. Differentially Accumulated Metabolites (DAMs) in PCN033 and ΔevfG
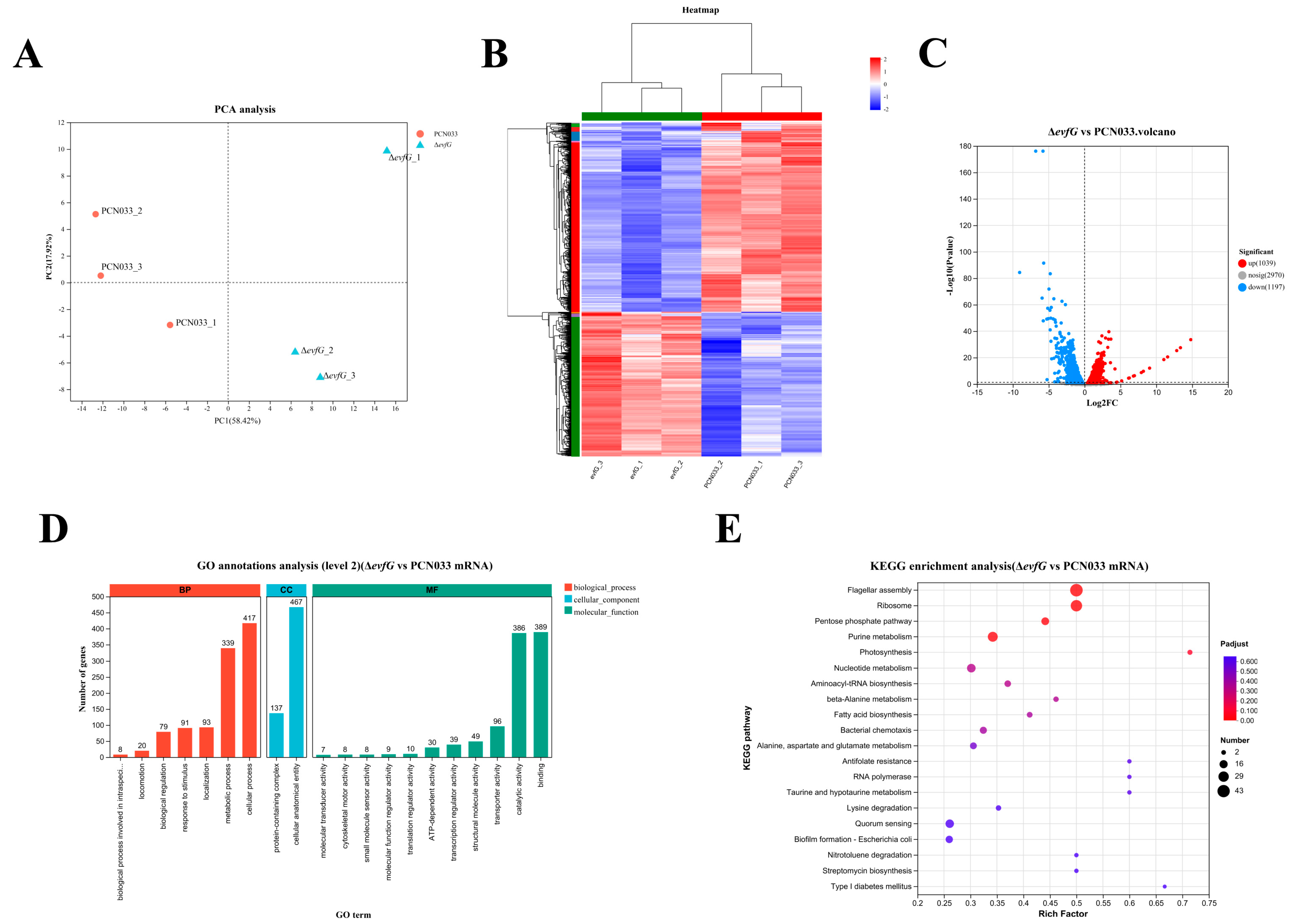
3.5. Transcriptome Profiles Between PCN033 and ΔevfG
3.6. DEGs Between PCN033 and ΔevfG
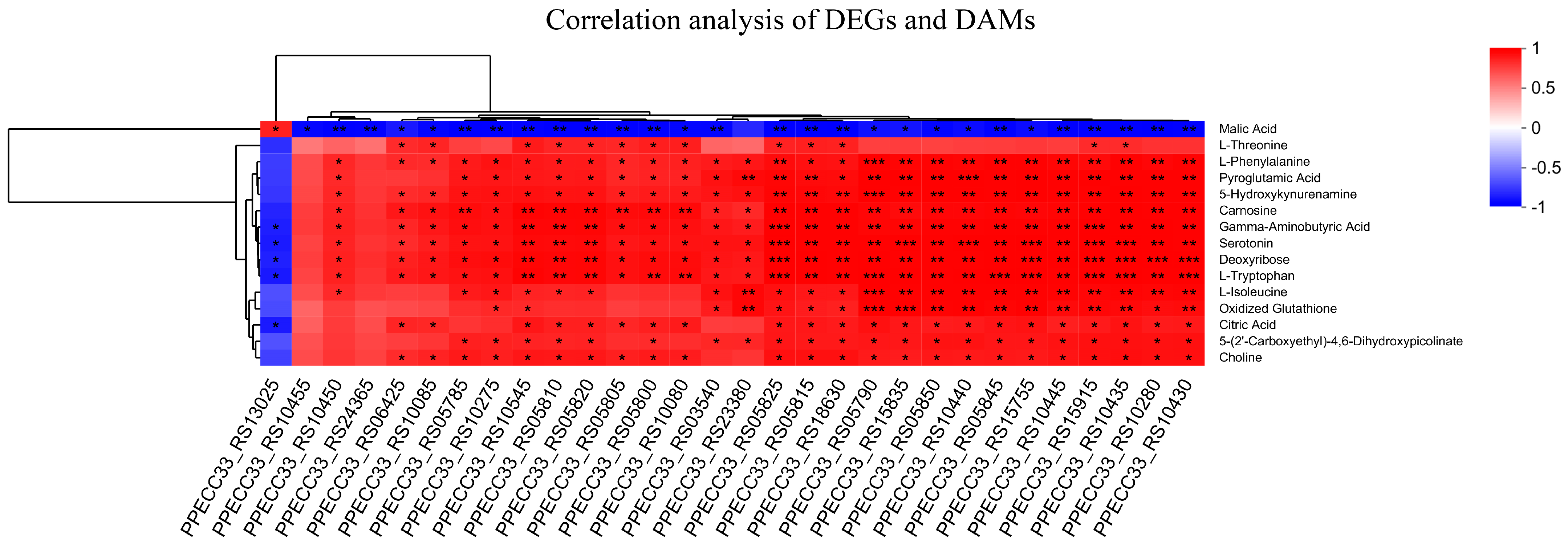
3.7. The Correlation Analysis of DAMs and DEGs
4. Discussion
4.1. EvfG Is Essential for the Motility of Porcine ExPEC PCN033
4.2. The Reduced Motility of the Mutant ΔevfG Is Related to the Energy Metabolism
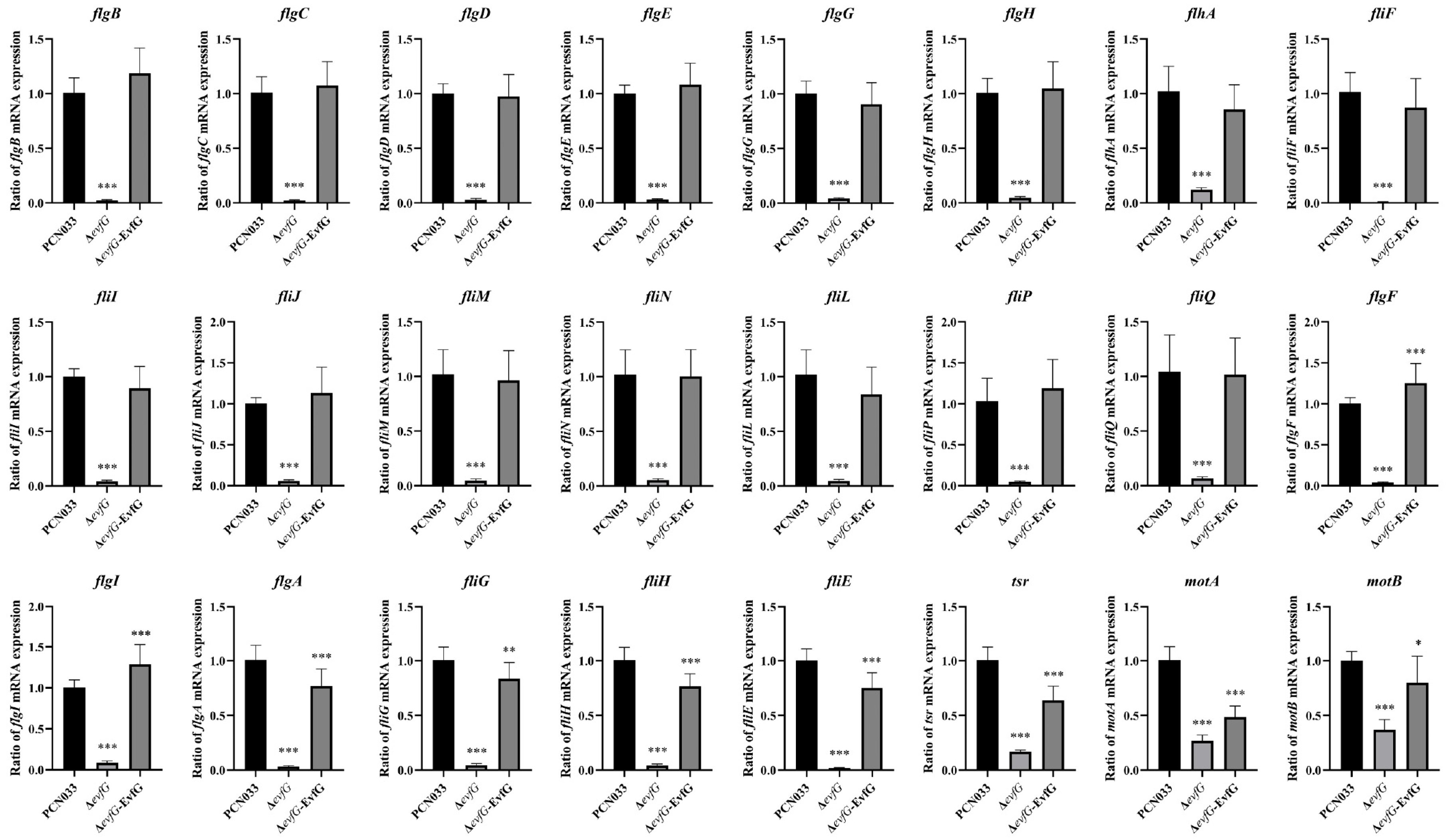
4.3. The Reduced Motility of the Mutant ΔevfG Was Associated with the Loss of Flagellar-Associated Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weston, E.J.; Pondo, T.; Lewis, M.M.; Martell-Cleary, P.; Morin, C.; Jewell, B.; Daily, P.; Apostol, M.; Petit, S.; Farley, M.; et al. The burden of invasive early-onset neonatal sepsis in the United States, 2005–2008. Pediatr. Infect. Dis. J. 2011, 30, 937–941. [Google Scholar] [CrossRef] [PubMed]
- Mellata, M. Human and Avian Extraintestinal Pathogenic Escherichia coli: Infections, Zoonotic Risks, and Antibiotic Resistance Trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef]
- Huang, S.; Chi, F.; Peng, L.; Bo, T.; Zhang, B.; Liu, L.; Wu, X.; Mor-Vaknin, N.; Markovitz, D.M.; Cao, H.; et al. Vimentin, a Novel NF-κB Regulator, Is Required for Meningitic Escherichia coli K1-Induced Pathogen Invasion and PMN Transmigration across the Blood-Brain Barrier. PLoS ONE 2016, 11, e0162641. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Tang, X.; Zhang, X.; Ding, Y.; Zhao, Z.; Wu, B.; Cai, X.; Liu, Z.; He, Q.; Chen, H. Serotypes and virulence genes of extraintestinal pathogenic Escherichia coli isolates from diseased pigs in China. Vet. J. 2011, 192, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Tan, C.; Zhang, X.; Zhao, Z.; Xia, X.; Wu, B.; Guo, A.; Zhou, R.; Chen, H. Antimicrobial resistances of extraintestinal pathogenic Escherichia coli isolates from swine in China. Microb. Pathog. 2011, 50, 207–212. [Google Scholar] [CrossRef]
- Jahanbakhsh, S.; Smith, M.G.; Kohan-Ghadr, H.; Letellier, A.; Abraham, S.; Trott, D.J.; Fairbrother, J.M. Dynamics of extended-spectrum cephalosporin resistance in pathogenic Escherichia coli isolated from diseased pigs in Quebec, Canada. Int. J. Antimicrob. Agents 2016, 48, 194–202. [Google Scholar] [CrossRef]
- Moulin-Schouleur, M.; Schouler, C.; Tailliez, P.; Kao, M.; BréE, A.; Germon, P.; Oswald, E.; Mainil, J.; Blanco, M.; Blanco, J. Common Virulence Factors and Genetic Relationships between O18:K1:H7 Escherichia coli Isolates of Human and Avian Origin. J. Clin. Microbiol. 2006, 44, 3484–3492. [Google Scholar] [CrossRef]
- Zhu, Y.; Dong, W.; Ma, J.; Yuan, L.; Hejair, H.M.; Pan, Z.; Liu, G.; Yao, H. Characterization and virulence clustering analysis of extraintestinal pathogenic Escherichia coli isolated from swine in China. BMC Vet. Res. 2017, 13, 94. [Google Scholar] [CrossRef]
- Biran, D.; Ron, E.Z. Extraintestinal Pathogenic Escherichia coli. In Escherichia coli, a Versatile Pathogen; Current Topics in Microbiology and Immunology; Springer: Cham, Switzerland, 2018; pp. 149–161. [Google Scholar] [CrossRef]
- Li, X.; Hu, H.; Zhu, Y.; Wang, T.; Lu, Y.; Wang, X.; Peng, Z.; Sun, M.; Chen, H.; Zheng, J.; et al. Population structure and antibiotic resistance of swine extraintestinal pathogenic Escherichia coli from China. Nat. Commun. 2024, 15, 5811. [Google Scholar] [CrossRef]
- Das, S.; Chaudhuri, K. Identification of a unique IAHP (IcmF associated homologous proteins) cluster in Vibrio cholerae and other proteobacteria through in silico analysis. In Silico Biol. 2003, 3, 287–300. [Google Scholar]
- Mougous, J.D.; Cuff, M.E.; Raunser, S.; Shen, A.; Zhou, M.; Gifford, C.A.; Goodman, A.L.; Joachimiak, G.; OrdoñEz, C.L.; Lory, S.; et al. A Virulence Locus of Pseudomonas aeruginosa Encodes a Protein Secretion Apparatus. Science 2006, 312, 1526–1530. [Google Scholar] [CrossRef] [PubMed]
- Pukatzki, S.; Ma, A.T.; Sturtevant, D.; Krastins, B.; Sarracino, D.; Nelson, W.C.; Heidelberg, J.F.; Mekalanos, J.J. Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc. Natl. Acad. Sci. USA 2006, 103, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Pilhofer, M.; Henderson, G.P.; Jensen, G.J.; Mekalanos, J.J. Type VI secretion requires a dynamic contractile phage tail-like structure. Nature 2012, 483, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Basler, M.; Ramagopal, U.A.; Bonanno, J.B.; Sauder, J.M.; Pukatzki, S.; Burley, S.K.; Almo, S.C.; Mekalanos, J.J. Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc. Natl. Acad. Sci. USA 2009, 106, 4154–4159. [Google Scholar] [CrossRef]
- Lu, W.; Lu, H.; Huo, X.; Wang, C.; Zhang, Z.; Zong, B.; Wang, G.; Dong, W.; Li, X.; Li, Y.; et al. EvfG is a multi-function protein located in the Type VI secretion system for ExPEC. Microbiol. Res. 2024, 283, 127647. [Google Scholar] [CrossRef]
- Ho, B.T.; Fu, Y.; Dong, T.G.; Mekalanos, J.J. Vibrio cholerae type 6 secretion system effector trafficking in target bacterial cells. Proc. Natl. Acad. Sci. USA 2017, 114, 9427–9432. [Google Scholar] [CrossRef]
- MacIntyre, D.L.; Miyata, S.T.; Kitaoka, M.; Pukatzki, S. The Vibrio cholerae type VI secretion system displays antimicrobial properties. Proc. Natl. Acad. Sci. USA 2010, 107, 19520–19524. [Google Scholar] [CrossRef]
- Filloux, A. The type VI secretion system: A tubular story. EMBO J. 2009, 28, 309–310. [Google Scholar] [CrossRef]
- Whitney, J.C.; Beck, C.M.; Goo, Y.A.; Russell, A.B.; Harding, B.N.; De Leon, J.A.; Cunningham, D.A.; Tran, B.Q.; Low, D.A.; Goodlett, D.R.; et al. Genetically distinct pathways guide effector export through the type VI secretion system. Mol. Microbiol. 2014, 92, 529–542. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, X.; Shou, J.; Zong, B.; Zhang, Y.; Tan, J.; Chen, J.; Hu, L.; Zhu, Y.; Chen, H.; et al. Roles of Hcp family proteins in the pathogenesis of the porcine extraintestinal pathogenic Escherichia coli type VI secretion system. Sci. Rep. 2016, 6, 26816. [Google Scholar] [CrossRef]
- Zong, B.; Zhang, Y.; Wang, X.; Liu, M.; Zhang, T.; Zhu, Y.; Zheng, Y.; Hu, L.; Li, P.; Chen, H.; et al. Characterization of multiple type-VI secretion system (T6SS) VgrG proteins in the pathogenicity and antibacterial activity of porcine extra-intestinal pathogenic Escherichia coli. Virulence 2019, 10, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Tan, J.; Lu, H.; Wang, G.; Dong, W.; Wang, C.; Li, X.; Tan, C. Function of Rhs proteins in porcine extraintestinal pathogenic Escherichia coli PCN033. J. Microbiol. 2021, 59, 854–860. [Google Scholar] [CrossRef]
- Islam, M.I.; Bae, J.H.; Ishida, T.; Ridone, P.; Lin, J.; Kelso, M.J.; Sowa, Y.; Buckley, B.J.; Baker, M.a.B. Novel Amiloride Derivatives That Inhibit Bacterial Motility across Multiple Strains and Stator Types. J. Bacteriol. 2021, 203, e00367-21. [Google Scholar] [CrossRef]
- Wadhwa, N.; Berg, H.C. Bacterial motility: Machinery and mechanisms. Nat. Rev. Microbiol. 2022, 20, 161–173. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikström, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef]
- Chaban, B.; Hughes, H.V.; Beeby, M. The flagellum in bacterial pathogens: For motility and a whole lot more. Semin. Cell Dev. Biol. 2015, 46, 91–103. [Google Scholar] [CrossRef]
- Wu, X.; Xu, J.; Yang, X.; Wang, D.; Xu, X. Integrating Transcriptomics and Metabolomics to Explore the Novel Pathway of Fusobacterium nucleatum Invading Colon Cancer Cells. Pathogens 2023, 12, 201. [Google Scholar] [CrossRef]
- Zong, B.; Xiao, Y.; Li, R.; Li, H.; Wang, P.; Yang, X.; Zhang, Y. Transcriptome and metabolome profiling to elucidate the mechanism underlying the poor growth of Streptococcus suis serotype 2 after orphan response regulator CovR deletion. Front. Vet. Sci. 2023, 10, 1280161. [Google Scholar] [CrossRef]
- Darnton, N.C.; Turner, L.; Rojevsky, S.; Berg, H.C. Dynamics of bacterial swarming. Biophys. J. 2010, 98, 2082–2090. [Google Scholar] [CrossRef]
- Duan, Q.; Zhou, M.; Liang, H.; Zhu, X.; Guo, Z.; Li, Y.; Hardwidge, P.R.; Zhu, G. Contribution of flagellin subunit FliC to piglet epithelial cells invasion by F18ab E. coli. Vet. Microbiol. 2013, 166, 220–224. [Google Scholar] [CrossRef]
- Liu, F.; Fu, J.; Liu, C.; Chen, J.; Sun, M.; Chen, H.; Tan, C.; Wang, X. Characterization and distinction of two flagellar systems in extraintestinal pathogenic Escherichia coli PCN033. Microbiol. Res. 2016, 196, 69–79. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, T.; Fossheim, L.E.; Zhang, X. FliC, a Flagellin Protein, Is Essential for the Growth and Virulence of Fish Pathogen Edwardsiella tarda. PLoS ONE 2012, 7, e45070. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, Y.; Choi, O.; Jeong, Y.; Jeong, J.; Lim, J.Y.; Kim, M.; Moon, J.S.; Suga, H.; Hwang, I. Regulation of polar flagellum genes is mediated by quorum sensing and FlhDC in Burkholderia glumae. Mol. Microbiol. 2007, 64, 165–179. [Google Scholar] [CrossRef]
- Homma, M.; Kojima, S. Roles of the second messenger c-di-GMP in bacteria: Focusing on the topics of flagellar regulation and Vibrio spp. Genes Cells 2022, 27, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Tomás, J.M.; Fulton, K.M.; Twine, S.M.; Merino, S. Generation of null mutants to elucidate the role of bacterial glycosyltransferases in bacterial motility. J. Vis. Exp. 2022, 181, e63231. [Google Scholar] [CrossRef]
- Judge, A.; Dodd, M.S. Metabolism. Essays Biochem. 2020, 64, 607–647. [Google Scholar] [CrossRef]
- Berg, H.C. The rotary motor of bacterial flagella. Annu. Rev. Biochem. 2003, 72, 19–54. [Google Scholar] [CrossRef]
- Colin, R.; Sourjik, V. Emergent properties of bacterial chemotaxis pathway. Curr. Opin. Microbiol. 2017, 39, 24–33. [Google Scholar] [CrossRef]
- Milo, R.; Jorgensen, P.; Moran, U.; Weber, G.; Springer, M. BioNumbers--the database of key numbers in molecular and cell biology. Nucleic Acids Res. 2010, 38, D750–D753. [Google Scholar] [CrossRef]
- Ling, N.; Wang, X.; Liu, D.; Shen, Y.; Zhang, D.; Ou, D.; Fan, H.; Wang, J.; Ding, Y.; Zhang, J.; et al. Role of fliC on biofilm formation, adhesion, and cell motility in Cronobacter malonaticus and regulation of luxS. Food Chem. Toxicol. 2021, 149, 111940. [Google Scholar] [CrossRef]
- Tang, H.; Billings, S.; Wang, X.; Sharp, L.; Blair, D.F. Regulated underexpression and overexpression of the FliN protein of Escherichia coli and evidence for an interaction between FliN and FliM in the flagellar motor. J. Bacteriol. 1995, 177, 3496–3503. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (5′ to 3′) | Reverese (5′ to 3′) |
|---|---|---|
| flgB | ACCTCAACGCAACACATTCC | CGCTAAGGCTCATCTGGTATTG |
| flgC | ATGGCACTGCTGAATATTTTTGATA | CACCTTTACGCCGCCTGT |
| flgD | CCACCAGTAGTAGTTCGCTCACG | GCTGACCGTGCTGATTTGTGCC |
| flgG | TACACCGAAACGCAATCCTC | TGGACACCGCTTTACTGTTG |
| flgH | GTCTACTCAGGTGGCGGATG | ACATTGGCGACAGGTTAAGGA |
| flgE | CTCAAGCGGTTAGCGGATTA | GTGGTCGTGCCATCGGTA |
| flhA | CGCCTCATCTTCACCAATCA | TTGTTCTCGATGGTATGCCG |
| fliF | TCGCACCATTCGTCATACCA | CATCTGTTCGTTGCTGAGAGG |
| fliI | CCCAATCAATGCCCTGCTTAC | CACCAATCAAACCCACGACAA |
| fliJ | GCGATATGAGTGCCGGGATG | GCTGTTCAGGGCAATGTCAA |
| fliM | GCCGACCAACCTGAACCTTA | CCACTTTAGTCGGGAAGCGT |
| fliN | CCGCACACGAATGACCATC | TATCAGCGACTACGACAACTTC |
| fliL | TGGCATTCGCATCAGGTTG | CATCTTTCAGTCGCAGGGTTAT |
| fliP | GCGCCACCTAACCAGGTATT | AACCCTAAATCCGCCTCACG |
| fliQ | CGCCACACAGATTAACGAAATG | CAGCAACAGATTGAGCATCCA |
| flgF | CACAATCTCGGTGCTCAATCC | CGACATCACACGCAAGGTT |
| flgI | CTGGTTCGGTGGTGATGAATC | GTATCTGGCTGGCTGACATTG |
| flgA | CTGCGGTAACGACAAACGATA | CAGGCTAACGGCATCAACAA |
| fliG | TGCTCGACGGTCAGAATCTC | GCGGTAATAACGGCTTCTTCC |
| fliH | GCTTGATAGTGTGATTGCTTCG | CGCTGAATAACGGTTCTTGCT |
| fliE | CCGCACCCGACCATTAGTT | AGCTTATTACGCACCTGAATCC |
| tsr | CAGCGATAACAATGCCTCCTAC | TCATTAGAGCCATCCACCTCAA |
| motA | ATAGAGCGTTTTGCGACCGA | CGATGGTGGGGACTTTCCTC |
| motB | GCTGATTCAGATTGCGGAGTA | ATGTTCGGCTGCTTATTCACTT |
| 16sRNA | GAATGCCACGGTGAATAC | GGTTACCTTGTTACGACTTC |
| Pathway | Metabolite | Formula | Peak Area | VIP | FC | p_Value | Type | |
|---|---|---|---|---|---|---|---|---|
| ΔevfG | PCN033 | |||||||
| Citrate cycle (TCA cycle) | Citric Acid | C6H8O7 | 5.05 ± 0.168 | 5.479 ± 0.076 | 1.2968 | 0.922 | 0.02568 | down |
| Malic Acid | C4H6O5 | 5.678 ± 0.036 | 5.411 ± 0.081 | 1.4736 | 1.049 | 0.0006089 | up | |
| Glyoxylate and dicarboxylate metabolism | Citric Acid | C6H8O7 | 5.05 ± 0.168 | 5.479 ± 0.076 | 1.2968 | 0.922 | 0.02568 | down |
| 3-Phosphoglycerate | C3H7O7P | 3.034 ± 0.201 | 4.617 ± 0.849 | 2.3762 | 0.657 | 0.07599 | down | |
| Malic Acid | C4H6O5 | 5.678 ± 0.036 | 5.411 ± 0.081 | 1.4736 | 1.049 | 0.0006089 | up | |
| Pentose phosphate pathway | Deoxyribose | C5H10O4 | 5.58 ± 0.04 | 5.957 ± 0.06 | 1.1794 | 0.937 | 0.03162 | down |
| 3-Phosphoglycerate | C3H7O7P | 3.034 ± 0.201 | 4.617 ± 0.849 | 2.3762 | 0.657 | 0.07599 | down | |
| Glycine, serine and threonine metabolism | Choline | C5H14NO+ | 6.157 ± 0.123 | 6.492 ± 0.04 | 1.1949 | 0.948 | 0.01386 | down |
| L-Tryptophan | C11H12N2O2 | 4.294 ± 0.061 | 4.935 ± 0.098 | 1.7677 | 0.87 | 0.002812 | down | |
| 3-Phosphoglycerate | C3H7O7P | 3.034 ± 0.201 | 4.617 ± 0.849 | 2.3762 | 0.657 | 0.07599 | down | |
| Aminoacyl-tRNA biosynthesis | L-Tryptophan | C11H12N2O2 | 4.294 ± 0.061 | 4.935 ± 0.098 | 1.7677 | 0.87 | 0.002812 | down |
| L-Phenylalanine | C9H11NO2 | 4.114 ± 0.125 | 4.496 ± 0.078 | 1.2485 | 0.915 | 0.0234 | down | |
| Tryptophan metabolism | Serotonin | C10H12N2O | 4.104 ± 0.042 | 4.729 ± 0.131 | 1.702 | 0.868 | 0.005014 | down |
| L-Tryptophan | C11H12N2O2 | 4.294 ± 0.061 | 4.935 ± 0.098 | 1.7677 | 0.87 | 0.002812 | down | |
| Alanine, aspartate and glutamate metabolism | Gamma-Aminobutyric Acid | C4H9NO2 | 5.967 ± 0.067 | 6.419 ± 0.073 | 1.4728 | 0.93 | 0.002632 | down |
| Citric Acid | C6H8O7 | 5.05 ± 0.168 | 5.479 ± 0.076 | 1.2968 | 0.922 | 0.02568 | down | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zong, B.; Wang, P.; Liu, W.; Wu, A.; Xiao, Y.; Fu, S.; Qiu, Y.; Zhang, Y.; Liu, W. The Regulatory Role of EvfG Through Coordinated Control of Flagellar Biosynthesis and Energy Metabolism in Porcine Extraintestinal Pathogenic Escherichia coli (ExPEC). Biology 2025, 14, 822. https://doi.org/10.3390/biology14070822
Zong B, Wang P, Liu W, Wu A, Xiao Y, Fu S, Qiu Y, Zhang Y, Liu W. The Regulatory Role of EvfG Through Coordinated Control of Flagellar Biosynthesis and Energy Metabolism in Porcine Extraintestinal Pathogenic Escherichia coli (ExPEC). Biology. 2025; 14(7):822. https://doi.org/10.3390/biology14070822
Chicago/Turabian StyleZong, Bingbing, Peiyi Wang, Wei Liu, Aihua Wu, Yong Xiao, Shulin Fu, Yinsheng Qiu, Yanyan Zhang, and Wentong Liu. 2025. "The Regulatory Role of EvfG Through Coordinated Control of Flagellar Biosynthesis and Energy Metabolism in Porcine Extraintestinal Pathogenic Escherichia coli (ExPEC)" Biology 14, no. 7: 822. https://doi.org/10.3390/biology14070822
APA StyleZong, B., Wang, P., Liu, W., Wu, A., Xiao, Y., Fu, S., Qiu, Y., Zhang, Y., & Liu, W. (2025). The Regulatory Role of EvfG Through Coordinated Control of Flagellar Biosynthesis and Energy Metabolism in Porcine Extraintestinal Pathogenic Escherichia coli (ExPEC). Biology, 14(7), 822. https://doi.org/10.3390/biology14070822